
Dwarf White Clover Supports Pollinators, Augments Nitrogen in Clover–Turfgrass Lawns, and Suppresses Root-Feeding Grubs in Monoculture but Not in Mixed Swards

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plot Establishment
2.2. Floral Coverage
2.3. Bee Visitation and Bee Assemblages on the Three Clover Types
2.4. Macroinvertebrate Communities in Monocultures and Dicultures of Clover and Tall Fescue
- Vacuum sampling was performed with a gas-powered leaf blower (Troy-Bilt, Cleveland, OH), reversed for suction with a soft mesh paint strainer clamped inside the intake tube to catch arthropods and organic matter. Sampling was done between 11:00 and 15:00 on dry, sunny days. A sample consisted of two parallel transects across a given plot, walking slowly while lightly dragging and guiding the 14-cm diameter opening of the intake tube through the plant canopy. The paint strainers with enclosed arthropods from a given plot were transferred to sealed paper bags and frozen until the invertebrates were sorted under a binocular microscope. The predominant taxa of insects and spiders were identified to family or, in some cases, to feeding guild (e.g., predatory Hemiptera (mostly Geochoridae and Nabidae), predatory Coleoptera (mainly Staphylinidae and Carabidae), parasitic wasps (including Braconidae, Ichneumonidae, and others), or small Diptera (mostly Chloropidae).
- The activity density of epigeal invertebrates was assessed with pitfall traps made from a pair of nested plastic cups (473 mL (16 oz), 9.53 cm (3.75 inch) top diameter; Solo, Lake Forest, IL) set into the ground level with the soil surface. This allowed the inner cup to be lifted and refifilled without disturbing the trap site. There were five traps per plot, a central trap and one halfway along a diagonal from the center toward each corner. Ethylene glycol (2.5 cm (1 inch)) was added to the cups to kill and preserve the captured invertebrates. The captures from each 3-d sample period were pooled within the plots and stored in 90% ethyl alcohol, with specimens sorted and identified as described above.
- Endogeic earthworms (mainly Apporectodea spp.) were sampled by digging two pits (20 × 20 cm, 21 cm deep), approximately 2 m apart within each plot and by hand-sorting to recover all specimens, which were counted and collectively weighed [70]. We sampled grub populations by using a gasoline-powered sod cutter to cut a 1.22 m long strip (46 cm wide, 8 cm deep) lengthwise through the center of each plot. Each strip was broken apart and examined for grubs, which were identified by their rastral patterns [71] and weighed.
2.5. Foliage-Feeding Caterpillar Assays
2.6. Nitrogen Benefits of Clover to Tall Fescue
2.7. Data Analyses
3. Results
3.1. Floral Coverage
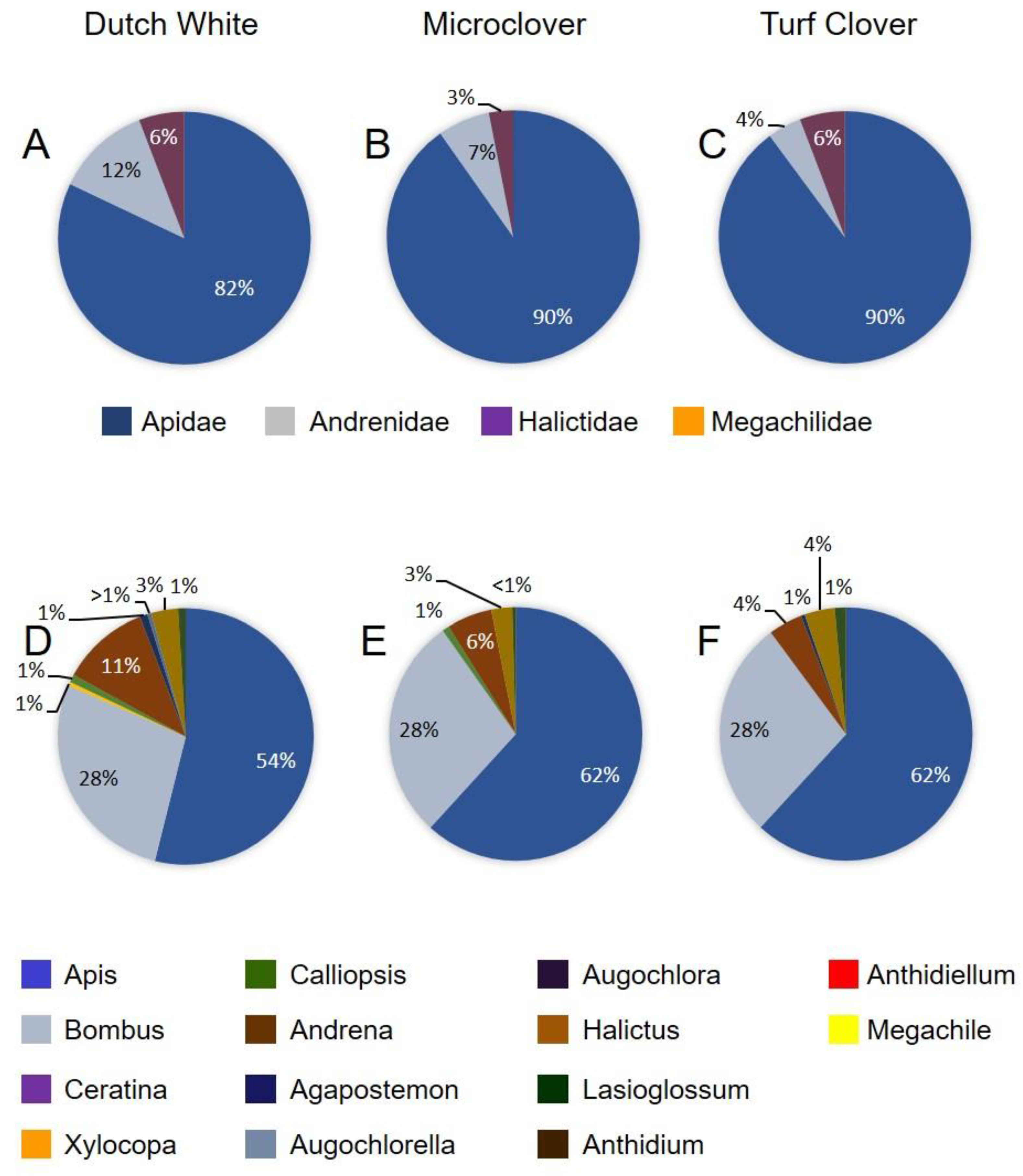
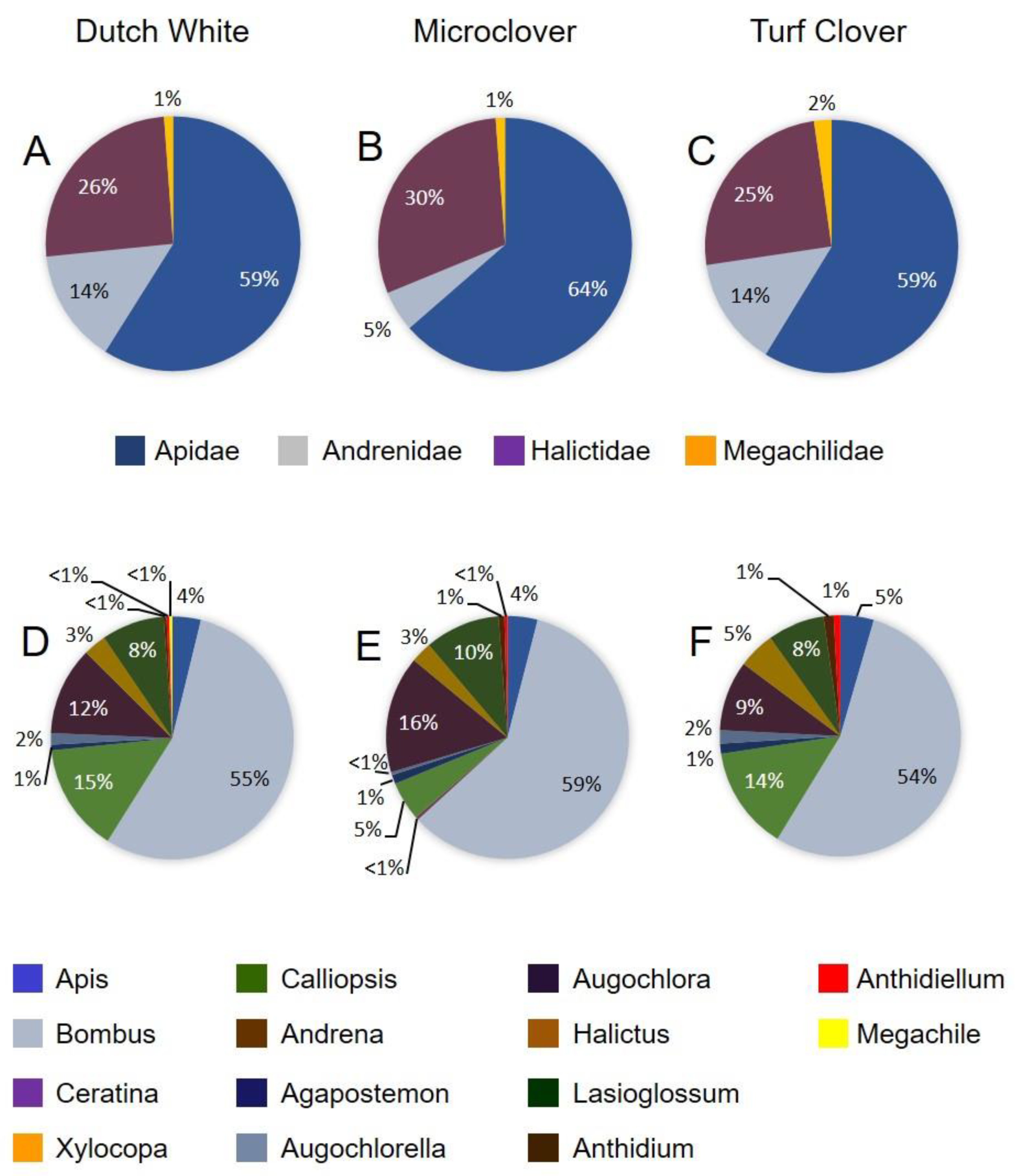
3.2. Bee Visitation and Assemblages on the Three Clover Types
3.3. Macroinvertebrate Communities in Monocultures and Dicultures of Clover and Tall Fescue
3.4. Foliage-Feeding Caterpillar Assays
3.5. Nitrogen Benefits of Clover to Tall Fescue
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ignatieva, M.; Hedblom, M. An alternative green carpet. Science 2018, 362, 148–149. [Google Scholar] [CrossRef] [Green Version]
- Ignatieva, M.; Haase, D.; Dushkovam, D.; Haase, A. Lawns in cities: From a globalized urban green space phenomenon to sustainable nature-based solutions. Land 2020, 9, 73. [Google Scholar] [CrossRef] [Green Version]
- Beard, J.B. Turf Management for Golf Courses, 2nd ed.; Wiley: New York, NY, USA, 2001. [Google Scholar]
- Milesi, C.; Running, S.W.; Elvidge, C.D.; Dietx, J.B.; Tuttle, B.T.; Nemani, R.R. Mapping and modelling the biogeochemical cycling of turfgrass in the United States. Environ. Manag. 2005, 36, 426–438. [Google Scholar] [CrossRef] [PubMed]
- Beard, J.B.; Green, R.L. The roles of turfgrasses in environmental protection and their benefits to humans. J. Environ. Qual. 1994, 23, 452–460. [Google Scholar] [CrossRef] [Green Version]
- Daniels, B.; Zaunbrecher, B.S.; Paas, B.; Ottermanns, R.; Ziefle, M.; Roß-Nickoll, M. Assessment of urban green space structures and their quality from a multidimensional perspective. Sci. Total Environ. 2018, 615, 1364–1378. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, J.A. Ecosystem services from turfgrass landscapes. Urban For. Urban Green. 2017, 26, 151–157. [Google Scholar] [CrossRef]
- Bandaranayake, W.; Qian, Y.; Parton, W.; Ojima, D.; Follett, R. Estimation of soil organic carbon changes in turfgrass systems using the CENTURY model. Agron. J. 2003, 95, 558–563. [Google Scholar] [CrossRef]
- Zirkle, G.; Lal, R.; Augustin, B. Modeling carbon sequestration in home lawns. HortScience 2011, 46, 808–814. [Google Scholar] [CrossRef]
- Selhost, A.; Lal, R. Net carbon sequestration potential and emissions in home lawn turfgrasses of the United States. Environ. Manag. 2013, 51, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Montero, C.A.; Bennett, T.B.; Neville, P.; Crawford, C.S.; Milne, B.T.; Ward, C.R. Potential environmental and economic impacts of turfgrass in Albuquerque, New Mexico (USA). Landscape Ecol. 1995, 10, 121–128. [Google Scholar] [CrossRef]
- Grube, A.; Donaldson, D.; Kiely, T.; Wu, L. Pesticides Industry Sales and Usage: 2006 and 2007 Market Estimates; U.S. Environmental Protection Agency: Washington, DC, USA, 2011.
- Turner, T.R.; Hummel, N.W., Jr. Nutritional requirements and fertilization. In Turfgrass; Waddington, D.W., Carrow, R.N., Shearman, R.C., Eds.; Agronomy Monographs Series; American Society of Agronomy; Crop Science Society of America; Soil Science Society of America: Madison, WI, USA, 2002; Volume 32, pp. 385–439. [Google Scholar]
- Law, N.; Band, L.; Grove, M. Nitrogen input from residential lawn care practices in suburban watersheds in Baltimore County, MD. J. Environ. Plan. Manag. 2004, 47, 737–755. [Google Scholar] [CrossRef]
- US EPA. Polluted Runoff: Nonpoint Source: Urban Areas. 2017. Available online: https://www.epa.gov/nps/nonpoint-source-urban-areas (accessed on 24 October 2021).
- Robertson, G.; Groffman, P. Nitrogen transformations. Soil Microbiol. Ecol. Biochem. 2007, 3, 341–364. [Google Scholar]
- Jørgensen, F.V.; Jensen, E.S.; Schjoerring, J.K. Dinitrogen fixation in white clover grown in pure stand and mixture with ryegrass estimated by the immobilized 15N isotope dilution method. Plant Soil 1999, 208, 293–305. [Google Scholar] [CrossRef]
- Sincik, M.; Acikgoz, E. Effects of white clover inclusion on turf characteristics, nitrogen fixation, and nitrogen transfer from white clover to grass species in turf mixtures. Comm. Soil Sci. Plant Anal. 2007, 38, 1861–1871. [Google Scholar] [CrossRef]
- Sparks, B.; Munshaw, G.; Williams, D.; Barrett, M.; Beasley, J.; Woosley, P. Preplant cultivation techniques and planting date effects on white clover establishment into an existing cool-season turfgrass sward. HortScience 2015, 50, 615–620. [Google Scholar] [CrossRef] [Green Version]
- Frame, J.; Newbould, P. Agronomy of white clover. Adv. Agron. 1986, 40, 1–88. [Google Scholar]
- Larson, J.L.; Kesheimer, A.J.; Potter, D.A. Pollinator assemblages on dandelions and white clover in urban and suburban lawns. J. Insect Conserv. 2014, 18, 863–873. [Google Scholar] [CrossRef]
- MacIvor, J.S.; Cabral, J.M.; Packer, L. Pollen specialization by solitary bees in an urban landscape. Urban Ecosyst. 2014, 17, 139–147. [Google Scholar] [CrossRef]
- Lerman, S.B.; Milam, J. Bee fauna and floral abundance within lawn-dominated suburban yards in Springfield, MA. Ann. Entomol. Soc. Am. 2016, 109, 713–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Heijden, S.A.G.; Roulund, N. Genetic gain in agronomic value of forage crops and turf: A review. In Sustainable Use of Genetic Diversity in Forage and Turf Breeding; Huyghe, S., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 247–260. [Google Scholar]
- Hejduk, S.; Kvasnovský, M. Comparison of white clover cultivars in low input turfs. Eur. J. Turfgrass Sci. 2014, 45, 23–24. [Google Scholar]
- Brilman, L. Reducing nitrogen loss by using microclover in turf and reclamation blends. Land and Water 2016, 60, 37–41. [Google Scholar]
- Blaine, T.W.; Clayton, S.; Robbins, P.; Grewal, P.S. Homeowner attitudes and practices towards residential landscape management in Ohio, USA. Environ. Manag. 2012, 50, 257–271. [Google Scholar] [CrossRef] [PubMed]
- Held, D.W.; Potter, D.A. Prospects for managing turfgrass pests with reduced chemical inputs. Annnu. Rev. Entomol. 2012, 57, 329–354. [Google Scholar] [CrossRef]
- Atwood, D.; Paisley-Jones, C. Pesticides Industry Sales and Usage: 2008–2012 Market Estimates; U.S. Environmental Protection Agency: Washington, DC, USA, 2017.
- Seitz, N.; Traynor, K.S.; Steinhauer, N.; Rennich, K.; Wilson, M.E.; Ellis, J.D.; Rose, R.; Tarpy, D.R.; Sagili, R.R.; Caron, D.M.; et al. A national survey of managed honey bee 2014–2015 annual colony losses in the USA. J. Apicult. Res. 2015, 54, 292–304. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Goulson, D.; Nicholls, E.; Botias, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef] [PubMed]
- Cameron, S.A.; Lozier, J.D.; Strange, J.P.; Koch, J.B.; Cordes, N.; Solter, L.F.; Griswold, T.L. Patterns of widespread decline in North American bumble bees. Proc. Nat. Acad. Sci. USA 2011, 108, 662–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, D.L.; Grames, E.M.; Forister, M.L.; Berenbaum, M.R.; Stopak, D. Insect decline in the Anthropocene: Death by a thousand cuts. Proc. Nat. Acad. Sci. USA 2021, 118, e2023989118. [Google Scholar] [CrossRef]
- Roulson, T.H.; Goodell, K. The role of resources and risks in regulating wild bee populations. Annu. Rev. Entomol. 2011, 56, 293–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tew, N.E.; Memmott, J.; Vaughan, I.P.; Bird, S.; Stone, G.N.; Potts, S.G.; Baldock, C.R. Quantifying nectar production by flowering plants in urban and rural landscapes. J. Ecol. 2021, 109, 1747–1757. [Google Scholar] [CrossRef]
- Kawahara, A.K.; Reeves, L.E.; Barber, J.R.; Black, S.H. Opinion: Eight simple actions that individuals can take to save insects from global declines. Proc. Nat. Acad. Sci. USA 2021, 118, e2002547117. [Google Scholar] [CrossRef]
- National Pollinator Garden Network. Million Pollinator Garden Challenge. Available online: http://millionpollinatorgardens.org (accessed on 24 October 2021).
- Baldock, K.C.R. Opportunities and threats for pollinator conservation in global towns and cities. Curr. Opin. Insect Sci. 2020, 38, 63–71. [Google Scholar] [CrossRef]
- Ramer, H.; Nelson, K.C.; Spivak, M.; Watkins, E.; Wolfin, J.; Pulscher, M. Exploring park visitor perceptions of ‘flowering bee lawns’ in neighborhood parks in Minneapolis, MN, US. Landsc. Urban Plan. 2019, 189, 117–128. [Google Scholar] [CrossRef]
- Lane, I.G.; Wolfin, J.; Watkins, E.; Spivak, M. Testing the establishment of eight forbs in mowed lawns of hard fescue (Festuca brevipila) for use in pollinator conservation. HortScience 2019, 54, 2150–2155. [Google Scholar] [CrossRef] [Green Version]
- Del Toro, I.; Ribbons, R.R. No Mow May lawns have higher pollinator richness and abundances: An engaged community provides floral resources for pollinators. Peer J. 2020, 8, e10021. [Google Scholar] [CrossRef] [PubMed]
- Wolfin, J.; Watkinsm, E.; Lan, E.; Portman, Z.; Spivak, M. Floral enhancement of turfgrass lawns benefits wild bees and hone bees (Apis mellifera). Urban Ecosyst 2021, in press. [Google Scholar]
- Roulston, T.H.; Cane, J.H. Pollen nutritional content and digestibility for animals. Plant Syst. Evol. 2000, 222, 187–209. [Google Scholar] [CrossRef]
- Baude, M.; Kunin, W.E.; Boatman, N.D.; Conyers, S.; Davies, N.; Gillepie, M.A.K.; Morton, R.D.; Smart, S.M.; Memmott, J. Historical nectar assessment reveals the fall and rise of floral resources in Britain. Nature 2016, 530, 85–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sponsler, D.B.; Shump, D.; Richardson, R.T.; Grozinger, C. Characterizing the floral resources of a North American metropolis using a honey bee foraging assay. Ecosphere 2020, 11, e03102. [Google Scholar] [CrossRef]
- Verboven, H.A.F.; Aertsen, W.; Brys, R.; Hermy, M. Pollination and seed set of an obligatory outcrossing plant in an urban–peri-urban gradient. Pers. Plant Ecol. Evol. Syst. 2014, 16, 121–131. [Google Scholar] [CrossRef]
- Kanduth, L.; Chartier, M.; Schönenberger, J.; Dellinger, A.S. Red and white clover provide food resources for honeybees and wild bees in urban environments. Nordic J. Bot. 2021, 39, e03005. [Google Scholar] [CrossRef]
- Vaudo, A.D.; Tooker, J.F.; Grozinger, C.M.; Patch, H.M. Bee nutrition and floral resource restoration. Curr. Opin. Insect Sci. 2015, 10, 133–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timberlake, T.P.; Vaughan, I.P.; Memmott, J. Phenology of farmland floral resources reveals seasonal gaps in nectar availability for bumblebees. J. Appl Ecol. 2019, 56, 1585–1596. [Google Scholar] [CrossRef]
- Hennig, E.I.; Ghazoul, J. Plant–pollinator interactions within the urban environment. Persp. Plant Ecol. Evol. Syst. 2011, 13, 137–150. [Google Scholar] [CrossRef]
- Dover, J.; Settele, J. The influences of landscape structure on butterfly distribution and movement: A review. J. Insect Conserv. 2009, 13, 3–27. [Google Scholar] [CrossRef]
- Dearborn, D.C.; Kark, S. Motivations for conserving urban biodiversity. Conserv. Biol. 2010, 24, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Banaszak-Cibicka, W.; Ratyńska, H.; Dylewski, Ł. Features of urban green space favourable for large and diverse bee populations (Hymenoptera: Apoidea: Apiformes). Urban For. Urban Green. 2016, 20, 448–452. [Google Scholar] [CrossRef]
- Beninde, J.; Veith, M.; Hochkirch, A. Biodiversity in cities needs space: A meta-analysis of factors determining intra-urban biodiversity variation. Ecol. Lett. 2015, 18, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Harrison, T.; Winfree, R. Urban drivers of plant-pollinator interactions. Func. Ecolo. 2015, 29, 879–888. [Google Scholar] [CrossRef]
- Turo, K.J.; Spring, M.R.; Silvakoff, F.S.; Delgado de la flor, Y.A.; Gardiner, M.M. Conservation in post-industrial cities: How does vacant land, management and landscape configuration influence urban bees. J. Appl. Ecol. 2021, 58, 58–69. [Google Scholar] [CrossRef]
- Bewrtoncini, A.P.; Machon, N.; Pavoine, S.; Muratet, A. Local gardening practices shape urban lawn floristic communities. Landsc. Urban Plan 2012, 105, 53–61. [Google Scholar] [CrossRef]
- Lerman, S.B.; Contosta, A.R.; Milam, J.; Bang, C. To mow or to mow less: Lawn mowing frequency affects abundance and diversity in suburban lawns. Biol. Conserv. 2018, 221, 160–174. [Google Scholar] [CrossRef]
- Brosnan, J.; Dickson, K.; Sorochan, J.; Thoms, A.; Stier, J. Large crabgrass, white clover, and hybrid bermudagrass athletic field playing quality in response to simulated traffic. Crop Sci. 2014, 54, 1838–1843. [Google Scholar] [CrossRef]
- Turner, T.; Carroll, M. Microclover-Tall Fescue Lawns in the Mid-Atlantic region. University of Maryland Turfgrass Technical Update TT121. July 2015. Available online: http://www.mdturfcouncil.org/resources/Documents/TT%20Bulletins/TT-121%20Microclover%20_%20Tall%20Fescue%20Lawns%20in%20the%20Mid-Atlantic.pdf (accessed on 24 October 2021).
- NTEP 2019. National Turfgrass Evaluation Program. 2015 National Cool-Season (Low Input) Test. 2019. Available online: https://ntep.org/data/cs15l/cs15l_20-8/cs15lmo119t.txt (accessed on 24 October 2021).
- McCurdy, J.D.; McElroy, J.S.; Guertal, E.A. White clover (Trifolium repens) establishment within dormant bermudagrass turf: Cultural considerations, establishment timing, seeding rate, and cool-season companion grass species. HortScience 2013, 48, 1556–1561. [Google Scholar] [CrossRef] [Green Version]
- Landschoot, P.; Carroll, M. Tolerance of microclover to postemergence and preemergence herbicides. Crop Forag. Turfgrass Manag. 2016, 2, 226. [Google Scholar] [CrossRef]
- Ascher, J.S.; Pickering, J. DiscoverLife Bee Species Guide and World Checklist (Hymenoptera: Apoidea: Anthophila). Available online: https://www.discoverlife.org/mp/20q?search=Apoidea (accessed on 24 October 2021).
- Mitchell, T.B. Bees of the Eastern United States. In North Carolina Agricultural Experiment Station Technical Bulletins; Food and Agriculture Organization of the United Nations: Raleigh, NC, USA, 1960; Volume 141. [Google Scholar]
- Michener, C.D. The Bees of the World; Johns Hopkins University Press: Baltimore, MD, USA, 2007. [Google Scholar]
- LaBerge, W.E. A revision of the bees of the genus Andrena of the Western Hemisphere. Part XIII. Subgenera Simandrena and Taeniandrena. Trans. Am. Entomol. Soc. 1989, 115, 1–56. [Google Scholar]
- Sheffield, C.S.; Ratti, C.; Packer, L.; Griswold, T. Leafcutter and mason bees of the genus Megachile Latreille (Hymenoptera: Megachilidae) in Canada and Alaska. Can. J. Arthropod Iden. 2011, 18, 1–107. [Google Scholar]
- Redmond, C.T.; Kesheimer, A.; Potter, D.A. Earthworm community composition, seasonal population structure, and casting activity on Kentucky golf courses. Appl. Soil Ecol. 2014, 75, 116–123. [Google Scholar] [CrossRef]
- Vittum, P.J. Turfgrass Insects of the United States and Canada, 3rd ed.; Cornell University Press: Ithaca, NY, USA, 2020. [Google Scholar]
- Magurran, A.E. Measuring Biological Diversity; Blackwell: Mauldin, MA, USA, 2004. [Google Scholar]
- Rosenzweig, M.L. Reconciliation ecology and the future of species diversity. Oryx 2003, 37, 194–205. [Google Scholar] [CrossRef] [Green Version]
- Davidson, A.W.; Potter, D.A. Response of plant-feeding, predatory, and soil-inhabiting invertebrates to Acremonium endophyte and nitrogen fertilization in tall fescue turf. J. Econ. Entomol. 1995, 88, 367–379. [Google Scholar] [CrossRef]
- Scriber, J.M.; Slansky, F., Jr. The nutritional ecology of immature insects. Annu. Rev. Entomol. 1981, 26, 183–211. [Google Scholar] [CrossRef]
- Barbosa, P.; Hines, J.; Kaplan, I.; Martinson, H.; Szczepaniec, A.; Szendrei, Z. Associational resistance and associational susceptibility: Having right or wrong neighbors. Annu. Rev. Ecol. Syst. 2009, 40, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Pleasants, M. Bumblebee response to variation in nectar availability. Ecology 1981, 62, 1648–1661. [Google Scholar] [CrossRef]
- Bixby-Brosi, A.; Potter, D.A. Beneficial and innocuous invertebrates in turf. In Handbook of Turfgrass Insect Pests, 2nd ed.; Brandenburg, R., Freeman, C., Eds.; Entomological Society of America: Landham, MD, USA, 2012; pp. 87–93. [Google Scholar]
- Cockfield, S.D.; Potter, D.A. Predatory insects and spiders from suburban lawns in Lexington, Kentucky. Great Lakes Entomol. 1984, 17, 179–184. [Google Scholar]
- Dobbs, E.K.; Potter, D.A. Conservation biological control and pest performance in lawn turf: Does mowing height matter? Environ. Manag. 2014, 53, 648–659. [Google Scholar] [CrossRef] [PubMed]
- Keathley, C.P.; Potter, D.A. Arthropod abundance in tall fescue, Lolium arundinaceum, pastures containing novel ‘safe’ endophytes. J. Appl. Entomol. 2012, 136, 576–587. [Google Scholar] [CrossRef]
- Miller, D.M.; Redmond, C.T.; Flythe, M.D.; Potter, D.A. Evaluation of ‘Jackal’ AR601(Avanex) and Kentucky-31 endophytic tall fescues for suppressing types of invertebrates that contribute to bird strike hazard at airports. Crop Forag. Turfgrass Manag. 2017, 3, 1–11. [Google Scholar] [CrossRef]
- Potter, D.A.; Held, D.W. Biology and management of the Japanese beetle. Annu. Rev. Entomol. 2002, 47, 175–205. [Google Scholar] [CrossRef] [Green Version]
- Fleming, W.E. Biology of the Japanese Beetle; U.S. Department of Agricultur Technical Bulletins: Washington, DC, USA, 1972. [Google Scholar]
- Hervé, M.R.; Erb, M. Distinct defense strategies allow different grassland species to cope with root herbivore attack. Oecologia 2019, 191, 127–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gleadow, R.M.; Møller, B.L. Cyanogenic glycosides: Synthesis, physiology, and phenotypic plasticity. Annu. Rev. Plant Biol. 2014, 65, 155–185. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, T.; Ohta, S.; Ômura, H. A cyanogenic glucoside of Trifolium repens deters oviposition by the common grass yellow Eurema mandarina. Physiol. Entomol. 2019, 44, 222–229. [Google Scholar] [CrossRef]
- Carlsen, S.; Understrup, A.; Fomsgaard, I.; Mortensen, A.; Ravnskov, S. Flavonoids in roots of white clover: Interaction of arbuscular mycorrhizal fungi and a pathogenic fungus. Plant Soil 2008, 302, 33–43. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| September 2015 b | Spring 2016 | ||||
|---|---|---|---|---|---|
| Plot Type a | Composition | Flowers per Plot | Flowers per Plot | Bee Counts per Plot c | |
| 31 May | 20 June | ||||
| Monoculture | Dutch white clover | 35.0 ± 10.9 b | 946 ± 148 a | 3261 ± 269 a | 14.8 ± 1.2 |
| Monoculture | Microclover | 268.0 ± 35.6 a | 820 ± 45 ab | 2136 ± 206 c | 11.4 ± 2.0 |
| Monoculture | Turf Clover | 46.2 ± 15.7 b | 509 ± 37 b | 2908 ± 167 b | 11.2 ± 1.7 |
| Diculture | Dutch white clover + TF | 0 | 139 ± 59 | 440 ± 130 | NA |
| Diculture | Microclover + TF | 0 | 244 ± 43 | 498 ± 75 | NA |
| Diculture | Turf Clover + TF | 0 | 169 ± 42 | 510 ± 64 | NA |
| Monoculture | TF | 0 | 0 | 16 ± 10 d | NA |
| F6,24 (p) all plot types | 30.5 (<0.01) | 98.5 (<0.01) | |||
| F2,8 (p) clover monocultures | 53.8 (<0.001) | 4.79 (0.04) | 31.2 (<0.01) | 1.49 (0.28) | |
| F2,8 (p) clover dicultures | 3.23 (0.09) | 0.54 (0.60) | |||
| Apidae | Turf Clover | Dutch White | Microclover | |||
|---|---|---|---|---|---|---|
| June | August | June | August | June | August | |
| Apis mellifera L. | 128 | 10 | 111 | 10 | 139 | 10 |
| Bombus impatiens Cresson | 45 | 120 | 39 | 145 | 45 | 148 |
| Bombus griseocollis DeGeer | 13 | 1 | 18 | 0 | 19 | 0 |
| Ceratina dupla Say | 0 | 0 | 0 | 0 | 0 | 1 |
| Xylocopa virginica (L.) | 0 | 0 | 1 | 0 | 0 | 0 |
| Andrenidae | ||||||
| Calliopsis andreniformis Smith | 0 | 31 | 2 | 38 | 2 | 13 |
| Andrena sp. | 9 | 0 | 23 | 0 | 13 | 0 |
| Halictidae | ||||||
| Agapostemon virescens (Fab.) | 1 | 3 | 2 | 2 | 0 | 3 |
| Augochlorella aurata (Smith) | 0 | 4 | 1 | 4 | 0 | 1 |
| Augochlora pura Say | 0 | 21 | 0 | 31 | 0 | 39 |
| Halictus rubicundus (Christ) | 5 | 8 | 4 | 5 | 5 | 2 |
| Halictus confusus Smith | 3 | 3 | 3 | 3 | 1 | 5 |
| Lasioglossum sp. | 3 | 17 | 2 | 22 | 1 | 25 |
| Megachilidae | ||||||
| Anthidium manicatum (L.) | 0 | 3 | 0 | 1 | 0 | 2 |
| Anthidiellum notatum (Latreille) | 0 | 2 | 0 | 1 | 0 | 1 |
| Megachile brevis Say | 0 | 0 | 0 | 1 | 0 | 0 |
| Total bees sampled | 207 | 223 | 205 | 263 | 225 | 250 |
| Mean (SE) species richness per plot a | 6.0 (0.3) | 8.4 (0.5) | 6.6 (0.8) | 7.4 (0.7) | 5.4 (0.5) | 7.2 (0.5) |
| Mean (SE) species diversity b | 2.25 (0.16) | 3.06 (0.12) | 2.95 (0.40) | 2.90 (0.20) | 2.30 (0.16) | 2.63 (0.28) |
| Plot Type | Herbivore Type a | |||||
|---|---|---|---|---|---|---|
| Leaf-Hoppers | Frog-Hoppers | Flea Beetles | Aphids | Grass Flies | Slugs | |
| Dutch white clover (M) | 166 ± 21 | 49 ± 16 | 56 ± 10 | 52 ± 23 | 736 ± 72 | 18 ± 2 |
| Microclover (M) | 183 ± 26 | 32 ± 6 | 64 ± 17 | 54 ± 19 | 635 ± 74 | 15 ± 2 |
| Turf Clover (M) | 171 ± 17 | 33 ± 4 | 73 ± 18 | 41 ± 19 | 576 ± 53 | 12 ± 2 |
| Dutch white + TF (D) | 303 ± 62 | 106 ± 13 | 57 ± 7 | 25 ± 11 | 536 ± 160 | 18 ± 3 |
| Micro + TF (D) | 289 ± 33 | 101 ± 13 | 92 ± 31 | 41 ± 16 | 534 ± 79 | 16 ± 4 |
| Turf Clo + TF (D) | 271 ± 34 | 91 ± 28 | 64 ± 9 | 25 ± 12 | 491 ± 114 | 17 ± 2 |
| Tall fescue | 213 ± 42 | 53 ± 18 | 57 ± 11 | 38 ± 11 | 502 ± 135 | 21 ± 5 |
| F7,24 (p) b | 3.7 (<0.01) | 3.8 (<0.01) | 0.54 (0.78) | 0.44 (0.85) | 0.70 (0.65) | 1.0 (0.15) |
| M vs. TF | ns | ns | ns | ns | ns | 1.9 (0.07) |
| M vs. D | 4.6 (<0.01) | 4.6 (<0.01) | ns | ns | ns | ns |
| D vs. TF | 2.1 (<0.05) | 2.5 (<0.05) | ns | ns | ns | ns |
| Plot Type | Predator Type a | |||||
|---|---|---|---|---|---|---|
| Spiders | Rove Beetles | Ground Beetles | Ants | Predatory Bugs | Lady Beetles | |
| Dutch white clover (M) | 185 ±15 | 64 ± 6 | 18 ± 3 | 115 ± 22 | 34 ± 3 | 8 ± 2 |
| Microclover (M) | 191 ± 18 | 81 ± 13 | 22 ± 5 | 120 ± 9 | 34 ± 11 | 9 ± 3 |
| Turf Clover (M) | 221 ± 22 | 77 ± 15 | 17 ± 4 | 112 ± 17 | 32 ± 7 | 12 ± 6 |
| Dutch white + TF (D) | 192 ± 20 | 87 ± 12 | 11 ± 2 | 13 ± 20 | 17 ± 3 | 4 ± 2 |
| Micro + TF (D) | 191 ± 10 | 87 ± 21 | 18 ± 3 | 101 ± 6 | 22 ± 7 | 10 ± 4 |
| Turf Clo + TF (D) | 176 ± 15 | 99 ± 13 | 15 ± 2 | 96 ± 18 | 17 ± 4 | 7 ± 3 |
| Tall fescue | 182 ± 11 | 85 ± 13 | 18 ± 3 | 102 ± 16 | 16 ± 4 | 10 ± 12 |
| F7,24 (p) | 0.77 (0.6) | 0.62 (0.71) | 1.87 (0.13) | 0.28 (0.94) | 1.64 (0.18) | 0.58 (0.74) |
| Plot Type | Earthworms a | P. japonica Grubs b | ||
|---|---|---|---|---|
| Number | Mass (g) per Worm | Number | Mass (mg) per Grub | |
| Dutch white clover (M) | 43 ± 7 | 234 ± 38 | 2.0 ± 1.0 | 127 ± 15 |
| Microclover (M) | 56 ± 5 | 145 ± 14 | 6.2 ± 1.1 | 134 ± 10 |
| Turf Clover (M) | 54 ± 12 | 172 ± 15 | 4.6 ± 1.3 | 124 ± 7 |
| Dutch white + TF (D) | 34 ± 11 | 241 ± 43 | 33.8 ± 5.1 | 176 ± 11 |
| Micro + TF (D) | 40 ± 6 | 187 ± 23 | 36.2 ± 6.8 | 179 ± 4 |
| Turf Clo + TF (D) | 38 ± 7 | 163 ± 22 | 38.5 ± 5.6 | 162 ± 6 |
| Tall fescue | 28 ± 5 | 219 ± 36 | 38.4 ± 9.4 | 191 ± 5 |
| F6,24 (p) | 1.7 (0.18) | 1.6 (0.18) | 10.8 (<0.001) | 12.1 (<0.001) |
| M vs. TF (t1, p) | ns | ns | 5.7 (<0.001) | 7.0 (<0.001) |
| M vs. D (t1, p) | ns | ns | 7.3 (<0.001) | 7.0 (<0.001) |
| D vs. TF (t1, p) | ns | ns | 0.4 (0.68) | 2.2 (0.04) |
| Foliage Type | Week 1 a | Week 2 b | |||
|---|---|---|---|---|---|
| % Survival | Instar Attained | Wt (mg) Attained | Instar Attained | Wt (mg) Attained | |
| Dutch white | 98.8 ± 1.2 | 1.8 ± 0.1 | 13.6 ± 1.7 ab | 3.1 ± 0.1 ab | 163 ± 9 a |
| Microclover | 95.0 ± 2.6 | 1.5 ± 0.1 | 7.5 ± 1.3 b | 2.9 ± 0.1 b | 91 ± 12 b |
| Turf Clover | 88.8 ± 3.5 | 1.6 ± 0.1 | 11.7 ± 2.5 a | 3.3 ± 0.1 a | 144 ± 15 a |
| Tall fescue | 93.8 ± 2.6 | 1.3 ± 0.1 | 5.7 ± 0.5 c | 2.2 ± 0.1 c | 31 ± 3 c |
| F (p) c | 2.49 (0.09) | 2.54 (0.08) | 4.23 (0.02) | 20.2 (<0.01) | 33.5 (0.01) |
| Clovers vs. TF (t1, p) | Ns | 2.19 (0.04) | 2.56 (0.02) | 7.1 (<0.01) | 9.0 (<0.01) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Potter, D.A.; Redmond, C.T.; McNamara, T.D.; Munshaw, G.C. Dwarf White Clover Supports Pollinators, Augments Nitrogen in Clover–Turfgrass Lawns, and Suppresses Root-Feeding Grubs in Monoculture but Not in Mixed Swards. Sustainability 2021, 13, 11801. https://doi.org/10.3390/su132111801
Potter DA, Redmond CT, McNamara TD, Munshaw GC. Dwarf White Clover Supports Pollinators, Augments Nitrogen in Clover–Turfgrass Lawns, and Suppresses Root-Feeding Grubs in Monoculture but Not in Mixed Swards. Sustainability. 2021; 13(21):11801. https://doi.org/10.3390/su132111801
Chicago/Turabian StylePotter, Daniel A., Carl T. Redmond, Timothy D. McNamara, and Gregg C. Munshaw. 2021. "Dwarf White Clover Supports Pollinators, Augments Nitrogen in Clover–Turfgrass Lawns, and Suppresses Root-Feeding Grubs in Monoculture but Not in Mixed Swards" Sustainability 13, no. 21: 11801. https://doi.org/10.3390/su132111801
APA StylePotter, D. A., Redmond, C. T., McNamara, T. D., & Munshaw, G. C. (2021). Dwarf White Clover Supports Pollinators, Augments Nitrogen in Clover–Turfgrass Lawns, and Suppresses Root-Feeding Grubs in Monoculture but Not in Mixed Swards. Sustainability, 13(21), 11801. https://doi.org/10.3390/su132111801