
Relationships between Some Biodiversity Indicators and Crown Damage of Pinus sylvestris L. in Natural Old Growth Pine Forests

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Research Area
2.1.1. Kampinos National Park
- Czerwińskie Góry (CG)—species dominating the forest areas: pine about 200–210 years old *, habitat type: fresh mixed coniferous forest.
- Wilków (W)—species dominating the forest areas: pine about 180–200 years old *, habitat type: fresh coniferous forest.
- Granica (Gr)—species dominating the forest areas: pine about 160–170 years old *, habitat type: fresh mixed coniferous forest.
- Sieraków (S)—species dominating the forest areas: pine about 190–200 years old *, habitat type: fresh coniferous forest.
2.1.2. Bory Tucholskie National Park
- Gacno (Ga)—species dominating the forest areas: pine about 140–150 years old *, habitat type: fresh coniferous forest.
- Płęsno (P)—species dominating the forest areas: pine about 120–130 years old *, habitat type: fresh coniferous forest.
- Mielnica (M)—species dominating the forest areas: pine about 150–190 years old *, habitat type: fresh mixed coniferous and fresh coniferous forest.
- Kocioł (K)—species dominating the forest areas: pine about 100–110 years old *, habitat type: fresh coniferous forest.
2.2. Collection of Plant Material and Observations of Defoliation Level
2.3. Molecular Analyses
2.4. Phytosociological Evaluation
2.5. Statistical Analysis
2.5.1. SSR Analysis
2.5.2. Phytosociological Analysis
2.5.3. Stand Health Analysis
3. Results
3.1. Health Condition of Tree Crowns
3.2. Biodiversity at the Genetic Level
3.3. Biodiversity at the Species Level
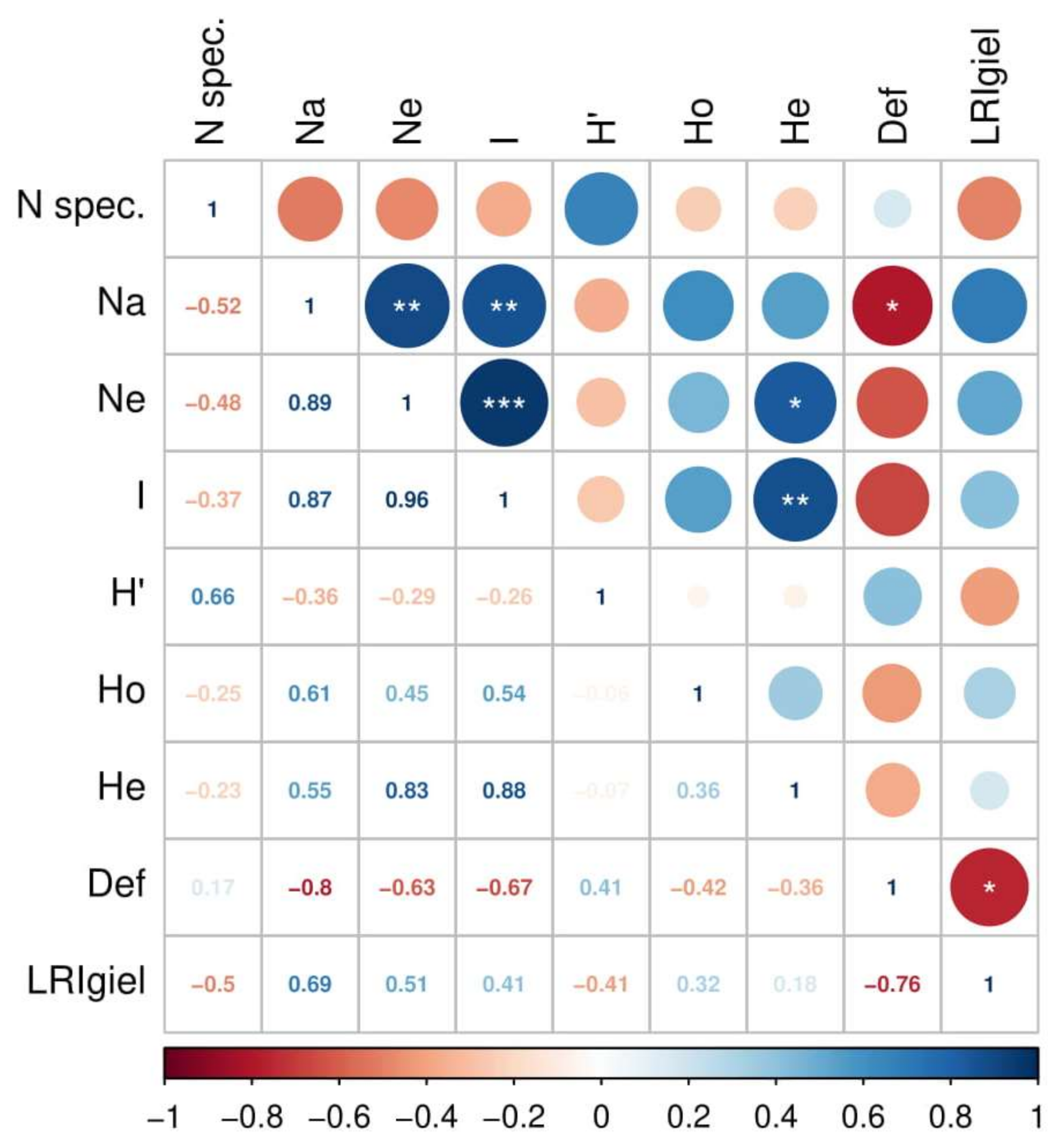
3.4. Correlation between Biodiversity and Stand Health
4. Discussion
4.1. Defoliation
4.2. Species Diversity
4.3. Genetic Diversity
4.3.1. Genetic Diversity of Populations
4.3.2. Interactions Observed
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stock, T. Convention on Biological Diversity; UNEP: Rio de Janeiro, Brazil, 1992; (1760 U.N.T.S. 69). [Google Scholar]
- Burczyk, J. Mating system variation in a Scots pine clonal seed orchard. Silvae Genet. 1998, 47, 155–158. [Google Scholar]
- Adams, W.T.; Campbell, R.K.; Kitzmiller, J.H. Genetic considerations in reforestation. In Reforestation Practices in Southwestern Oregon and Northern California; Hobbs, S.D., Tesch, S.D., Owston, P.W., Eds.; Forest Research Laboratory, Oregon State University: Corvallis, OR, USA, 1992; pp. 284–308. [Google Scholar]
- Klisz, M.; Jastrzębowski, S.; Ukalski, K.; Ukalska, J.; Przybylski, P. Adaptation of Norway spruce populations in Europe: A case study from northern Poland. New Zealand For. Sci. 2017, 47, 2–9. [Google Scholar] [CrossRef]
- Ivetić, V.; Devetaković, J.; Nonić, M.; Stanković, D.; Šijačić-Nikolić, M. Genetic diversity and forest reproductive material–from seed source selection to planting. iForest Biogeosciences For. 2016. [Google Scholar] [CrossRef]
- Bell, D.L.; Sultan, S.E. Dynamic phenotypic plasticity for root growth in Polygonum: A comparative study. Am. J. Bot. 1999, 86, 807–819. [Google Scholar] [CrossRef] [PubMed]
- Mitchell-Olds, T. Genetic constraints on life-history evolution: Quantitative-trait loci influencing growth and flowering in Arabidopsis thaliana. Evolution 1996, 50, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Isajev, V.; Ivetić, V.; Lučić, A.; Rakonjac, L.J. Gene pool conservation and tree improvement in Serbia. Genetika 2009, 41, 309–327. [Google Scholar] [CrossRef]
- Bergelson, J.; Roux, F. Towards identifying genes underlying ecologically relevant traits in Arabidopsis thaliana. Nat. Rev. Genet. 2010, 11, 867–879. [Google Scholar] [CrossRef] [PubMed]
- Glazier, A.M.; Nadeau, J.H.; Aitman, T.J. Finding genes that underlie complex traits. Science 2002, 298, 2345–2349. [Google Scholar] [CrossRef]
- Zgorzelski, M.; Pawłowska, T. Geomorfologia. Kampinoski Park Narodowy. In Przyroda Kampinoskiego Parku Narodowego; Andrzejewski, R., Ed.; Kampinoski Park Narodowy: Izabelin, Poland, 2003; Volume 1, pp. 87–95. [Google Scholar]
- Matuszkiewicz, J.M. Biogeograficzna charakterystyka obszaru Parku Narodowego “Bory Tucholskie”. In Świat Roślin i Grzybów Parku Narodowego “Bory Tucholskie”; Matuszkiewicz, J.M., Ed.; National Park „Bory Tucholskie”: Charzykowy, Poland, 2012; pp. 13–31. [Google Scholar]
- Spellerberg, I.F.; Fedor, P.J. A tribute to Claude-Shannon (1916–2001) and a plea for more rigorous use of species richness, species diversity and the “Shannon-Wiener” Index. Glob. Ecol. Biogeogr. 2003, 12, 177–179. [Google Scholar] [CrossRef]
- Vendramin, G.G.; Anzidei, M.; Madaghiele, A.; Sperisen, C.; Bucci, G. Chloroplast microsatellite analysis reveals the presence of population subdivision in Norway spruce (Picea abies K.). Genome 2000, 43, 68–78. [Google Scholar] [CrossRef]
- Rajora, O.P.; Rahman, M.H.; Buchert, G.P.; Dancik, B.P. Microsatellite DNA analysis of genetic effects of harvesting on old-growth eastern white pine (Pinus strobus L.) in Ontario, Canada. Mol. Ecol. 2000, 9, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Klisz, M.; Ukalska, J.; Koprowski, M.; Tereba, A.; Puchałka, R.; Przybylski, P.; Jastrzębowski, S.; Nabais, C. Effect of provenance and climate in intra-annual density fluctuations of Norway spruce Picea abies (L.) Karst. Pol. Agric. Meteorol. 2019, 269–270, 145–156. [Google Scholar] [CrossRef]
- Williams, C. The peculiarities of pine genome. In Proceedings of the Plant and Animal Genome VII Conference, San Diego, CA, USA, 17−21 January 1999. [Google Scholar]
- Scalfi, M.; Piotti, A.; Rossi, M.; Piovani, P. Genetic variability of Italian southern Scots pine (Pinus sylvestris L.) populations: The rear edge of the range. Eur. J. For. Res. 2009, 128, 377. [Google Scholar] [CrossRef]
- Konecka, A.; Tereba, A.; Studnicki, M.; Nowakowska, J.A. Allele rzadkie i prywatne jako miara bogactwa puli genetycznej materiału sadzeniowego sosny zwyczajnej. Sylwan 2019, 163, 948–956. [Google Scholar] [CrossRef]
- Lorenz, M. International co-operative programme on assessment and monitoring of air pollution effects on forests–ICP forests. Water Air Soil Pollut. 1995, 85, 1221–1226. [Google Scholar] [CrossRef]
- Lech, P. Przydatność szacunkowej metody określania defoliacji drzew do badań stanu zdrowotnego lasu. Sylwan 1995, 8, 99–107. [Google Scholar]
- Dobbertin, M.; Brang, P. Crown defoliation improves tree mortality models. For. Ecol. Manag. 2001, 141, 271–284. [Google Scholar] [CrossRef]
- Jaszczak, R.; Miotke, M. Defoliation of the top part and whole crown of Scots pine (Pinus sylvestris L.). Sylwan 2009, 153, 607–616. [Google Scholar]
- Soranzo, N.; Provan, J.; Powell, W. Characterization of microsatellite loci in Pinus sylvestris L. Mol. Ecol. 1998, 7, 1260–1261. [Google Scholar]
- Chagne, D.; Chaumeil, P.; Ramboer, A.; Collada, C.; Guevara, A.; Cervera, M.T.; Vendramin, G.G.; Garcia, V.; Frigerio, J.M.; Echt, C.; et al. Cross-species transferability and mapping of genomic and CDNA SSRs in pines. Theor. Appl. Gen. 2004, 109, 1204–1214. [Google Scholar] [CrossRef]
- Matuszkiewicz, W. Przewodnik Do Oznaczania Zbiorowisk Roślinnych Polski; PWN: Warszawa, Poland, 2006. [Google Scholar]
- GBIF.org. GBIF Home Page. 2020. Available online: https://www.gbif.org (accessed on 13 January 2021).
- Peakall, R.; Smouse, P. GENEALEX 6.5: Genetic Analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Goudet, J. FSTAT, a Program to Estimate and Test Gene Diversities and Fixation Indices (Version 2.9.4). Available online: http://www2.unil.ch/popgen/softwares/fstat.htm (accessed on 21 November 2003).
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Dzwonko, Z.; Loster, S. Wskaźnikowe gatunki roślin starych lasów i ich znaczenie dla ochrony przyrody i kartografii roślinności. Prace Geogr. IGiPZ Pan. 2001, 178, 120–132. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 29 March 2020).
- Silva, L.J.; Laercio, S. Duncan Test, Tukey Test and Scott-Knott Test. R package Version 1.0-1. Available online: https://CRAN.R-project.org/package=laercio (accessed on 29 March 2020).
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix (Version 0.84). Available online: https://github.com/taiyun/corrplot (accessed on 29 March 2020).
- Vu, V.Q. Ggbiplot: A Ggplot2 Based Biplot. R Package Version 0.55. Available online: http://github.com/vqv/ggbiplot (accessed on 29 March 2020).
- Sierota, Z. Przerzedzenie koron drzew jako efekt stresu i źródło stresu. Sylwan 1995, 8, 5–24. [Google Scholar]
- Wójcik, R. Analiza zmian ilościowych aparatu asymilacyjnego w drzewostanach sosnowych nadleśnictwa Wyszków. Sylwan 2000, 144, 39–46. [Google Scholar]
- Borecki, T.; Wójcik, R. Ocena stanu uszkodzenia drzewostanów Nadleśnictwa Krotoszyn. Sylwan 1996, 7, 5–21. [Google Scholar]
- Kalliokoski, T. Root System Traits of Norway Spruce, Scots Pine and Silver Birch in Mixed Boreal Forests: An Analysis of Root Architecture, Morphology and Anatomy. Ph.D. Thesis, University of Helsinki, Helsinki, Finland, 2011. [Google Scholar]
- Mickovski, S.; Ennos, A.R. A morphological and mechanical study of the root systems of suppressed crown Scots pine. Trees 2002, 16, 274–280. [Google Scholar] [CrossRef]
- Kozlowski, T.T. (Ed.) Water Deficits and Plant Growth; Academic Press: Cambridge, MA, USA, 1978; Volume 5. [Google Scholar]
- Tyburski, Ł.; Przybylski, P. Health condition of the Scots pine (Pinus sylvestris L.) in Kampinos National Park–preliminary studies. Folia For. Pol. 2016, 58, 240–245. [Google Scholar] [CrossRef]
- Głowacki, Z.; Ferchmin, M. Chronione, rzadkie i zagrożone gatunki roślin naczyniowych Kampinoskiego Parku Narodowego. In Kampinoski Park Narodowy. Przyroda Kampinoskiego Parku Narodowego; Andrzejewski, R., Ed.; Kampinoski Park Narodowy: Izabelin, Poland, 2003; Volume 1, pp. 259–274. [Google Scholar]
- Robledo-Arnuncio, J.J.; Smouse, P.E.; Gil, L.; Alía, R. Pollen movement under alternative silvicultural practices in native populations of Scots pine (Pinus sylvestris L.) in central Spain. For. Ecol. Manag. 2004, 197, 245–255. [Google Scholar] [CrossRef]
- Raja, R.G.; Tauer, C.G.; Wittwer, R.F.; Huang, Y. Regeneration methods affect genetic variation and structure in shortleaf pine (Pinus echinata Mill.). For. Genet. 1998, 5, 171–178. [Google Scholar]
- Nowakowska, J.A. Zmienność Genetyczna Polskich Wybranych Populacji Sosny Zwyczajnej (Pinus sylvestris L.) na Podstawie Analiz polimorfizmu DNA; Instytut Badawczy Leśnictwa: Sękocin Nowy, Poland, 2007. [Google Scholar]
- Ramakrrishnan, U.; Hadly, E.A.; Mountain, J.L. Detecting past population bottlenecks using temporal genetic data. Mol. Ecol. 2005, 14, 2915–2922. [Google Scholar] [CrossRef] [PubMed]
- Chybicki, I.J.; Dzialuk, A.; Trojankiewicz, M.; Slawski, M.; Burczyk, J. Spatial genetic structure within two contrasting stands of scots pine (Pinus sylvestris L.). Silvae Genet. 2007, 57, 4–5. [Google Scholar] [CrossRef][Green Version]
- Kavaliauskas, D. Genetic Structure and Genetic Diversity of Scots Pine (Pinus sylvestris L.) in Latuania. Ph.D. Thesis, Aleksandras Stulginskis University, Akademija, Lituania, 2015. [Google Scholar]
- Winicov, I. New molecular approaches to improving salt tolerance in crop plants. Ann. Bot. 1998, 82, 703–710. [Google Scholar] [CrossRef]
- Parsons, P.A. Conservation genetics: Adaptation to stress and the preservation of genetic diversity. Biol. J. Linn. Soc. 1996, 58, 471–482. [Google Scholar] [CrossRef]
- Boer Den, P.J.; Szyszko, J.; Vermeulen, R. Spreading the risk of extinction by genetic diversity in populations of the carabid beetle Pterostichus oblongopunctatus F. (Coleoptera, Carbidae). Neth. J. Zool. 1993, 43, 242–259. [Google Scholar] [CrossRef]
- Rankevich, D.; Lavie, B.; Nevo, E.; Beiles, A.; Arad, Z. Genetic and physiological adaptations of the prosobranch landsnail Pomatia olivieri to microclimatic stresses on Mount Carmel. Israel. Isr. J. Zool. 1996, 42, 425–441. [Google Scholar]
- Kopp, R.L.; Wissing, T.E.; Guttman, S.I. Genetic indicators of environmental tolerance among fish populations exposed to acid deposition. Biochem. Syst. Ecol. 1994, 22, 459–475. [Google Scholar] [CrossRef]
- Via, S.; Lande, R. Evolution of genetic variability in a spatially heterogeneous environment: Effects of genotype-environment interaction. Genet. Res. 1987, 49, 147–156. [Google Scholar] [CrossRef]
- Scheiner, S.M. Genetics and evolution of phenotypic plasticity. Ann. Rev. Ecol. Syst. 1993, 23, 1–14. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Loci | Repeat Motif. | Starter Sequences | Product Size |
|---|---|---|---|
| SPAG 7.14(VIC) | (TG)17(AG)21 | F: TTCGTAGGACTAAAAATGTGTG R: CAAAGTGGATTTTGACCG | 209 |
| SPAC 11.6(NED) | (CA)29(TA)7 | F: CTTCACAGGACTGATGTTCA R: TTACAGCGGTTGGTAAATG | 165 |
| NZPR 11.4(6-FAM) | (CA)15(CA)13(TA)22 | F: AAGATGACCCACATGAAGTTTGG R: GGAGCTTTATAACATATCTCGATGC | 193 |
| SsrPt_ctg4363(VIC) | (AT)10 | F: TAATAATTCAAGCCACCCCG R: AGCAGGCTAATAACAACACGC | 100 |
| PtTX3107(PET) | (CAT)14 | F: AAACAAGCCCACATCGTCAATC R: TCCCCTGGATCTGAGGA | 160 |
| Braun–Blanquet Score | Value Taken for Calculations |
|---|---|
| r | 0.1 |
| + | 0.5 |
| 1 | 2.5 |
| 2 | 12.5 |
| 3 | 37.5 |
| 4 | 62.5 |
| 5 | 87.5 |
| Df | Sum. Sq | Mean Sq | F Value | Pr (>F) | ||
|---|---|---|---|---|---|---|
| Defoliation | Population | 7.00 | 10,466.00 | 1495.20 | 11.26 | 5.22 × 10−13 *** |
| Residuals | 390.00 | 51,794.00 | 132.80 | |||
| LRIgiel | Population | 7.00 | 8.78 | 1.25 | 2.96 | 0.00481 ** |
| Residuals | 390.00 | 164.77 | 0.42 |
| Populations | Defoliation (%) | LRIgiel |
|---|---|---|
| Granica | a(46.9) | a(1.52) |
| Cz. Góry | ab(41.1) | ab(1.78) |
| Wilków | abc(39.9) | b(1.92) |
| Sieraków | bc(38.7) | ab(1.86) |
| Gacno | bcd(35.5) | b(2.00) |
| Mielnca | cd(33.3) | b(1.92) |
| Kocioł | d(31.2) | b(2.02) |
| Płęsno | d(31.1) | ab(1.81) |
| Population | N Spec. | Na | Ne | I | Ho | Fis |
|---|---|---|---|---|---|---|
| Sieraków KNP | 25 | 14.400 | 8.438 | 2.202 | 0.579 | 0.324 * |
| S.E. | 2.731 | 2.156 | 0.256 | 0.080 | ||
| Cz. Góry KNP | 37 | 13.400 | 7.778 | 2.151 | 0.764 | 0.101 |
| S.E. | 2.619 | 1.868 | 0.235 | 0.064 | ||
| Granica KNP | 36 | 13.200 | 8.093 | 2.160 | 0.644 | 0.244 * |
| S.E. | 2.267 | 2.001 | 0.247 | 0.059 | ||
| Wilków KNP | 21 | 13.400 | 7.670 | 2.081 | 0.627 | 0.246 * |
| S.E. | 2.600 | 2.145 | 0.276 | 0.064 | ||
| BTNP Kocioł | 15 | 18.000 | 9.864 | 2.384 | 0.779 | 0.110 * |
| S.E. | 4.528 | 2.706 | 0.274 | 0.044 | ||
| BTNP Gacno | 16 | 17.200 | 8.713 | 2.222 | 0.764 | 0.090 * |
| S.E. | 4.620 | 2.665 | 0.325 | 0.074 | ||
| BTNP Płęsno | 33 | 15.800 | 8.375 | 2.242 | 0.723 | 0.145 * |
| S.E. | 3.292 | 2.415 | 0.278 | 0.026 | ||
| BTNP Mielnica | 49 | 15.000 | 8.364 | 2.187 | 0.656 | 0.224 * |
| S.E. | 3.564 | 2.346 | 0.289 | 0.086 | ||
| Total | 29 | 15.050 | 8.412 | 2.204 | 0.692 | 0.185 * |
| S.E. | 1.117 | 0.745 | 0.089 | 0.024 |
| Sieraków KNP | Cz. Góry KNP | Granica KNP | Wilków KNP | BTNP Kocioł | BTNP Gacno | BTNP Płęsno | BTNP Mielnica | |
|---|---|---|---|---|---|---|---|---|
| Sieraków KNP | 0.000 | ns | ns | ns | ns | ns | 0.014 | 0.028 |
| Cz.Góry KNP | 0.010 | 0.000 | ns | ns | ns | 0.044 | ns | 0.024 |
| Granica KNP | 0.010 | 0.007 | 0.000 | ns | ns | ns | 0.014 | ns |
| Wilków KNP | 0.008 | 0.009 | 0.011 | 0.000 | ns | ns | 0.000 | 0.001 |
| BTNP Kocioł | 0.008 | 0.008 | 0.007 | 0.008 | 0.000 | ns | ns | ns |
| BTNP Gacno | 0.006 | 0.009 | 0.008 | 0.006 | 0.007 | 0.000 | 0.004 | 0.003 |
| BTNP Płęsno | 0.010 | 0.009 | 0.012 | 0.016 | 0.012 | 0.010 | 0.000 | ns |
| BTNP Mielnica | 0.011 | 0.011 | 0.009 | 0.014 | 0.010 | 0.011 | 0.006 | 0.000 |
| Number of Species | BTNP | KNP | BTNP + KNP |
|---|---|---|---|
| Total number of species | 59 | 55 | 84 |
| Vascular plant species | 45 | 53 | 69 |
| Bryophytes species | 11 | 2 | 12 |
| Lichen species | 3 | 0 | 3 |
| Shannon–Wiener index (H’) | 2.14137 | 2.29675 | 2.40883 |
| National Park | Surface | Number of Species in the Plot | Number of old Growth Forest Species | Age of the Dominant P. sylvestris L. in the Stand | H’ | SD |
|---|---|---|---|---|---|---|
| KNP | CG | 36 | 12 | 200–210 (avg. 205) | 1.79144 | 0.31258 |
| Gr | 34 | 15 | 160–170 (avg. 165) | 1.96834 | 0.28773 | |
| W | 21 | 10 | 180–200 (avg. 190) | 1.36353 | 0.09822 | |
| S | 25 | 10 | 190–200 (avg. 195) | 1.83306 | 0.36663 | |
| BTNP | M | 48 | 15 | 150–190 (avg. 170) | 2.10052 | 0.28560 |
| Ga | 16 | 3 | 140–150 (avg. 145) | 1.68053 | 0.06668 | |
| K | 15 | 3 | 100–110 (avg. 105) | 1.51323 | 0.13578 | |
| P | 33 | 4 | 120–130 (avg. 125) | 1.96201 | 0.37896 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Przybylski, P.; Mohytych, V.; Rutkowski, P.; Tereba, A.; Tyburski, Ł.; Fyalkowska, K. Relationships between Some Biodiversity Indicators and Crown Damage of Pinus sylvestris L. in Natural Old Growth Pine Forests. Sustainability 2021, 13, 1239. https://doi.org/10.3390/su13031239
Przybylski P, Mohytych V, Rutkowski P, Tereba A, Tyburski Ł, Fyalkowska K. Relationships between Some Biodiversity Indicators and Crown Damage of Pinus sylvestris L. in Natural Old Growth Pine Forests. Sustainability. 2021; 13(3):1239. https://doi.org/10.3390/su13031239
Chicago/Turabian StylePrzybylski, Paweł, Vasyl Mohytych, Paweł Rutkowski, Anna Tereba, Łukasz Tyburski, and Kateryna Fyalkowska. 2021. "Relationships between Some Biodiversity Indicators and Crown Damage of Pinus sylvestris L. in Natural Old Growth Pine Forests" Sustainability 13, no. 3: 1239. https://doi.org/10.3390/su13031239
APA StylePrzybylski, P., Mohytych, V., Rutkowski, P., Tereba, A., Tyburski, Ł., & Fyalkowska, K. (2021). Relationships between Some Biodiversity Indicators and Crown Damage of Pinus sylvestris L. in Natural Old Growth Pine Forests. Sustainability, 13(3), 1239. https://doi.org/10.3390/su13031239