
Generating Multifunctional Landscape through Reforestation with Native Trees in the Tropical Region: A Case Study of Gunung Dahu Research Forest, Bogor, Indonesia




 , ,
, ,
Abstract
1. Introduction
2. Methodology
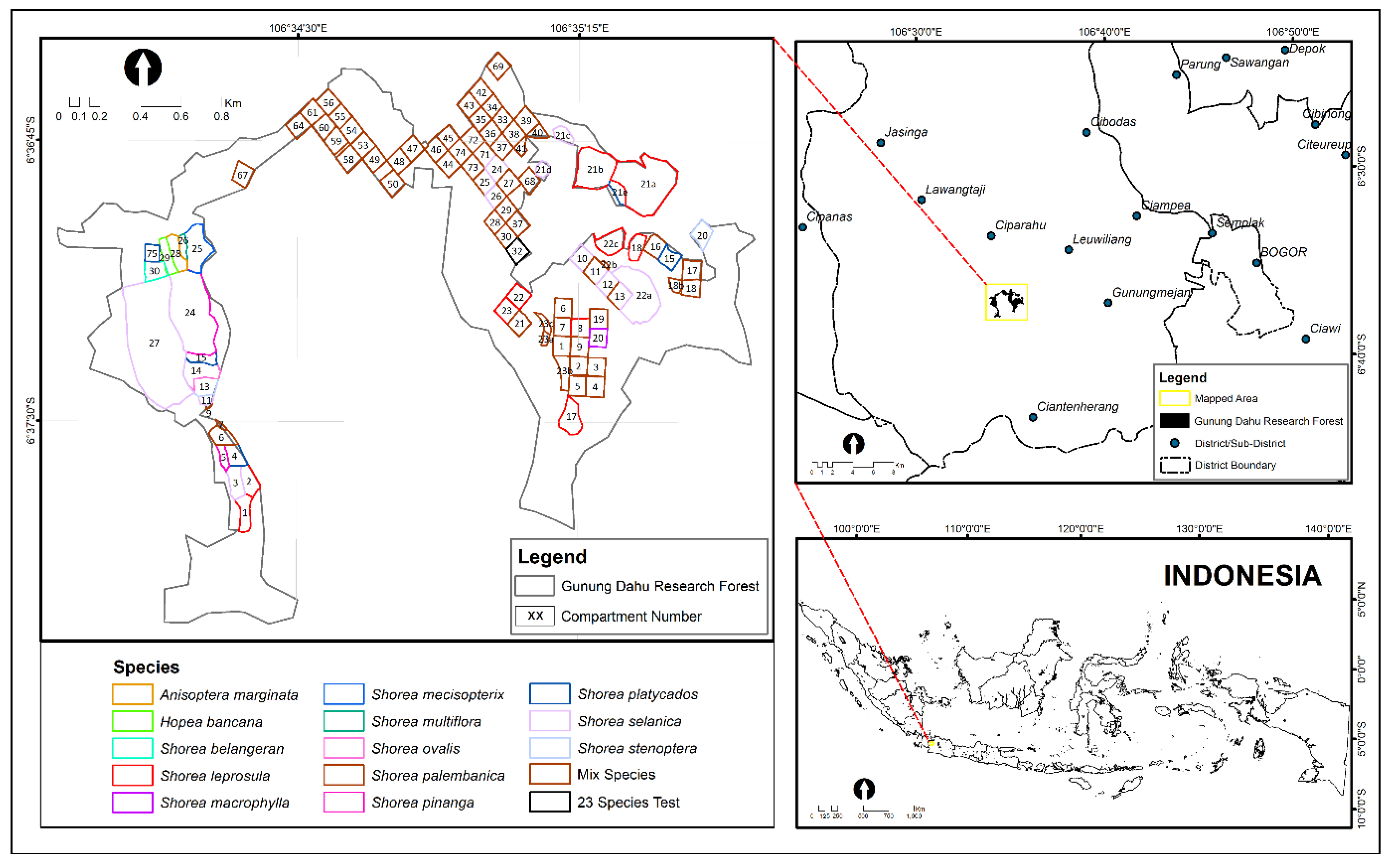
2.1. Study Area
2.2. Measurement of the Impact Generated by Reforestation
- (a)
- How reforestation by native trees yield timber stock and increase forest cover;
- (b)
- To what extent planted native trees can support a self-sustaining ecosystem through their natural regeneration capacity;
- (c)
- How reforestation can enrich soil characteristics, and increase land productivity and potential hydrological value;
- (d)
- How biological interdependent has been established from successful reforestation using native trees;
- (e)
- To what extend reforestation success can also provide economic value to the surrounding community.
2.2.1. Measurement on Growth Performance, Timber Stock, and Forest Cover Dynamic
2.2.2. Review and Measurement on Natural Regeneration Capacity
2.2.3. Measurement of Soil Physicochemical Characteristics and Potential Hydrological Value
2.2.4. Review on Ectomycorrhizal Association
2.2.5. Assessment of the Impact of Reforestation on Environmental Services and the Community Livelihood
3. Results and Discussion
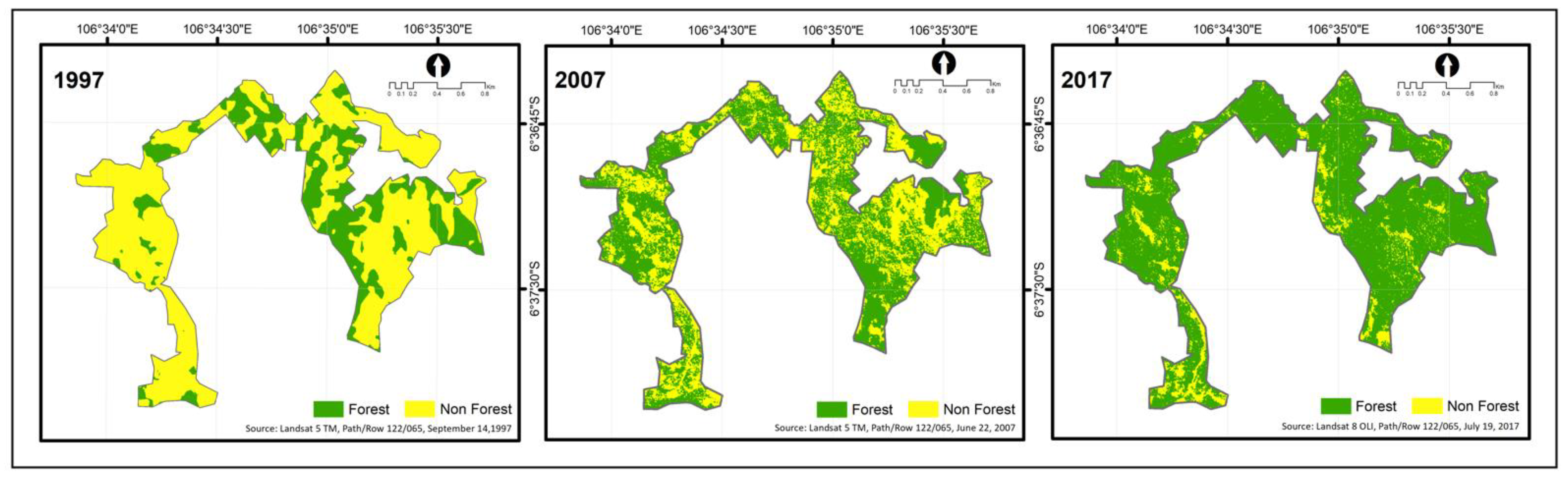
3.1. Reforestation Yields Timber Stock and Increases Forest Cover
3.2. The Capacity of Natural Regeneration of the Planted Trees Plays a Key Role in the Self-Sustaining Forest
3.3. Reforestation Improves the Productivity of Land, Soil Characteristic, and the Potential Hydrological Value
3.4. The Formation of the Ectomycorrhizal Association in Reforested GDRF Landscape
3.5. Reforested Landscape Delivers Environmental Services Value and Supports Community Livelihood
3.6. The Challenges and Opportunities of Restoration in the GDRF Landscape
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Yirdaw, E.; Tigabu, M.; Monge, A. Rehabilitation of degraded dryland ecosystems—Review. Silva Fenn. 2017, 50, 32. [Google Scholar] [CrossRef]
- Scholes, R.; Montanarella, L.; Brainich, A.; Barge, N.; ten Brink, B.; Cantele, M.; Erasmus, B.; Fisher, J.; Gardner, T.; Holland, T.G.; et al. Summary for Policymakers of the Assessment Report on Land Degradation and Restoration of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; IPBES Secretariat: Bonn, Germany, 2018. [Google Scholar]
- Kettle, C.J. Ecological considerations for using dipterocarps for restoration of lowland rainforest in southeast asia. Biodivers. Conserv. 2010, 19, 1137–1151. [Google Scholar] [CrossRef]
- Potapov, P.; Hansen, M.C.; Laestadius, L.; Turubanova, S.; Yaroshenko, A.; Thies, C.; Smith, W.; Zhuravleva, I.; Komarova, A.; Minnemeyer, S.; et al. Esipova E. The last frontiers of wilderness: Tracking loss of intact forest landscapes from 2000 to 2013. Sci. Adv. 2017, 3, e1600821. [Google Scholar] [CrossRef] [PubMed]
- Tidwell, T.L. Nexus between food, energy, water, and forest ecosystems in the USA. J. Environ. Stud. Sci. 2016, 6, 214–224. [Google Scholar] [CrossRef]
- Lamb, D.; Erskine, P.D.; Parrotta, J.A. Restoration of degraded tropical forest landscapes. Science 2005, 5754, 1628–1632. [Google Scholar] [CrossRef]
- Fisher, R.F. Amelioration of degraded rain forest soils by plantations of native trees. Soil Sci. Soc. Am. J. 2006, 59, 544–549. [Google Scholar] [CrossRef]
- Castro, J.; Zamora, R.; Hódar, J.A.; Gómez, J.M. Use of shrubs as nurse plants: A new technique for reforestation in mediterranean mountains. Restor. Ecol. 2002, 10, 297–305. [Google Scholar] [CrossRef]
- Elliott, S.; Navakitbumrung, P.; Kuarak, C.; Zangkum, S.; Anusarnsunthorn, V.; Blakesley, D. Selecting framework tree species for restoring seasonally dry tropical forests in northern thailand based on field performance. For. Ecol. Manag. 2003, 184, 177–191. [Google Scholar] [CrossRef]
- Kitao, M.; Yoneda, R.; Tobita, H.; Matsumoto, Y.; Maruyama, Y.; Arifin, A.; Azani, A.M.; Muhamad, M.N. Susceptibility to photoinhibition in seedlings of six tropical fruit tree species native to malaysia following transplantation to a degraded land. Trees-Struct. Funct. 2006, 20, 601–610. [Google Scholar] [CrossRef]
- Kenzo, T.; Yoneda, R.; Matsumoto, Y.; Azani, M.A.; Majid, N.M. Leaf photosynthetic and growth responses on four tropical tree species to different light conditions in degraded tropical secondary forest, Peninsular Malaysia. Jpn. Agric. Res. Q. 2008, 42, 299–306. [Google Scholar] [CrossRef]
- Lamb, D. Regreening the Bare Hills; Springer, Business Media: New York, NY, USA, 2011. [Google Scholar]
- Berndes, G.; Abts, B.; Asikainen, A.; Cowie, A.L.; Dale, V.; Egnell, G.; Lindner, M.; Marelli, L.; Paré, D.; Pingoud, K.; et al. Forest biomass, carbon neutrality and climate change mitigation. Sci. Policy 2016, 3, 3–27. [Google Scholar]
- Nunes, L.J.R.; Meireles, C.I.R.; Gomes, C.J.P.; Ribeiro, N.M.C.A. Forest management and climate change mitigation: A review on carbon cycle flow models for the sustainability of resources. Sustainability 2019, 11, 5276. [Google Scholar] [CrossRef]
- Kirschbaum, M.U.F. The Carbon Sequestration Potential of Tree Plantations in Australia. In Environmental Management: The Role of Eucalypts and Other Fast Growing Species; Eldridge, K.G., Crowe, M.P., Old, K.M., Eds.; CSIRO Forestry and Forest Products: Canberra, Australia, 1996; pp. 77–89. [Google Scholar]
- Nejat, P.; Jomehzadeh, F.; Taheri, M.M.; Gohari, M.; Muhd, M.Z.A. A Global review of energy consumption, CO2 emissions and policy in the residential sector (with an overview of the top ten co2 emitting countries). Renew. Sustain. Energy Rev. 2015, 43, 843–862. [Google Scholar] [CrossRef]
- Rahman, F.A.; Aziz, M.M.A.; Saidur, R.; Bakar, W.A.W.A.; Hainin, M.R.; Putrajaya, R.; Hassan, N.A. Pollution to solution: Capture and sequestration of carbon dioxide (CO2) and its utilization as a renewable energy source for a sustainable future. Renew. Sustain. Energy Rev. 2017, 71, 112–126. [Google Scholar] [CrossRef]
- Wang, J.; Meng, J. Identifying indigenous tree species for land reforestation, forest restoration, and plantation transformation on Hainan Island, China. J. Mt. Sci. 2018, 15, 2433–2444. [Google Scholar] [CrossRef]
- Di Sacco, A.; Hardwick, K.A.; Blakesley, D.; Brancalion, P.H.S.; Breman, E.; Cecilio Rebola, L.; Chomba, S.; Dixon, K.; Elliott, S.; Ruyonga, G.; et al. Ten golden rules for reforestation to optimize carbon sequestration, biodiversity recovery and livelihood benefits. Glob. Chang. Biol. 2021, 27, 1328–1348. [Google Scholar] [CrossRef]
- MacKenzie, W.H.; Mahony, C.R. An Ecological approach to climate change-informed tree species selection for reforestation. For. Ecol. Manag. 2021, 481, 118705. [Google Scholar] [CrossRef]
- Lampela, M.; Jauhiainen, J.; Sarkkola, S.; Vasander, H. Promising native tree species for reforestation of degraded tropical peatlands. For. Ecol. Manag. 2017, 394, 52–63. [Google Scholar] [CrossRef]
- Jensen, D.A.; Rao, M.; Zhang, J.; Grøn, M.; Tian, S.; Ma, K.; Svenning, J.-C. The potential for using rare, native species in reforestation—A case study of yews (Taxaceae) in China. For. Ecol. Manag. 2021, 482, 118816. [Google Scholar] [CrossRef]
- Nunes, S.; Gastauer, M.; Cavalcante, R.B.L.; Ramos, S.J.; Caldeira, C.F.; Silva, D.; Rodrigues, R.R.; Salomão, R.; Oliveira, M.; Souza-Filho, P.W.M.; et al. Challenges and opportunities for large-scale reforestation in the Eastern Amazon using native species. For. Ecol. Manag. 2020, 466, 118120. [Google Scholar] [CrossRef]
- Sudrajat, D.J.; Rustam, E. Reforestation by direct seeding of Gmelina arborea using seed briquettes: Composition, size and site preparation, and sowing date. IOP Conf. Ser. Earth Environ. Sci. 2020, 533, 12014. [Google Scholar] [CrossRef]
- Casas, J.V.; Tulod, A.M.; Aribal, L.G.; Patricio, J.H.P. Exotic Gmelina arborea Roxb. plantation supports better understory plant diversity than native Nauclea orientalis (L.) plantation 30 years after their establishment in a watershed area in Southern Philippines. J. Biodivers. Environ. Sci. 2017, 11, 134–147. [Google Scholar]
- Tang, L.; Ang, L.; Ho, W. The potential of Acacia mangiun × Acacia auriculiformis hybrid as an afforestation species for impoverished sand tailings. Int. J. Agric. For. Plant. 2016, 4, 68–71. [Google Scholar]
- Permadi, D.B.; Burton, M.; Pandit, R.; Walker, I.; Race, D. Which smallholders are willing to adopt Acacia mangium under long-term contracts? Evidence from a choice experiment study in Indonesia. Land Use Policy 2017, 65, 211–223. [Google Scholar] [CrossRef]
- Moro, M.F.; Castro, A.S.F. A check list of plant species in the urban forestry of Fortaleza, Brazil: Where are the native species in the country of megadiversity? Urban Ecosyst. 2015, 18, 47–71. [Google Scholar] [CrossRef]
- Richardson, D.M.; Le Roux, J.J.; Wilson, J.R.U. Australian acacias as invasive species: Lessons to be learnt from regions with long planting histories. South. For. J. For. Sci. 2015, 77, 31–39. [Google Scholar] [CrossRef]
- Silva, L.D.D.; Lima, A.P.L.; de Lima, S.F.; Silva, R.C.; Paniago, G.F.; Silva, L.D.D.; Lima, A.P.L.; de Lima, S.F.; Silva, R.C.; Paniago, G.F. Controlled-release Fertilizer in the Production and Quality of Acacia mangium Seedlings. Floresta Ambiente 2019, 26. [Google Scholar] [CrossRef]
- Liu, T.; Jiang, K.; Tan, Z.; He, Q.; Zhang, H.; Wang, C. A Method for Performing Reforestation to Effectively Recover Soil Water Content in Extremely Degraded Tropical Rain Forests. Front. Ecol. Evol. 2021, 9, 19. [Google Scholar] [CrossRef]
- Moreira da Silva, A.P.; Schweizer, D.; Rodrigues Marques, H.; Cordeiro Teixeira, A.M.; Nascente dos Santos, T.V.M.; Sambuichi, R.H.R.; Badari, C.G.; Gaudare, U.; Brancalion, P.H.S. Can current native tree seedling production and infrastructure meet an increasing forest restoration demand in Brazil? Restor. Ecol. 2017, 25, 509–515. [Google Scholar] [CrossRef]
- Jalonen, R.; Valette, M.; Boshier, D.; Duminil, J.; Thomas, E. Forest and landscape restoration severely constrained by a lack of attention to the quantity and quality of tree seed: Insights from a global survey. Conserv. Lett. 2018, 11, e12424. [Google Scholar] [CrossRef]
- Rachmat, H.H.; Subiakto, A.; Susilowati, A. Mass vegetative propagation of rare and endangered tree species of Indonesia by shoot cuttings by KOFFCO method and effect of container type on nursery storage of rooted cuttings. Biodiversitas 2018, 19, 2353–2358. [Google Scholar] [CrossRef]
- Lu, Y.; Ranjitkar, S.; Xu, J.; Ou, X.; Zhou, Y.; Ye, J.-F.; Wu, X.-F.; Weyerhaeurser, H.; He, J. Propagation of Native Tree Species to Restore Subtropical Evergreen Broad-Leaved Forests in SW China. Forest 2016, 7, 12. [Google Scholar] [CrossRef]
- Nichols, J.; Vanclay, J. Domestication of native tree species for timber plantations: Key insights for tropical island nations. Int. For. Rev. 2012, 14, 402–413. [Google Scholar] [CrossRef][Green Version]
- Luna-Nieves, A.L.; Meave, J.A.; Morellato, L.P.C.; Ibarra-Manríquez, G. Reproductive phenology of useful Seasonally Dry Tropical Forest trees: Guiding patterns for seed collection and plant propagation in nurseries. For. Ecol. Manag. 2017, 393, 52–62. [Google Scholar] [CrossRef]
- Schmidt, I.B.; de Urzedo, D.I.; Piña-Rodrigues, F.C.M.; Vieira, D.L.M.; de Rezende, G.M.; Sampaio, A.B.; Junqueira, R.G.P. Community-based native seed production for restoration in Brazil—The role of science and policy. Plant Biol. 2019, 21, 389–397. [Google Scholar] [CrossRef]
- Ashton, P.; Kettle, C.J. Dipterocarp Biology as a Window to the Understanding of Tropical Forest Structure: Where are we Looking Now? Biotropica 2012, 44, 575–576. [Google Scholar] [CrossRef]
- Hiroshi, A.; Nakai, Y.; Hasegawa, G.P. Economic importance of the endemic Sumatran lowland dipterocarp tree species (Shorea javanica). Afr. J. Trop. Agric. 2015, 3, 163–172. [Google Scholar]
- Symington, C.F. Malayan Forest Records No. 16, Foresters’ Manual of Dipterocarps; Universiti Malaya: Kuala Lumpur, Malysia, 1974. [Google Scholar]
- Brearley, F.Q.; Banin, L.F.; Saner, P. The ecology of the Asian dipterocarps. Plant Ecol. Divers. 2016, 9, 429–436. [Google Scholar] [CrossRef]
- Lestari, D.A.; Fiqa, A.P.; Fauziah, F.; Budiharta, S. Growth evaluation of native tree species planted on post coal mining reclamation site in East Kalimantan, Indonesia. Biodiversitas 2019, 20, 134–143. [Google Scholar] [CrossRef]
- Pamoengkas, P.; Rachmat, H.H.; Khalifa, N. The growth of shorea leprosula at various planting distances and slopes in gunung dahu research forest, Bogor, Indonesia. Biodiversitas 2020, 21, 4396–4404. [Google Scholar] [CrossRef]
- Yuwati, T.W.; Rachmanadi, D.; Qirom, M.A.; Santosa, P.B.; Kusin, K.; Tata, H.L. Peatland Restoration in Central Kalimantan by Rewetting and Rehabilitation with Shorea balangeran. In Tropical Peatland Eco-management; Osaki, M., Tsuji, N., Foead, N., Rieley, J., Eds.; Springer: Singapore, 2021; pp. 595–611. ISBN 978-981-33-4654-3. [Google Scholar]
- Daisuke, H.; Tanaka, K.; Joseph Jawa, K.; Ikuo, N.; Katsutoshi, S. Rehabilitation of degraded tropical rainforest using dipterocarp trees in Sarawak, Malaysia. Int. J. For. Res. 2013, 2013, 683017. [Google Scholar] [CrossRef]
- Ådjers, G.; Hadengganan, S.; Kuusipalo, J.; Nuryanto, K.; Vesa, L. Enrichment planting of dipterocarps in logged-over secondary forests: Effect of width, direction and maintenance method of planting line on selected Shorea species. For. Ecol. Manag. 1995, 73, 259–270. [Google Scholar] [CrossRef]
- Shono, K.; Davies, S.J.; Chua, Y.K. performance of 45 native tree species on degraded lands in Singapore. J. Trop. For. Sci. 2007, 19, 25–34. [Google Scholar]
- Hassan, A.; Wahab, R.; Alias, M.A.; Salim, R.M. Growth performance of 9-years-old selected 5 indigenous wood species planted on degraded forest land. Int. J. Agric. Res. 2007, 2, 302–306. [Google Scholar] [CrossRef]
- Subiakto, A.; Rachmat, H.H.; Sakai, C. Choosing native tree species for establishing man-made forest: A new perspective for sustainable forest management in changing world. Biodiversitas 2016, 17, 620–625. [Google Scholar] [CrossRef]
- Rachmat, H.H.; Pamoengkas, P.; Sholihah, L.; Fambayun, R.A.; Susilowati, A. The effect of planting technique on the growth of two Shorea species in Gunung Dahu, Bogor, Indonesia. Biodiversitas 2020, 21, 4131–4138. [Google Scholar] [CrossRef]
- Wu, Q. GIS and Remote Sensing Applications in Wetland Mapping and Monitoring. In Comprehensive Geographic Information Systems; Huang, B., Ed.; Elsevier: Oxford, UK, 2018; pp. 140–157. [Google Scholar]
- Maulidah, I.; Pamoengkas, P.; Rachmat, H.H. Natural Regeneration Potential of Shorea Pinanga Scheff and Shorea Platyclados Slooten Ex Endert. in Gunung Dahu Research Forest, Bogor-West Java. IOP Conf. Ser. Earth Environ. Sci. 2019, 394, 012063. [Google Scholar] [CrossRef]
- Nurkhaeni, W. Potensi Regenerasi Alami Shorea leprosula Miq. Di Hutan Penelitian Gunung Dahu, Leuwiliang, Kabupaten Bogoritle, Bogor; IPB University: Bogor, Indonesia, 2019. [Google Scholar]
- Wang, W.; Zhong, Z.; Wang, Q.; Wang, H.; Fu, Y.; He, X. Glomalin contributed more to carbon, nutrients in deeper soils, and differently associated with climates and soil properties in vertical profiles. Sci. Rep. 2017, 7, 13003. [Google Scholar] [CrossRef]
- Supriyanto, I.U. Teknik Pengukuran Penutupan Tajuk dan Pembukaan Tajuk Tegakan Dengan Menggunakan Spherical Densiometer; SEAMEO-BIOTROP: Bogor, Indonesia, 2001. [Google Scholar]
- Chotimah, T. Pertumbuhan Meranti (Shorea leprosula Miq) di Hutan Penelitian Gunung Dahu, Leuwiliang, Kabupaten Bogor; Bogor Agricultural University: Bogor, Indonesia, 2020. [Google Scholar]
- Pertiwi, P.R. Pengaruh Kelerengan Terhadap Pertumbuhan Shorea platyclados di Hutan Penelitian Gunung Dahu, Leuwiliang, Kabupaten Bogor; Bogor Agricultural University: Bogor, Indonesia, 2020. [Google Scholar]
- Saputra, N.E. Analisis Sifat Fisik Tanah dan Laju Infiltrasi pada Berbagai Penggunaan Lahan di Hutan Penelitian Gunung Dahu; Bogor Agricultural University: Bogor, Indonesia, 2020. [Google Scholar]
- Nuraini, T. Keanekaragaman Fungi Ektomikoriza di Hutan Penelitian Meranti Gunung Dahu Leuwiliang Kabupaten Bogor; Bogor Agicultural University: Bogor, Indonesia, 2017. [Google Scholar]
- Kasongat, H.; Gofur, M.A. Ponisri Identifikasi dan kenaekargaman jenis jamur ektomikoriza pada hutan jati di Seram bagian timur. Median 2019, 11, 39–46. [Google Scholar] [CrossRef]
- Karmilasanti, K.; Maharani, R. Keanekaragaman Jenis Jamur Ektomikoriza Pada Ekosistem Hutan Dipterokarpa Di Khdtk Labanan, Berau, Kalimantan Timur. J. Penelit. Ekosist. Dipterokarpa 2016, 2, 57–66. [Google Scholar] [CrossRef]
- Brundrett, M.; Bougher, N.; Dell, B.; Grove, T.; Malajczuk, N. Working with Mycorrhizae in Forestry and Agriculture; ACIAR: Canberra, Australia, 1995. [Google Scholar]
- Rachmat, H.H.; Fambayun, R.A. Restorasi Bentang Alam Terdegradasi: Pembelajaran dari Beberapa Kisah Pemulihan Lahan dengan Pengembangan Teknik KOFFCO. In Konservasi KEHATI Skala Demo Plot; Sawitri, R., Ed.; IPB Press: Bogor, Indonesia, 2019; pp. 269–300. [Google Scholar]
- LMDH. Association of Forest Village Community. 2020; Pabangbon Village: Bogor, Indonesia, 2020. [Google Scholar]
- P3H. Gunung Dahu Long-Term Management Plan; Pusat Litbang Hutan: Bogor, Indonesia, 2020. [Google Scholar]
- Widiyatno, W.; Na’iem, M.; Kanzaki, M.; Purnomo, S.; Jatmiko, J. Application of silviculture treatment to support rehabilitation on logged over area (LOA) of tropical rainforest, Central Kalimantan, Indonesia. Int. J. Sustain. Futur. Hum. Secur. 2013, 1, 50–55. [Google Scholar] [CrossRef]
- Soerianegara, I.; Lemmens, R.H.M.J. Plant Resources of South-East Asia No 5(1) Timber Trees: Major Commercial Timbe; Backhuys Publishers: Leiden, The Netherlands, 1995; ISBN 9022010333. [Google Scholar]
- Widiyatno, W.; Soekotjo, S.; Naiem, M.; Hardiwinoto, S.; Purnomo, S. Pertumbuhan meranti (Shorea spp.) pada sistem tebang pilih tanam jalur dengan teknik silvikultur intensif (TPTJ-SILIN). J. Penelit. Hutan Dan Konserv. Alam 2011, 8, 373–383. [Google Scholar] [CrossRef][Green Version]
- Na’iem, M.; Faridah, E. Model of Intensive Enrichment Planting (TPTII), Silviculture System of 93 Evaluation of Silvicultural System Application Activities. In Indonesia’s Dipterocarps Forest Management: A Lesson Learned; Faculty of Forestry GMU and International Tropical Timber Organization: Yogyakarta, Indonesia, 2006; pp. 25–36. [Google Scholar]
- Newman, M.F.; Burgess, P.F.; Whitmore, T.C. Manuals of Dipterocarps for Foresters: Borneo Island Light Hardwoods: Anisoptera, Parashorea, Shorea (Red, White and Yellow Meranti); Royal Botanic Garden Edinburgh and CIFOR: Edinburgh, UK, 1996; ISBN 1-872291-76-7. [Google Scholar]
- Tampubolon, S.; Manurung, T.F.; Latifah, S. Sebaran Tengkawang (Shorea spp.) berdasarkan Fitogeografi pada Hutan Adat Pengajit Desa Sahan Kecamatan Seluas Kabupaten Bengkayang. J. Hutan Lestari 2018, 6, 883–893. [Google Scholar]
- Mindawati, N.; Tiryana, T. Pertumbuhan Jenis Pohon Khaya anthotheca di Jawa Barat. Bull. Penelit. Hutan 2002, 632, 47–58. [Google Scholar]
- Pamoengkas, P.; Prasetia, R. Pertumbuhan Meranti Merah (Shorea leprosula Miq) Dalam Sistem. Silvikultur Tebang Pilih Tanam Jalur (Studi Kasus di Areal. IUPHHK-HA PT. Sari Bumi Kusuma, Kalimantan Tengah). J. Silvikultur Trop. 2014, 5, 174–180. [Google Scholar]
- Van Gardingen, P.R.; McLeish, M.J.; Phillips, P.D.; Fadilah, D.; Tyrie, G.; Yasman, I. Financial and ecological analysis of management options for logged-over Dipterocarp forests in Indonesian Borneo. For. Ecol. Manag. 2003, 183, 1–29. [Google Scholar] [CrossRef]
- Marsoem, S.N. Studi Mutu Kayu Jati di Hutan Rakyat Gunungkidul I. Pengukuran Laju Pertumbuhan. J. Ilmu Kehutan. 2015, 7, 108–122. [Google Scholar]
- Kuswandi, R. Julius Dwi Nugroho Diameter Increment of Remnant Stands in Logged-Over Forest in Papua. J. Wasian 2019, 6, 125–133. [Google Scholar] [CrossRef]
- Lal, A.B. Silvicultural Systems and Forest Management; Jugal Kishore and Co.: Dehradun, India, 1961. [Google Scholar]
- Anderson, J. Illipe Nuts (Shorea spp.) as Potential Agricultural Crops. In Proceedings of the Symposium on South East Asian Plant Genetic Resources; Williams, J.T., Lamoureux, C.H., Soetjipto, N.W., Eds.; International Board for Plant Genetic Resources: Bogor, Indonesia, 1975; pp. 217–230. [Google Scholar]
- Weinland, G. Plantation. In A Review of Dipterocarps: Taxonomy, Ecology and Silviculture; Appanah, S., Turnbull, J.W., Eds.; CIFOR: Bogor, Indonesia, 1998; pp. 151–186. [Google Scholar]
- Ordonez, J.C.; Luedeling, E.; Kindt, R.; Tata, H.L.; Harja, D.; Jamnadass, R.; van Noordwijk, M. Constraints and opportunities for tree diversity management along the forest transition curve to achieve multifunctional agriculture. Curr. Opin. Environ. Sustain. 2014, 6, 54–60. [Google Scholar] [CrossRef]
- Ashton, P.S. Dipterocarpaceae. In Flora Malesiana I; van Steenis, C.G.G.J., Ed.; Martinus Nijhoff: Hague, The Netherland, 1982; pp. 237–552. [Google Scholar]
- Krishnapillay, B.; Tompsett, P.B. Seed Handling. In A Review of Taxonomy, Ecology and Silviculture; Appanah, S., Turnbull, J., Eds.; CIFOR: Bogor, Indonesia, 1998; pp. 80–88. [Google Scholar]
- Numata, S.; Yasuda, M.; Okuda, T.; Kachi, N.; Noor, N.S.M. Temporal and spatial patterns of mass flowerings on the Malay Peninsula. Am. J. Bot. 2003, 90, 1025–1031. [Google Scholar] [CrossRef]
- Fitriasari, D.A. Evaluasi Hasil Studi Pengaruh Inokulasi Fungi Ektomikoriza Terhadap Respon Pertumbuhan Bibit Shorea spp. di Persemaian; Bogor Agricultural University: Bogor, Indonesia, 2011. [Google Scholar]
- Wahyudi, A.; Sari, N.; Saridan, A. Ekologi, Morfologi & Upaya Konservasi. In Shorea Leprosula Miq dan Shorea Johorensis Foxw: Ekologi, Silvikultur, Budidaya dan Pengembangan; Balai Besar Penelitian Dipterokarpa: Samarinda, Indonesia, 2014; pp. 3–13. ISBN 978-602-9096-13-2. [Google Scholar]
- Fajri, M.; Fernandes, A. Harvesting Patterns of Tengkawang (Shorea machrophylla) and Its Natural Regeneration in Community’s Garden. J. Penelit. Ekosist. Dipterokarpa 2015, 1, 81–88. [Google Scholar] [CrossRef][Green Version]
- Hilwan, I.; Handayani, E.P. Keanekaragaman Mesofauna dan Makrofauna Tanah pada Areal Bekas Tambang Timah di Kabupaten Belitung, Provinsi Kepulauan Bangka-Belitung. J. SIlvikultur Trop. 2013, 4, 35–41. [Google Scholar]
- Panjaitan, S. Pertumbuhan dan komposisi jenis permudaan alam pada rumpang tebangan di Kalimantan Selatan. J. Penelit. Dipterokarpa 2013, 7, 63–74. [Google Scholar] [CrossRef]
- Soekotjo. Teknik Silvikultur Intensif (SILIN); Gadjah Mada University Press: Yogyakarta, Indonesia, 2009. [Google Scholar]
- Hardiwinoto, S.; Adriana, A.; Hadi Nurjanto, H.; Widiyatno, W.; Dhina, F.; Priyo, E. Pengaruh sifat fisika media terhadap kemampuan berakar dan pembentukan akar stek pucuk S. platyclados di PT Sari Bumi Kusuma Kalimantan Tengah. J. Pemuliaan Tanam. Hutan 2010, 4, 37–47. [Google Scholar] [CrossRef]
- Holilullah, H.; Afandi, A.; Novpriansyah, H. Karakteristik sifat fisik tanah pada lahan produksi rendah dan tinggi di PT Great Giant Pinapple. J. Agrotek Trop. 2015, 3, 278–282. [Google Scholar]
- Wong, M. Impact of dipterocarp seedlings on the vegetative and reproductive characteristics of Labisia Pumila (Myrsinaceae) in the understorey. Malays. For. 1981, 44, 370–376. [Google Scholar]
- Kusneti, M. The effect of Phrynium parvum Ridley (Marantaceae) on the growth and density of Dipterocarpus sublamellatus seedling. In Forest Biology and Conservation in Borneo; Murtedza, M., Omar, S., Eds.; Yayasan Sabah: Kinabalu, Malaysia, 1992; pp. 339–347. [Google Scholar]
- Mashudi, M.; Pudjiono, S.; Rayan, R.; Sulaeman, M. Pengaruh asal populasi dan pohon induk terhadap pertumbuhan bibit meranti tembaga Shorea leprosula Miq. sebagai materi untuk perbanyakan klonal. J. Penelit. Dipterokarpa 2012, 6, 97–109. [Google Scholar] [CrossRef][Green Version]
- Hardjowigeno, H.S. Klasifikasi Tanah dan Pedogenesis; Akademika Pressindo: Jakarta, Indonesia, 2003; ISBN 979-8035-48-8. [Google Scholar]
- Darmawijaya, M.I. Klasifikasi Tanah: Dasar Teori Bagi Peneliti Tanah dan Pelaksana Pertanian di Indonesia; Gadjah Mada University Press: Yogyakarta, Indonesia, 2005; ISBN 9794201537. [Google Scholar]
- Stothoff, S.A.; Or, D.; Groeneveld, D.P.; Jones, S.B. The effect of vegetation on infiltration in shallow soils underlain by fissured bedrock. J. Hydrol. 1999, 218, 169–190. [Google Scholar] [CrossRef]
- Aini, Z.Z.; Khasanah, N.; Kusuma, Z. Degradasi sifat fisik tanah sebagai akibat alih guna lahan hutan menjadi sistem kopi monokultur: Kajian perubahan makroporositas tanah. Agrivita 2004, 26, 60–68. [Google Scholar]
- Winanti, T. Pekarangan Sebagai Media Peresapan Air Hujan Dalam Upaya PengelolaanSumberdaya Air. In Proceedings of the Konferensi Nasional Pusat Studi Lingkungan BKPSL; Udayana University: Bali, Indonesia, 1996. [Google Scholar]
- Li, X.; Li, J. Distribution and Conservation Strategy of Eight Threatened Gymnosperm Species in China; China Academic of Science: Wuhan, China, 2004; pp. 1–5. [Google Scholar]
- Mindawati, N. Pratiwi Produksi dan laju dekomposisi serasah Acacia mangium Willd. Bul. Penelit. Hutan 2008, 6, 65–77. [Google Scholar]
- Haron, K.; Brookes, P.C.; Anderson, J.M.; Zakaria, Z.Z. Microbial biomass and soil organic matter dynamics in oil palm (Elaeis guineensis jacq.) plantations, West Malaysia. Soil Biol. Biochem. 1998, 30, 547–552. [Google Scholar] [CrossRef]
- Khasanah, N.; van Noordwijk, M.; Ningsih, H.; Rahayu, S. Carbon neutral? No change in mineral soil carbon stock under oil palm plantations derived from forest or non-forest in Indonesia. Agric. Ecosyst. Environ. 2015, 211, 195–206. [Google Scholar] [CrossRef]
- Brahene, S.W.; Owusu-Bennoah, E.; Abekoe, M.K. Dynamics of soil carbon sequestration under oil palm plantations of different ages. In Proceedings of the Global Symposium on Soil Organic Carbon; FAO: Rome, Italy, 2017; pp. 1–4. [Google Scholar]
- Alexander, M. Introduction to Soil Microbiology, 2nd ed.; Wiley Eastern Limited: Calcutta, India, 1978. [Google Scholar]
- Stevenson, F. Humus Chemistry: Genesis, Composition, Reactions, 2nd ed.; Willey Interscience: New York, NY, USA, 1994; ISBN 978-0-471-59474-1. [Google Scholar]
- Erizilina, E.; Pamoengkas, P.; Darwo, D. Hubungan sifat fisik dan kimia tanah dengan pertumbuhan meranti merah di khdtk haurbentes. J. Nat. Resour. Environ. Manag. 2019, 9, 68–74. [Google Scholar] [CrossRef]
- Anas, I. Biologi Tanah Dalam Praktek; Pusat Antar Universitas Bioteknologi IPB: Bogor, Indonesia, 1989. [Google Scholar]
- Rao, S.N.S. Mikroorganisme Tanah dan Pertumbuhan Tanaman; UI Press: Jakarta, Indonesia, 1994; ISBN 979-456-124-0. [Google Scholar]
- Breure, A.M. Soil biodiversity: Measurements, Indicators, Threats and soil functions. In Proceedings of the International Conference on Soil and Compost Eco-Biology; SoilAce: Leon, Spain, 2004; pp. 83–96. [Google Scholar]
- Carney, K.M.; Matson, P.A. Plant communities, soil microorganisms, and soil carbon cycling: Does altering the world belowground matter to ecosystem functioning? Ecosystems 2005, 8, 928–940. [Google Scholar] [CrossRef]
- Hölldobler, B.; Wilson, E.O. The Ants; Harvard University Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Wallwork, J.A. Ecology of Soil Animals; Blower, J.G., Ed.; McGraw Hill: London, UK, 1971. [Google Scholar]
- Soepardi, G. Sifat dan Ciri Tanah; IPB Press: Bogor, Indonesia, 1983. [Google Scholar]
- Wezel, A.; Rajot, J.L.; Herbrig, C. Influence of shrubs on soil characteristics and their function in Sahelian agro-ecosystems in semi-arid Niger. J. Arid Environ. 2000, 44, 383–398. [Google Scholar] [CrossRef]
- Nugroho, Y. Analisis Kualitas Lahan Untuk Pengembangan Model Pertanaman Jati (Tectona Grandis L.F) Rakyat di Tropika Basah; Universitas Brawijaya: Kota Malang, Indonesia, 2015. [Google Scholar]
- Borror, D.J.; Triplehorn, C.A.; Ohnson, N.F. Pengenalan Pelajaran Serangga, 6th ed.; Gadjah Mada University Press: Yogyakarta, Indonesia, 1992; ISBN 979-420-237-1. [Google Scholar]
- Sutanto, R. Dasar-Dasar Ilmu Tanah: Konsep dan Kenyataan; Kanisius: Yogyakarta, Indonesia, 2005; ISBN 9792104674. [Google Scholar]
- Hakim, H.; Nyakpa, Y.; Lubis, A.M.; Nugroho, S.G.; Saul, M.R.; Diha, M.A.; Hong, G.B.; Bailey, H.H. Dasar-Dasar Ilmu Tanah; Universitas Lampung: Lampung, Indonesia, 1986. [Google Scholar]
- Kohnke, H. Soil Physics; McGraw-Hill Book Company: New York, NY, USA, 1968. [Google Scholar]
- Ginting, S. Pendugan Laju Infiltrasi Menggunakan Parameter Sifat Tanah Pada Kawasan Berlereng; University Sumatera Utara: Medan, Indonesia, 2009. [Google Scholar]
- Allen, M.F.; Swenson, W.; Querejeta, J.I.; Egerton-Warburton, L.M.; Treseder, K.K. Ecology of Mycorrhizae: A Conceptual Framework for Complex Interactions among Plants and Fungi. Annu. Rev. Phytopathol. 2003, 41, 271–303. [Google Scholar] [CrossRef] [PubMed]
- Baghel, R.K.; Sharma, R.; Pandey, A.K. Activity of acid phosphatase in the ectomycorrhizal fungus Cantharellus tropicalis under controlled conditions. J. Trop. For. Sci. 2009, 21, 218–222. [Google Scholar]
- Bechem, E.E.T.; Alexander, I.J. Phosphorus nutrition of ectomycorrhizal Gnetum africanum plantlets from Cameroon. Plant Soil 2012, 353, 379–393. [Google Scholar] [CrossRef]
- Onguene, N.A.; Kuyper, T.W. Importance of the ectomycorrhizal network for seedling survival and ectomycorrhiza formation in rain forests of south Cameroon. Mycorrhiza 2002, 12, 13–17. [Google Scholar] [CrossRef]
- Whipps, J.M. Prospects and limitations for mycorrhizas in biocontrol of root pathogens. Can. J. Bot. 2004, 82, 1198–1227. [Google Scholar] [CrossRef]
- Dell, B. Role of mycorrhiza fungi in ecosystems. CMU J. 2002, 1, 47–55. [Google Scholar]
- Duñabeitia, M.K.; Hormilla, S.; Garcia-Plazaola, J.I.; Txarterina, K.; Arteche, U.; Becerril, J.M. Differential responses of three fungal species to environmental factors and their role in the mycorrhization of Pinus radiata D. Don. Mycorrhiza 2004, 14, 11–18. [Google Scholar] [CrossRef]
- Ahonen-Jonnarth, U.; Roitto, M.; Markkola, A.M.; Ranta, H.; Neuvonen, S. Effects of nickel and copper on growth and mycorrhiza of Scots pine seedlings inoculated with Gremmeniella abietina. For. Pathol. 2004, 34, 337–348. [Google Scholar] [CrossRef]
- Amornpitak, T.Y.; Taweerat, V.; Morakot, T.; Supapon, C.; Sansnarak, R. Diversity of Ectomycorrhizal Fungi on Dipterocarpaceae in Thailand. J. Biol. Sci. 2006, 6, 1059–1064. [Google Scholar]
- Eyssartier, E.; Stubbe, D.; Walleyn, R.; Verbeken, A. New Records of Cantharellus Species (Basidiomycota, Cantharellaceae) from Malaysian Dipterocarp Rainforest. Fungal Divers. 2009, 36, 57–67. [Google Scholar]
- Reddy, M.S.; Singla, S.; Natarajan, K.; Senthilarasu, G. Pisolithus indicus, a new species of ectomycorrhizal fungus associated with Dipetrocarps in India. Mycologia 2005, 97, 839–843. [Google Scholar] [CrossRef]
- Noor, M.; Saridan, A. Keanekaragaman fungi makro pada tegakan benih Dipterocarpaceae di Taman Nasional Tanjung Puting dan Taman Nasional Sebangau Kalimantan Tengah. J. Penelit. Dipterokarpa 2013, 7, 53–62. [Google Scholar] [CrossRef][Green Version]
- Tata, H.L.; van Noordwijk, M.; Summerbell, R.; Werger, M.J.A. Limited response to nursery-stage mycorrhiza inoculation of Shorea seedlings planted in rubber agroforest in Jambi, Indonesia. New For. 2010, 39, 51–74. [Google Scholar] [CrossRef]
- Santos-Silva, C.; Goncalves, A.; Louro, R. Canopy cover influence on macrofungal richness and sporocarp production in montado ecosystems. Agrofor. Syst. 2011, 82, 149–159. [Google Scholar] [CrossRef]
- Chotimah, T.; Wasis, B.; Rachmat, H.H. Population of Macrofauna, Mesofauna, and Ectomycorrhizae’s Fruit Body at Shorea leprosula stand at Gunung Dahu Forest research. J. Penelit. Hutan Dan Konserv. Alam 2020, 17, 79–98. [Google Scholar] [CrossRef]
- Brearley, F.Q. Ectomycorrhizal associations of the Dipterocarpeace. Biotropica 2012, 44, 637–648. [Google Scholar] [CrossRef]
- Brearley, F.Q.; Saner, P.; Uchida, A.; Burslem, D.F.R.P.; Hector, A.; Nilus, R.; Scholes, J.D.; Egli, S. Testing the importance of a common ectomycorrhizal network for dipterocarp seedling growth and survival in tropical forests of Borneo. Plant Ecol. Divers. 2016, 9, 563–576. [Google Scholar] [CrossRef]
- Suwannasai, N.; Dokmai, P.; Yamada, A.; Watling, R.; Phosri, C. First ectomycorrhizal syntheses between astraeus sirindhorniae and dipterocarpus alatus (Dipterocarpaceae), pure culture characteristics, and molecular detection. Biodiversitas 2020, 21, 231–238. [Google Scholar]
- Weemstra, M.; Peay, K.G.; Davies, S.J.; Mohamad, M.; Itoh, A.; Tan, S.; Russo, S.E. Lithological constraints on resource economies shape the mycorrhizal composition of a Bornean rain forest. New Phytol. 2020, 228, 253–268. [Google Scholar] [CrossRef]
- Cuenca, G.; Lovera, M. Seasonal variation and distribution at different soil depths of arbuscular mycorrhizal fungi spores in a tropical sclerophyllous shrubland. Botany 2010, 88, 54–64. [Google Scholar] [CrossRef]
- Liu, J.; Kim, Y.S.; Richardson, C.E.; Tom, A.; Ramakrishnan, C.; Birey, F.; Katsumata, T.; Chen, S.; Wang, C.; Wang, X.; et al. Genetically targeted chemical assembly of functional materials in living cells, tissues, and animals. Science 2020, 367, 1372–1376. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Malajczuk, N.; Brundrett, M.; Dell, B. Fruiting of putative ectomycorrhizal fungi under blue gum [Eucalyptus globulus plantations of different ages in Western Australia. Mycorrhiza 1999, 8, 225–261. [Google Scholar] [CrossRef]
- Lee, S.; Watling, R.; Sikin, Y.N. Ectomycorrhizal basidiomata fruiting in lowland rain forests of Peninsular Malaysia. Bois Des Trop. 2002, 274, 33–43. [Google Scholar]
- Ontl, T.A.; Janowiak, M.K.; Swanston, C.W.; Daley, J.; Handler, S.; Cornett, M.; Hagenbuch, S.; Handrick, C.; McCarthy, L.; Patch, N. Forest management for carbon sequestration and climate adaptation. J. For. 2020, 118, 86–101. [Google Scholar] [CrossRef]
- Zam, S.I. In vitro Bioremediation of Dirtied Soil by Oil Refinery Waste in different pH Concentration) Syukria Ikhsan Zam. J. Agroteknologi 2011, 1, 1–7. [Google Scholar]
- Djaenudin, D. Laporan Hasil Kajian Kontribusi Sosial Ekonomi Hutan Penelitian Gunung Dahu; Pusat Litbang Hutan: Bogor, Indonesia, 2019. [Google Scholar]
- Vigiak, O.; Ribolzi, O.; Pierret, A.; Valentin, C.; Sengtaheuanghoung, O.; Noble, A. Filtering of water pollutants by riparian vegetation: Bamboo versus native grasses and rice in a Lao catchment. Unasylva 2007, 58, 11–16. [Google Scholar]
- Dosskey, M.G.; Vidon, P.; Gurwick, N.P.; Allan, C.J.; Duval, T.P.; Lowrance, R. The Role of Riparian Vegetation in Protecting and Improving Chemical Water Quality in Streams1. J. Am. Water Resour. Assoc. 2010, 46, 261–277. [Google Scholar] [CrossRef]
- Luqman, M.; Butt, T.M.; Tanvir, A.; Atiq, M.; Hussan, M.Z.Y.; Yaseen, M. Phytoremediation of polluted water by trees: A review. Afr. J. Agric. Res. 2013, 8, 1591–1595. [Google Scholar]
- Trabucco, A.; Zomer, R.J.; Bossio, D.A.; van Straaten, O.; Vercot, L.V. Climate change mitigation through afforestation/reforestation: Aglobal analysis of hydrologic impacts with four case studies. Agric. Ecosyst. Environ. 2008, 126, 81–97. [Google Scholar] [CrossRef]
- Ilstedt, U.; Malmer, A.; Verbeeten, E.; Murdiyarso, D. The effect of afforestation on water infiltration in the tropics: A systematic review and meta-analysis. For. Ecol. Manag. 2007, 251, 45–51. [Google Scholar] [CrossRef]
- Bargués Tobella, A.; Reese, H.; Almaw, A.; Bayala, J.; Malmer, A.; Laudon, H.; Ilstedt, U. The effect of trees on preferential flow and soil infiltrability in an agroforestry parkland in semiarid Burkina Faso. Water Resour. Res. 2014, 50, 3342–3354. [Google Scholar] [CrossRef]
- Chazdon, R.L.; Wilson, S.J.; Brondizio, E.; Guariguata, M.R.; Herbohn, J. Key challenges for governing forest and landscape restoration across different contexts. Land Use Policy 2021, 104, 104854. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Spacing Distance (m × m) | Planting Techniques | Average DBH (cm) | Average Height (m) | Volume (m3/ha) | DMAI (cm/Year) |
|---|---|---|---|---|---|---|
| Shorea leprosula * | 2 × 2 | Total | 13.7 | 12.5 | 195.9 | 0.81 |
| Shorea leprosula * | 3 × 3 | Total | 19.7 | 14.9 | 215.4 | 1.16 |
| Sorea leprosula * | 4 × 4 | Total | 24.9 | 16.9 | 127.6 | 1.47 |
| Shorea selanica * | 2 × 2 | Total | 13.9 | 13.3 | 181.4 | 0.82 |
| Shorea selanica * | 3 × 3 | Total | 15.2 | 10.8 | 106.0 | 0.89 |
| Shorea selanica * | 4 × 4 | Total | 19.6 | 16.2 | 86.0 | 1.15 |
| Shorea pinanga | 4 × 4 | Mix | 20.0 | 16.5 | 15.9 | 0.90 |
| Shorea macrophylla | 4 × 4 | Mix | 20.0 | 11.7 | 10.2 | 1.00 |
| Shorea stenoptera | 4 × 4 | Mix | 16.6 | 7.1 | 48.6 | 1.00 |
| Shorea platyclados | 4 × 8 | Mix | 43.1 | 23.0 | 220.8 | 2.15 |
| Shorea platyclados | 4 × 4 | Mix | 22.6 | 10.7 | 160.6 | 1.32 |
| Shorea selanica ** | 2 × 2 | Total | 16.4 | 16.8 | 138.2 | 0.75 |
| Shorea leprosula ** | 2 × 2 | Line | 24.7 | 17.1 | 80.9 | 1.11 |
| Shorea palembanica | 4 × 4 | Mix | 17.8 | 7.0 | 32.8 | 1.05 |
| Shorea ovalis | 4 × 4 | Mix | 24.4 | 8.8 | 76.9 | 1.44 |
| Shorea guiso | 4 × 4 | Mix | 16.4 | 7.0 | 31.5 | 0.96 |
| Anisoptera sp. | 4 × 4 | Mix | 20.5 | 12.0 | 74.1 | 1.21 |
| Hopea bancana | 4 × 4 | Mix | 14.4 | 9.5 | 33.7 | 0.84 |
| Shorea balangeran | 4 × 4 | Mix | 22.9 | 10.9 | 83.7 | 1.34 |
| Shorea mecisopteryx | 4 × 4 | Mix | 12.7 | 7.2 | 17.0 | 0.75 |
| Shorea smitiana | 4 × 4 | Mix | 25.3 | 11.3 | 107.0 | 1.49 |
| Hopea gregaria | 4 × 4 | Mix | 21.5 | 11.3 | 77.0 | 1.27 |
| Hopea odorata | 4 × 4 | Mix | 15.7 | 9.3 | 33.5 | 0.92 |
| Vatica sumatrana | 4 × 4 | Mix | 18.7 | 10.3 | 31.9 | 1.10 |
| Shorea multiflora | 4 × 4 | Mix | 12.8 | 9.0 | 21.8 | 0.75 |
| Hopea dryobalanoides | 4 × 4 | Mix | 16.1 | 9.0 | 34.1 | 0.94 |
| Shorea johorensis | 4 × 4 | Mix | 20.3 | 11.2 | 68.1 | 1.19 |
| Shorea parvifolia | 4 × 4 | Mix | 18.6 | 12.6 | 63.8 | 1.09 |
| Shorea ribrera | 4 × 4 | Mix | 15.1 | 10.6 | 35.7 | 0.89 |
| Anisoptera marginata | 4 × 4 | Mix | 10.7 | 6.1 | 10.2 | 0.62 |
| Dryobalanops aromatica | 4 × 4 | Mix | 23.6 | 10.4 | 85.0 | 1.01 |
| Shorea laevis | 4 × 4 | Mix | 13.6 | 7.1 | 19.3 | 0.80 |
| Dryobalanops lanceolata | 4 × 4 | Mix | 14.0 | 6.7 | 19.4 | 0.83 |
| Dryobalanops oblongifolia | 4 × 4 | Mix | 17.4 | 7.2 | 32.1 | 1.03 |
| Parashorea lucida | 4 × 4 | Mix | 11.2 | 6.5 | 12.0 | 0.66 |
| Parashorea aptera | 4 × 4 | Mix | 12.9 | 6.6 | 16.2 | 0.76 |
| Shorea curtisii | 4 × 4 | Mix | 10.5 | 6.2 | 10.1 | 0.62 |
| Shorea palcifera | 4 × 4 | Mix | 11.2 | 7.5 | 13.9 | 0.66 |
| Shorea seminis | 4 × 4 | Mix | 11.0 | 8.0 | 14.2 | 0.64 |
| Species | Plot | Planting Distance (m) | Year of Planting | Year of the First Flowers (Age, Years Old) | Frequency of Flowering Since Planted to 2020 | Number of Potential Parent Trees | Number of Offspring | Soil Texture | Canopy Coverage (%) | SLOPE (%) | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Seedling | Sapling | ||||||||||
| Shorea leprosula | 1 | 2 × 2 | 1997 | 2015 (18) | 4 times | 11 | >7000 | 0 | Loamy sand | 85.96 | 35 |
| 2 | 3 × 3 | 1997 | 2015 (18) | 4 times | 34 | 1751 | 0 | Loamy sand | 67.50 | 20 | |
| 3 | 4 × 4 | 1997 | 2020 (23) | once | 3 | 346 | 0 | Loamy sand | 66.70 | 35 | |
| 5 | 4 × 4 | 1997 | 2014 (17) | 4 times | 27 | 1320 | 1 | Loamy sand | 68.80 | 35 | |
| 6 | 4 × 4 | 1999 | 2015 (16) | 3 times | 5 | 512 | 0 | Loamy sand | 62.50 | 25 | |
| 7 | 4 × 4 | 1999 | 2015 (16) | 4 times | 6 | 562 | 1 | Loamy sand | 57.36 | 20 | |
| 21a | 4 × 4 | 1999 | 2018 (19) | once | 24 | 0 | 0 | Sandy loam | 78.94 | 40 | |
| Shorea pinanga | 5 | 4 × 4 | 1998 | 2016 (18) | 4 times | 40 | 0 | 0 | Sandy clay | 62.82 | 50 |
| 24 | 4 × 4 | 2000 | 2016 (16) | 4 times | 24 | 489 | 2 | Sandy clay | 68.28 | 35 | |
| Shorea platyclados | 04 | 4 × 4 | 1998 | 2014 (16) | annually | 27 | >11,000 | 357 | Sandy clay | 62.82 | 50 |
| 15 | 4 × 4 | 1999 | 2014 (15) | 3 times | 8 | 127 | 14 | Sandy clay | 80.5 | 40 | |
| 20 | 4 × 4 | 1999 | 2016 (17) | annually | 96 | >12,000 | 248 | Sandy clay | 84.92 | 40 | |
| 21e | 4 × 4 | 1999 | 2018 (19) | annually | 13 | 57 | 0 | Sandy clay | 84.66 | 70 | |
| Shorea selanica | 12 | 4 × 4 | 1998 | 2019 (21) | once | 2 | 5 | 0 | Loamy sand | 67.5 | 20 |
| Shorea macrophylla * | - | 4 × 4 | 2000 | 2018 (18) | 2 times | 5 | 0 | 0 | Sandy clay | 82.5 | 40 |
| Shorea stenoptera * | - | 4 × 4 | 2000 | 2018 (18) | 2 times | 6 | 0 | 0 | Sandy clay | 82.5 | 40 |
| No | Slope Classes (%) | Average | MAI of Diameter (cm/Year) | MAI of Height (m/Year) | |
|---|---|---|---|---|---|
| Diameter (cm) | Total Height (m) | ||||
| 1. | Flat (0–<8%) | 36.3 | 16.7 | 1.8 | 0.8 |
| 2. | Gentle/mild (8–<15) | 40.5 | 20.4 | 2.0 | 1.0 |
| 3. | Fairly steep (15–<25%) | 31.0 | 18.6 | 1.5 | 0.9 |
| 4. | Steep (25–45%) | 35.5 | 19.8 | 1.7 | 1.0 |
| 5. | Very steep (>45%) | 31.4 | 17.9 | 1.5 | 0.9 |
| Physical Soil Properties | Vegetation Cover | Assessment Criteria [119] | |
|---|---|---|---|
| S. leprosula | S. selanica | ||
| Bulk Density | 1.29 | 1.26 | High |
| Soil porosity | 51.06 | 52.32 | Good |
| Moisture content | |||
| – pF1 | 49.64 | 50.77 | |
| – pF2 | 48.78 | 49.12 | |
| – pF3 | 40.11 | 41.28 | - |
| – pF4 | 16.47 | 19.10 | |
| Drainage pore | |||
| • Very fast | - | - | |
| • Fast | 3.21 | 2.29 | |
| • Slow | 7.84 | 8.67 | - |
| Permeability | 1.37 | 2.13 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rachmat, H.H.; Ginoga, K.L.; Lisnawati, Y.; Hidayat, A.; Imanuddin, R.; Fambayun, R.A.; Yulita, K.S.; Susilowati, A. Generating Multifunctional Landscape through Reforestation with Native Trees in the Tropical Region: A Case Study of Gunung Dahu Research Forest, Bogor, Indonesia. Sustainability 2021, 13, 11950. https://doi.org/10.3390/su132111950
Rachmat HH, Ginoga KL, Lisnawati Y, Hidayat A, Imanuddin R, Fambayun RA, Yulita KS, Susilowati A. Generating Multifunctional Landscape through Reforestation with Native Trees in the Tropical Region: A Case Study of Gunung Dahu Research Forest, Bogor, Indonesia. Sustainability. 2021; 13(21):11950. https://doi.org/10.3390/su132111950
Chicago/Turabian StyleRachmat, Henti Hendalastuti, Kirsfianti Linda Ginoga, Yunita Lisnawati, Asep Hidayat, Rinaldi Imanuddin, Rizki Ary Fambayun, Kusumadewi Sri Yulita, and Arida Susilowati. 2021. "Generating Multifunctional Landscape through Reforestation with Native Trees in the Tropical Region: A Case Study of Gunung Dahu Research Forest, Bogor, Indonesia" Sustainability 13, no. 21: 11950. https://doi.org/10.3390/su132111950
APA StyleRachmat, H. H., Ginoga, K. L., Lisnawati, Y., Hidayat, A., Imanuddin, R., Fambayun, R. A., Yulita, K. S., & Susilowati, A. (2021). Generating Multifunctional Landscape through Reforestation with Native Trees in the Tropical Region: A Case Study of Gunung Dahu Research Forest, Bogor, Indonesia. Sustainability, 13(21), 11950. https://doi.org/10.3390/su132111950