1. Introduction
The European catfish
Silurus glanis (Linnaeus, 1758) is an apex predatory fish species in the freshwater ecosystems of central Europe. Its populations are expanding due to climate change, river damming, and introductions [
1,
2,
3]. Catfish predation may potentially drive freshwater ecosystems and wild fish populations. The predation pressure may be further increased by an intensive catfish stocking that causes unnaturally high predation pressure on local fishes. Conversely, anglers can significantly reduce the catfish populations through intensive angling [
4,
5]. The possible negative effect of catfish predation on local fish populations has been discussed [
1,
2,
3,
5]. While the catfish occupies various habitats, it mostly lives in larger rivers which are the natural habitat of the rheophilic fish species.
Rheophilic fish species are globally threatened organisms [
6]. The rheophilic fishes are a fish species that prefer to live, hunt, and reproduce in fast-moving waters. Their populations are decreasing due to anthropogenic actions—mainly river fragmentation, removal of shelters and spawning sites, water pollution, protection of piscivorous cormorants and otters, and angling pressure [
7,
8,
9,
10]. Despite intensive stocking efforts, a reintroduction of the rheophilic fishes failed to stabilize their populations in central Europe due to river fragmentation and poor environmental conditions [
11]. Conversely, catfish reintroductions were successful even outside of its native habitats—and catfish stocking has become a potential driver of the harvest rates of many prey fish species [
5].
The harvest rates of fish species are mostly driven by fish stocking, angling pressure, angler preferences, fishing gear used, angling restrictions, and environmental conditions [
12,
13,
14,
15]. Strong positive relationships between fish stocking and fish harvesting within the same fish species exist [
13,
16]. In addition, significant negative relationships between the stocking of predatory fish species and the harvest rates of their prey were also reported, showing that the intensive stocking of predators can lead to lower harvest rates of their prey [
17]. Even though multi-species stocking management could lead to different harvest rates in comparison to single species restocking, the analyses of multi-species relationships between stocking and harvest rates are still rare.
Fish stocking is usually performed for two reasons—either to provide fish for angling purposes or to bolster the wild fish populations. For angling purposes, species like the common carp Cyprinus carpio (Linnaeus, 1758), rainbow trout Oncorhynchus mykiss (Walbaum, 1792), or piscivorous fishes are stocked. For the bolstering of the wild populations, naturally occurring fish species that are negatively affected by anthropogenic pressure are stocked. The stocking of multiple species is performed because fisheries managers want to satisfy both the anglers (who fish for fun and food) and the conservationists (who want to stabilize the wild populations). While both reasons for stocking are acceptable (and functioning) in a vacuum, the problem is that stocking is performed on the same rivers simultaneously. When multi-species stocking is done at the wrong intensity and with the wrong mix of species, it can have negative effects on the harvest rates of fishes and the wild fish populations alike. This is because the stocked fish compete for food, shelter, and habitats. In particular, there is a concern that the intensive stocking of catfish could lead to lower harvest rates of the rheophilic fishes because it is performed on the same rivers. The author believes that he can help to partially enlighten this problem by analyzing this very precisely collected data on harvest rates and restocking rates. This data set has several advantages over the data sets used by other studies.
Previous studies either collected the data from a few rivers and streams or over a short period. They also analyzed only one fish species at a time and did not try to build multi-species models [
13,
16,
17]. Therefore, the multi-species interactions between fish harvesting and fish stocking remain unknown. This study aimed to bridge this knowledge gap by newly analyzing the harvesting–stocking relationship between catfish and three rheophilic fishes. The study utilized the precisely collected and relatively worldly unique data on fish harvesting and fish stocking that has been collected by the Czech Fishing Union since 2005 [
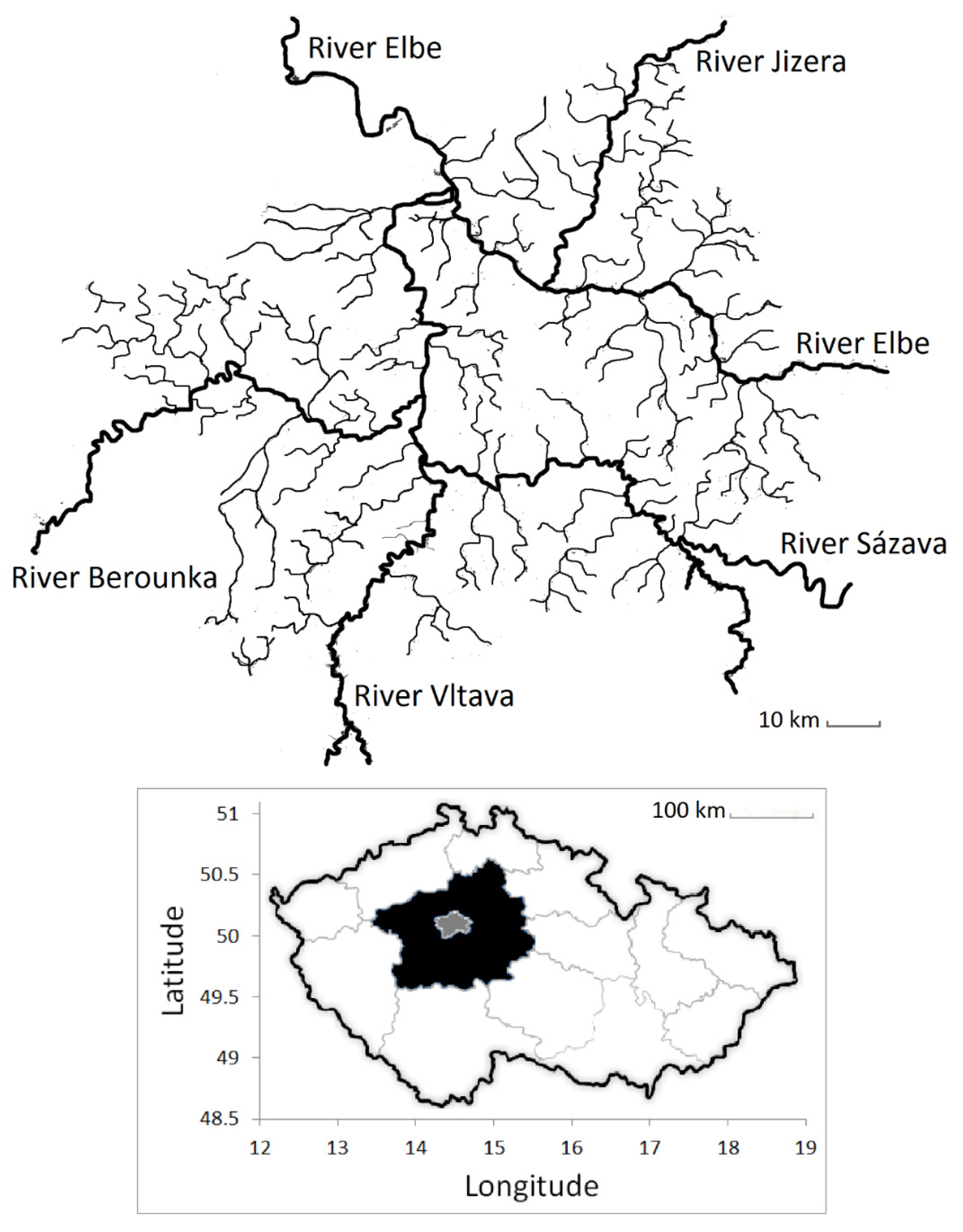
11]. The Union has been collecting the harvest rates and the stocking rates of all four fish species on each individual river and stream in the region, allowing a spatial-temporal comparison of the data.
This study aimed to analyze the relationship between the harvest rate and the stocking intensity of the predator European catfish and the harvest rates of its prey, the rheophilic fish species (Barbus barbus, Chondrostoma nasus, and Vimba vimba-Linneaus, 1758) in lowland mesotrophic rivers in Prague and the Czech Republic in central Europe. It also aimed to analyze the correlation between the harvest rate and an angling effort together with a fishery magnitude. It was expected that the stocking intensity of the rheophilic fishes would be positively correlated with their harvest rates. It was also expected that intensive catfish stocking would be negatively correlated with the harvest rate of rheophilic fishes. It was then expected that intensive catfish harvesting would be negatively correlated with harvest rates of rheophilic fishes. Finally, it was necessary to investigate if the angling effort and the size of a fishery also have a significant relationship with the harvest rates of the rheophilic fishes. It was expected to discover which factors are strongly correlated with the harvest rates of the rheophilic fishes. It was hypothesized that the harvest and stocking intensity of the catfish together with the stocking intensity of the rheophilic fishes would be more important than the effort and the fishery size.
3. Results
The surveyed anglers fished on the studied fishing sites 6.8 million times, and they harvested 107.5 tons of the rheophilic fish and 237 tons of the catfish (
Table A7 in an
Appendix D). The rheophilic fish and the catfish made 0.7% and 2% of the overall fish harvest by biomass, respectively. The anglers harvested 4.8, 3.0, 1.5, and 11.8 times more barbel, nase, vimba bream, and catfish (respectively) than what the fisheries managers stocked.
It was found that the harvest and stocking rates of the catfish were strongly correlated with the harvest rates of barbel, nase, and vimba bream (
Table 1,
Table 2 and
Table 3). A higher intensity of catfish stocking and harvesting resulted in a lower harvest rate of the rheophilic fish. An angling effort also strongly influenced the rheophilic fish harvest rates—each angler harvested less fish on intensively fished rivers. Conversely, the stocking rates of the rheophilic fish did not significantly affect their harvest rates.
Native rheophilic fish species with higher harvest rates showed stronger relationships with the stocking rates and the harvest rates of the catfish. Barbel was the most intensively harvested rheophilic fish species—and showed the strongest relationship with the catfish stocking intensity and its harvest rate (
Table 1). Conversely, nase was the least intensively harvested rheophilic fish species and showed the weakest relationship with the catfish stocking and harvesting (
Table 2). The harvest rate of native species (babel and vimba bream) was more affected by the catfish angling and restocking than the harvest of the non-native nase (
Table 3).
4. Discussion
The harvest rate of the rheophilic fishes was significantly correlated with the stocking intensity and harvest rate of the catfish together with the angling effort. This finding supports the initial hypothesis, although it was expected that the angling effort would be less significantly correlated. The size of the fishery proved to be insignificantly correlated with the harvest rates, which supports the hypothesis. The relationship between the prey harvest rate and the predator stocking rate could be driven by the predator–prey relationship between the catfish and the rheophilic fishes. In central Europe, catfish predation is further combined with predation by otters
Lutra lutra (Linnaeus, 1758), cormorants
Phalacrocorax carbo (Linnaeus, 1758), and gray herons
Ardea Cinerea (Linnaeus, 1758) [
7,
36]. River damming, water pollution (mainly organic pollution and microplastics), and removal of hideouts further pressure the rheophilic fish populations [
8,
9,
10,
21]. The anglers, the fisheries managers, and the representatives of angling clubs are concerned that the catfish expansion threatens the rheophilic fishes [
37]. The managers say that the intensive catfish stocking could directly cause the decreased harvest of the rheophilic fishes. This is because the highly predated fish populations are more timid and harder to exploit by the anglers [
38]. The wild fish populations adapted to the increased predation pressure by hiding which makes them less vulnerable to angling. Conversely, the higher harvest rates of the catfish could be caused by the fact that the hatchery-reared restocked catfish have more aggressive personalities than wild fish which makes them more likely to be predated and to actively seek prey and get harvested [
39,
40]. However, harvest rates are also driven by angling preferences.
The anglers could have switched their angling preferences from the rheophilic fishes to the stocked catfish. The rheophilic fishes are not strongly preferred by the anglers while the catfish is a “tier one” angling target. The anglers have gained access to high-quality fishing gear and a great know-how regarding trophy-sized catfish angling. Catfish angling has become a popular sport in Europe over the last 5–10 years [
3,
4,
19]. Its popularity launched introductions outside of the native range of the catfish, and the catfish became an expansive or even an invasive species [
3,
4,
19]. Angling “selfies” with a catch of a trophy-sized catfish have flooded the angling discussion forums on Facebook and Twitter. The promotion of catfish angling further encourages more anglers to specialize in catfish angling. Conversely, intensive catfish angling can cause a decrease in the local catfish populations which can release the rheophilic fish populations from the catfish predation pressure [
5]. This would explain the significant negative relationship between the harvest rates of the catfish and the rheophilic fish species.
The harvest rate of the rheophilic fishes was not significantly influenced by their stocking management. This result contradicts the initial hypothesis. The rheophilic fishes were likely stocked at a too low an intensity to be significantly harvested by the anglers. The stocked rheophilic fish were also too small to be immediately harvested, due to the relatively high minimum legal angling size of each restocked species. Stocked fish usually have a high post-stocking mortality, a higher vulnerability to predation, low adaptive skills, and an underdeveloped ability to catch wild prey [
41]. Most stocked fish survive in the wild for several months at maximum [
42]. They often have a lower survival rate in comparison to the wild fish [
43] but exceptions to this rule have also been reported [
44].
The angling effort strongly affected the harvest rates of the rheophilic fishes. Previous studies also agreed that the harvest rates of fish are driven by the angling effort [
33,
35,
45,
46]. The angling effort is relatively hard to estimate because each angler fishes at a different intensity. This brings errors to the data analysis.
The angling logbooks provided information on the angling effort and the harvest rates, but they are partially erroneous. The anglers may provide the incorrect size of the killed fish (e.g., they kill a 50 cm large catfish and write down a 70 cm catfish instead to meet the minimum legal angling size), they could incorrectly identify the killed fish (e.g., misinterpreting a nase for a vimba bream), they may not comply with the angling rules (e.g., ignoring the minimum angling size limits), they might prefer a specific fish species (e.g., releasing a caught barbel to make room for a common carp), and they often release the caught fish [
47,
48,
49].
Statistical analyses bring other errors. The analysis of multiple p-values increases the chance to incorrectly reject a null hypothesis. We made 21 statistical analyses at a 0.05 alpha level, meaning that some of the hypotheses were likely incorrectly rejected.




{kind=link}