
Evaluation of Species Invasiveness: A Case Study with Acacia dealbata Link. on the Slopes of Cabeça (Seia-Portugal)




Abstract
:1. Introduction
1.1. Framework
1.2. Characteristics of Acacia Dealbata That Favor Invasion
2. Materials and Methods
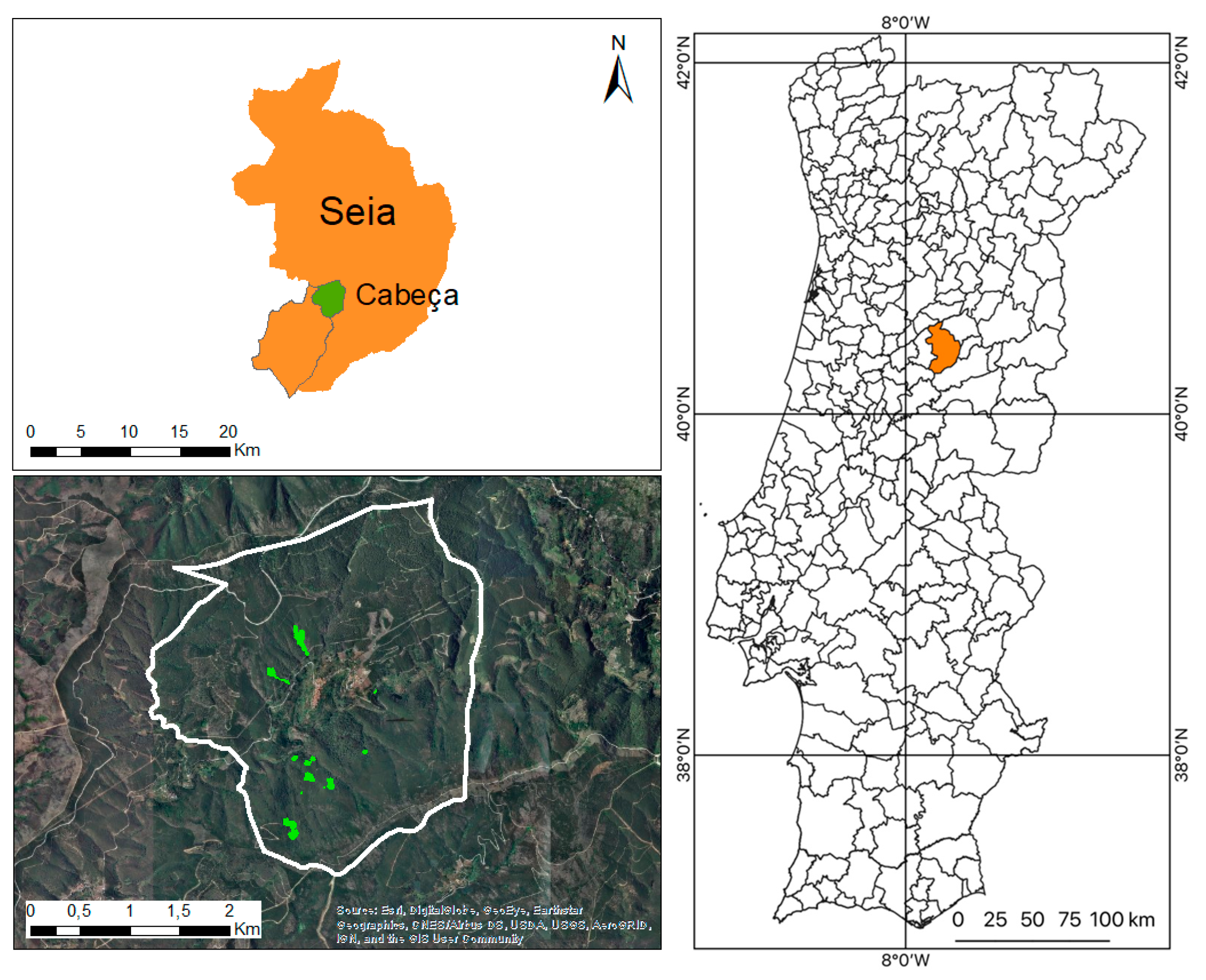
2.1. Characterization of the Area under Study
2.2. Data Collection and Analysis
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, R.J.; Muir, R.D.J.; Walpole, M.J.; Balmford, A.; Leader-Williams, N. Governance and the Loss of Biodiversity. Nature 2003, 426, 67–70. [Google Scholar] [CrossRef]
- Underwood, E.C.; Viers, J.H.; Klausmeyer, K.R.; Cox, R.L.; Shaw, M.R. Threats and Biodiversity in the Mediterranean Biome. Divers. Distrib. 2009, 15, 188–197. [Google Scholar] [CrossRef]
- Khapugin, A.A.; Kuzmin, I.V.; Silaeva, T.B. Anthropogenic Drivers Leading to Regional Extinction of Threatened Plants: Insights from Regional Red Data Books of Russia. Biodivers. Conserv. 2020, 29, 2765–2777. [Google Scholar] [CrossRef]
- Roux, J.J.L.; Hui, C.; Castillo, M.L.; Iriondo, J.M.; Keet, J.-H.; Khapugin, A.A.; Médail, F.; Rejmánek, M.; Theron, G.; Yannelli, F.A.; et al. Recent Anthropogenic Plant Extinctions Differ in Biodiversity Hotspots and Coldspots. Curr. Biol. 2019, 29, 2912–2918.e2. [Google Scholar] [CrossRef]
- Laface, V.L.A.; Musarella, C.M.; Cano Ortiz, A.; Quinto Canas, R.; Cannavò, S.; Spampinato, G. Three New Alien Taxa for Europe and a Chorological Update on the Alien Vascular Flora of Calabria (Southern Italy). Plants 2020, 9, 1181. [Google Scholar] [CrossRef] [PubMed]
- Morais, M.; Marchante, E.; Marchante, H. Big Troubles Are Already Here: Risk Assessment Protocol Shows High Risk of Many Alien Plants Present in Portugal. J. Nat. Conserv. 2017, 35, 1–12. [Google Scholar] [CrossRef]
- Lorenzo, P.; Palomera-Pérez, A.; Reigosa, M.J.; González, L. Allelopathic Interference of Invasive Acacia Dealbata Link on the Physiological Parameters of Native Understory Species. Plant Ecol. 2011, 212, 403–412. [Google Scholar] [CrossRef]
- Rodríguez-Echeverría, S.; Afonso, C.; Correia, M.; Lorenzo, P.; Roiloa, S.R. The Effect of Soil Legacy on Competition and Invasion by Acacia Dealbata Link. Plant Ecol. 2013, 214, 1139–1146. [Google Scholar] [CrossRef]
- Nunes, L.J.R.; Raposo, M.A.M.; Meireles, C.I.R.; Gomes, C.J.P.; Ribeiro, N.M.C.A. Carbon Sequestration Potential of Forest Invasive Species: A Case Study with Acacia Dealbata Link. Resources 2021, 10, 51. [Google Scholar] [CrossRef]
- Nunes, L.J.R.; Raposo, M.A.M.; Meireles, C.I.R.; Pinto Gomes, C.J.; Ribeiro, N.M.C.A. Control of Invasive Forest Species through the Creation of a Value Chain: Acacia Dealbata Biomass Recovery. Environments 2020, 7, 39. [Google Scholar] [CrossRef]
- Nunes, L.J.R.; Raposo, M.A.M.; Meireles, C.I.R.; Gomes, C.J.P.; Ribeiro, N.M.C.A. The Impact of Rural Fires on the Development of Invasive Species: Analysis of a Case Study with Acacia Dealbata Link. in Casal Do Rei (Seia, Portugal). Environments 2021, 8, 44. [Google Scholar] [CrossRef]
- Lazzaro, L.; Bolpagni, R.; Buffa, G.; Gentili, R.; Lonati, M.; Stinca, A.; Acosta, A.T.R.; Adorni, M.; Aleffi, M.; Allegrezza, M.; et al. Impact of Invasive Alien Plants on Native Plant Communities and Natura 2000 Habitats: State of the Art, Gap Analysis and Perspectives in Italy. J. Environ. Manag. 2020, 274, 111140. [Google Scholar] [CrossRef] [PubMed]
- Maitre, D.C.L.; Gaertner, M.; Marchante, E.; Ens, E.-J.; Holmes, P.M.; Pauchard, A.; O’Farrell, P.J.; Rogers, A.M.; Blanchard, R.; Blignaut, J.; et al. Impacts of Invasive Australian Acacias: Implications for Management and Restoration. Divers. Distrib. 2011, 17, 1015–1029. [Google Scholar] [CrossRef]
- Bengtsson, J.; Nilsson, S.G.; Franc, A.; Menozzi, P. Biodiversity, Disturbances, Ecosystem Function and Management of European Forests. For. Ecol. Manag. 2000, 132, 39–50. [Google Scholar] [CrossRef]
- Bardsley, D.; Edwards-Jones, G. Stakeholders’ Perceptions of the Impacts of Invasive Exotic Plant Species in the Mediterranean Region. GeoJournal 2006, 65, 199–210. [Google Scholar] [CrossRef] [Green Version]
- Hussain, M.I.; El-Sheikh, M.A.; Reigosa, M.J. Allelopathic Potential of Aqueous Extract from Acacia Melanoxylon R. Br. on Lactuca sativa. Plants 2020, 9, 1228. [Google Scholar] [CrossRef]
- Souza-Alonso, P.; Rodríguez, J.; González, L.; Lorenzo, P. Here to Stay. Recent Advances and Perspectives about Acacia Invasion in Mediterranean Areas. Ann. For. Sci. 2017, 74, 55. [Google Scholar] [CrossRef] [Green Version]
- Raposo, M.A.M.; Gomes, C.J.P.; Nunes, L.J.R. Selective Shrub Management to Preserve Mediterranean Forests and Reduce the Risk of Fire: The Case of Mainland Portugal. Fire 2020, 3, 65. [Google Scholar] [CrossRef]
- Mainali, K.P.; Warren, D.L.; Dhileepan, K.; McConnachie, A.; Strathie, L.; Hassan, G.; Karki, D.; Shrestha, B.B.; Parmesan, C. Projecting Future Expansion of Invasive Species: Comparing and Improving Methodologies for Species Distribution Modeling. Glob. Chang. Biol. 2015, 21, 4464–4480. [Google Scholar] [CrossRef] [PubMed]
- Martins, F.; Alegria, C.; Artur, G. Mapping Invasive Alien Acacia Dealbata Link Using ASTER Multispectral Imagery: A Case Study in Central-Eastern of Portugal. For. Syst. 2016, 25, 13. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, A.T.; Gonçalves, J.; Fernandes, R.F.; Alves, S.; Marcos, B.; Lucas, R.; Teodoro, A.C.; Honrado, J.P. Estimating Invasion Success by Non-Native Trees in a National Park Combining World View-2 Very High Resolution Satellite Data and Species Distribution Models. Diversity 2017, 9, 6. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo, P.; González, L.; Reigosa, M.J. The Genus Acacia as Invader: The Characteristic Case of Acacia Dealbata Link in Europe. Ann. For. Sci. 2010, 67, 101. [Google Scholar] [CrossRef] [Green Version]
- Marchante, H.; Morais, M.; Freitas, H.; Marchante, E. Guia Prático Para a Identificação de Plantas Invasoras Em Portugal; Imprensa da Universidade de Coimbra: Coimbra, Portugal, 2014; ISBN 978-989-26-0786-3. [Google Scholar]
- Nunes, L.J.R.; Raposo, M.A.M.; Meireles, C.I.R.; Pinto Gomes, C.J.; Ribeiro, N.M.C.A. Fire as a Selection Agent for the Dissemination of Invasive Species: Case Study on the Evolution of Forest Coverage. Environments 2020, 7, 57. [Google Scholar] [CrossRef]
- Malato-Beliz, J. A Serra de Monchique—Flora e Vegetação. Coleção de Parques Naturais No 10; Serviço Nacional de Parques, Reservas e Património Paisagístico: Lisboa, Portugal, 1982. [Google Scholar]
- Pinto-Gomes, C.; Meireles, C.; Raposo, M.; Castro, C.; Matos, R.; Santos, P. Guia da Excursão Geobotânica do XIII Seminário Internacional de Gestão e Conservação da Biodiversidade-Vale do Lobo; Loulé: Algarve, Portugal, 2019. [Google Scholar]
- Breton, C.; Guerin, J.; Ducatillion, C.; Médail, F.; Kull, C.A.; Bervillé, A. Taming the Wild and ‘Wilding’ the Tame: Tree Breeding and Dispersal in Australia and the Mediterranean. Plant Sci. 2008, 175, 197–205. [Google Scholar] [CrossRef]
- Paiva, J. Acacia Mill. In Flora Iberica—Plantas Vasculares de la Península Iberica e Islas Baleares; Talavera, S., Aedo, C., Castroviejo, S., Romero, C., Sáez, L., Salgueiro, F.J., Velayos, M., Eds.; Real Jardín Botánico, CSIC: Madrid, Spain, 1999; Volume VII, pp. 11–25. [Google Scholar]
- Liberal, M.; Esteves, M. Invasão de Acacia Dealbata Link No Parque Nacional Da Peneda-Gerês; SPCF/ADERE: Gerês, Portugal, 1999. [Google Scholar]
- Gallagher, R.V.; Leishman, M.R.; Miller, J.T.; Hui, C.; Richardson, D.M.; Suda, J.; Trávníček, P. Invasiveness in Introduced Australian Acacias: The Role of Species Traits and Genome Size. Divers. Distrib. 2011, 17, 884–897. [Google Scholar] [CrossRef]
- Rodríguez-Echeverría, S.; Fajardo, S.; Ruiz-Díez, B.; Fernández-Pascual, M. Differential Effectiveness of Novel and Old Legume–Rhizobia Mutualisms: Implications for Invasion by Exotic Legumes. Oecologia 2012, 170, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Milton, S.J.; Hall, A.V. Reproductive Biology of Australian Acacias in the South-Western Cape Province, South Africa. Trans. R. Soc. S. Afr. 1981, 44, 465–487. [Google Scholar] [CrossRef]
- Gibson, M.R.; Richardson, D.M.; Marchante, E.; Marchante, H.; Rodger, J.G.; Stone, G.N.; Byrne, M.; Fuentes-Ramírez, A.; George, N.; Harris, C.; et al. Reproductive Biology of Australian Acacias: Important Mediator of Invasiveness? Divers. Distrib. 2011, 17, 911–933. [Google Scholar] [CrossRef]
- Correia, M.; Castro, S.; Ferrero, V.; Crisóstomo, J.A.; Rodríguez-Echeverría, S. Reproductive Biology and Success of Invasive Australian Acacias in Portugal. Bot. J. Linn. Soc. 2014, 174, 574–588. [Google Scholar] [CrossRef] [Green Version]
- Passos, I.; Marchante, H.; Pinho, R.; Marchante, E. What We Don’t Seed: The Role of Long-Lived Seed Banks as Hidden Legacies of Invasive Plants. Plant Ecol. 2017, 218, 1313–1324. [Google Scholar] [CrossRef]
- Wilson, J.R.U.; Gairifo, C.; Gibson, M.R.; Arianoutsou, M.; Bakar, B.B.; Baret, S.; Celesti-Grapow, L.; DiTomaso, J.M.; Dufour-Dror, J.-M.; Kueffer, C.; et al. Risk Assessment, Eradication, and Biological Control: Global Efforts to Limit Australian Acacia Invasions. Divers. Distrib. 2011, 17, 1030–1046. [Google Scholar] [CrossRef] [Green Version]
- Silva, J.S.; Vaz, P.; Moreira, F.; Catry, F.; Rego, F.C. Wildfires as a Major Driver of Landscape Dynamics in Three Fire-Prone Areas of Portugal. Landsc. Urban Plan. 2011, 101, 349–358. [Google Scholar] [CrossRef] [Green Version]
- Catford, J.A.; Daehler, C.C.; Murphy, H.T.; Sheppard, A.W.; Hardesty, B.D.; Westcott, D.A.; Rejmánek, M.; Bellingham, P.J.; Pergl, J.; Horvitz, C.C.; et al. The Intermediate Disturbance Hypothesis and Plant Invasions: Implications for Species Richness and Management. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 231–241. [Google Scholar] [CrossRef]
- Vieites-Blanco, C.; González-Prieto, S.J. Invasiveness, Ecological Impacts and Control of Acacias in Southwestern Europe—A Review. Web Ecol. 2020, 20, 33–51. [Google Scholar] [CrossRef]
- Rivas-Martínez, S.; Penas, Á.; Díaz González, T.E.; Cantó, P.; del Río, S.; Costa, J.C.; Herrero, L.; Molero, J. Biogeographic Units of the Iberian Peninsula and Baelaric Islands to District Level. A Concise Synopsis. In The Vegetation of the Iberian Peninsula: Volume 1; Plant and Vegetation; Loidi, J., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 131–188. ISBN 978-3-319-54784-8. [Google Scholar]
- Rivas-Martínez, S.; Penas, Á.; del Río, S.; Díaz González, T.E.; Rivas-Sáenz, S. Bioclimatology of the Iberian Peninsula and the Balearic Islands. In The Vegetation of the Iberian Peninsula: Volume 1; Plant and Vegetation; Loidi, J., Ed.; Springer International Publishing: Cham, switzerland, 2017; pp. 29–80. ISBN 978-3-319-54784-8. [Google Scholar]
- Ferreira, N.; Vieira, G. Guia Geológico e Geomorfológico: Do Parque Natural da Serra da Estrela; Locais de Interesse Geológico e Geomorfológico; ICNF—Instituto da Conservação da Naturaleza: Lisboa, Portugal, 1999. [Google Scholar]
- Marques, J.E.; Duarte, J.M.; Constantino, A.T.; Martins, A.A.; Aguiar, C.; Rocha, F.T.; Inácio, M.; Marques, J.M.; Chaminé, H.I.; Teixeira, J.; et al. Vadose Zone Characterisation of a Hydrogeologic System in a Mountain Region: Serra Da Estrela Case Study (Central Portugal). In Aquifer Systems Management: Darcy’s Legacy In a World Impending Water Shortage; Chery, L., de Marsilly, G., Eds.; Taylor & Francis: London, UK, 2007; pp. 207–221. ISBN 978-0-415-44355-5. [Google Scholar]
- Beltran, R.S. Distribution and Autoecology off Prunus lusitanica L. in Iberian Peninsula. For. Syst. 2006, 15, 187–198. [Google Scholar] [CrossRef] [Green Version]
- Costa, J.C.; Neto, C.; Aguiar, C.; Capelo, J.; Espírito Santo, M.D.; Honrado, J.J.; Gomes, C.P.; Monteiro-Henriques, T.; Sequeira, M.; Lousã, M. Vascular plant communities in Portugal (continental, the Azores and Madeira). Glob. Geobot. 2012, 2, 1–180. [Google Scholar]
- Santos, M.; Freitas, R.; Crespí, A.L.; Hughes, S.J.; Cabral, J.A. Predicting Trends of Invasive Plants Richness Using Local Socio-Economic Data: An Application in North Portugal. Environ. Res. 2011, 111, 960–966. [Google Scholar] [CrossRef] [PubMed]
- Marchante, H.; Marchante, E.; Freitas, H. Invasion of the Portuguese Dune Ecosystems by the Exotic Species Acacia longifolia (Andrews) Willd.: Effects at the Community Level. In Plant Invasions: Ecological Threats and Managagement Solutions; Child, L.E., Brock, J.H., Brundu, G., Prach, K., Pysek, P., Wade, P.M., Williamson, M., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2003; pp. 75–85. [Google Scholar]
- Marchante, H.; Freitas, H.; Hoffmann, J.H. Seed Ecology of an Invasive Alien Species, Acacia longifolia (Fabaceae), in Portuguese Dune Ecosystems. Am. J. Bot. 2010, 97, 1780–1790. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.; Carvalho, J.C.; Gomes, P. Invasive Plants Induce the Taxonomic and Functional Replacement of Dune Spiders. Biol. Invasions 2018, 20, 533–545. [Google Scholar] [CrossRef]
- Hernández, L.; Martínez-Fernández, J.; Cañellas, I.; de la Cueva, A.V. Assessing Spatio-Temporal Rates, Patterns and Determinants of Biological Invasions in Forest Ecosystems. The Case of Acacia Species in NW Spain. For. Ecol. Manag. 2014, 329, 206–213. [Google Scholar] [CrossRef] [Green Version]
- Higgins, S.I.; Richardson, D.M.; Cowling, R.M. Validation of a Spatial Simulation Model of a Spreading Alien Plant Population. J. Appl. Ecol. 2001, 38, 571–584. [Google Scholar] [CrossRef]
- Correia, T.P.; Abreu, A.C.; Oliveira, R. Identificação de Unidades de Paisagem: Metodologia aplicada a Portugal Continental. FIT 2001, 36. [Google Scholar] [CrossRef] [Green Version]
- Raposo, M. Séries de Vegetação Prioritárias Para a Conservação No Centro e Sul de Portugal Continental. Botanique 2016, 1, 113–148. [Google Scholar]
- Pheloung, P.C.; Williams, P.A.; Halloy, S.R. A Weed Risk Assessment Model for Use as a Biosecurity Tool Evaluating Plant Introductions. J. Environ. Manag. 1999, 57, 239–251. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, M.R.; Aguiar, F.C.; Martins, M.J.; Rico, N.; Ferreira, M.T.; Correia, A.C. Carbon Stock Estimations in a Mediterranean Riparian Forest: A Case Study Combining Field Data and UAV Imagery. Forests 2020, 11, 376. [Google Scholar] [CrossRef] [Green Version]
- Guerrero, F.; Hernández, C.; Toledo, M.; Espinoza, L.; Carrasco, Y.; Arriagada, A.; Muñoz, A.; Taborga, L.; Bergmann, J.; Carmona, C. Leaf Thermal and Chemical Properties as Natural Drivers of Plant Flammability of Native and Exotic Tree Species of the Valparaíso Region, Chile. Int. J. Environ. Res. Public Health 2021, 18, 7191. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Sample | Di (m) | Df (m) | Increase (m) | LI14 (m) | ALI (m) |
|---|---|---|---|---|---|
| 1 | 40.50 | 65.38 | 24.88 | 12.44 | 1.04 |
| 2 | 51.40 | 81.32 | 29.92 | 14.96 | 1.25 |
| 3 | 23.60 | 36.10 | 12.50 | 6.25 | 0.52 |
| 4 | 32.20 | 43.89 | 11.69 | 5.85 | 0.49 |
| 5 | 18.10 | 34.60 | 16.50 | 8.25 | 0.69 |
| 6 | 56.27 | 72.16 | 15.89 | 7.95 | 0.66 |
| 7 | 51.24 | 96.67 | 45.43 | 22.72 | 1.89 |
| 8 | 59.43 | 84.67 | 25.24 | 12.62 | 1.05 |
| 9 | 0.00 | 9.61 | 9.61 | 4.81 | 0.40 |
| 10 | 11.14 | 17.37 | 6.23 | 3.12 | 0.26 |
| 11 | 46.13 | 69.39 | 23.26 | 11.63 | 0.97 |
| 12 | 41.63 | 58.78 | 17.15 | 8.58 | 0.71 |
| 13 | 38.54 | 57.69 | 19.15 | 9.58 | 0.80 |
| 14 | 30.11 | 51.39 | 21.28 | 10.64 | 0.89 |
| 15 | 0.00 | 17.03 | 17.03 | 8.52 | 0.71 |
| Average | 9.86 | 0.82 |
| Sample | Ai (m²) | Af (m²) | Increase (m²) | IR14 (%) | AIR (%) |
|---|---|---|---|---|---|
| 1 | 1287.60 | 5556.45 | 1691.13 | 131% | 11% |
| 2 | 2073.94 | 5782.58 | 2763.43 | 133% | 11% |
| 3 | 437.21 | 1961.50 | 508.00 | 116% | 10% |
| 4 | 813.92 | 2331.77 | 442.08 | 54% | 5% |
| 5 | 703.26 | 1545.50 | 842.24 | 120% | 10% |
| 6 | 3006 | 5087.23 | 2081.68 | 69% | 6% |
| 7 | 7206 | 16,406.74 | 9200.32 | 128% | 11% |
| 8 | 648 | 1189.59 | 541.44 | 84% | 7% |
| 9 | 0.00 | 141.94 | 141.94 | - | - |
| 10 | 114 | 280.94 | 166.56 | 146% | 12% |
| 11 | 2160 | 3702.04 | 1541.86 | 71% | 6% |
| 12 | 2254 | 3738.46 | 1484.60 | 66% | 5% |
| 13 | 1808 | 2884.51 | 1076.41 | 60% | 5% |
| 14 | 1926 | 2673.62 | 747.95 | 39% | 3% |
| 15 | 0.00 | 400.78 | 400.78 | - | - |
| Average | 94% | 8% |
| Sample | Df (m) | IL20 (m) | D20 (m) | A 2019 (m²) | A 2039 (m²) |
|---|---|---|---|---|---|
| 1 | 65.38 | 20.73 | 106.85 | 5556.45 | 44,797.83 |
| 2 | 81.32 | 24.93 | 131.19 | 5782.58 | 46,620.96 |
| 3 | 36.10 | 10.42 | 56.93 | 1961.50 | 15,814.22 |
| 4 | 43.89 | 9.74 | 63.37 | 2331.77 | 18,799.46 |
| 5 | 34.60 | 13.75 | 62.10 | 1545.50 | 12,460.30 |
| 6 | 72.16 | 13.24 | 98.64 | 5087.23 | 41,014.83 |
| 7 | 96.67 | 37.86 | 172.39 | 16,406.74 | 132,276.25 |
| 8 | 84.67 | 21.03 | 126.74 | 1189.59 | 9590.85 |
| 9 | 9.61 | 8.01 | 25.63 | 141.94 | 1144.36 |
| 10 | 17.37 | 5.19 | 27.75 | 280.94 | 2265.03 |
| 11 | 69.39 | 19.38 | 108.16 | 3702.04 | 29,847.00 |
| 12 | 58.78 | 14.29 | 87.36 | 3738.46 | 30,140.63 |
| 13 | 57.69 | 15.96 | 89.61 | 2884.51 | 23,255.82 |
| 14 | 51.39 | 17.73 | 86.86 | 2673.62 | 21,555.56 |
| 15 | 17.03 | 14.19 | 45.41 | 400.78 | 3231.21 |
| Average | 16.43 | Total areas | 53,683.65 | 432,814.31 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raposo, M.A.M.; Pinto Gomes, C.J.; Nunes, L.J.R. Evaluation of Species Invasiveness: A Case Study with Acacia dealbata Link. on the Slopes of Cabeça (Seia-Portugal). Sustainability 2021, 13, 11233. https://doi.org/10.3390/su132011233
Raposo MAM, Pinto Gomes CJ, Nunes LJR. Evaluation of Species Invasiveness: A Case Study with Acacia dealbata Link. on the Slopes of Cabeça (Seia-Portugal). Sustainability. 2021; 13(20):11233. https://doi.org/10.3390/su132011233
Chicago/Turabian StyleRaposo, Mauro A. M., Carlos J. Pinto Gomes, and Leonel J. R. Nunes. 2021. "Evaluation of Species Invasiveness: A Case Study with Acacia dealbata Link. on the Slopes of Cabeça (Seia-Portugal)" Sustainability 13, no. 20: 11233. https://doi.org/10.3390/su132011233
APA StyleRaposo, M. A. M., Pinto Gomes, C. J., & Nunes, L. J. R. (2021). Evaluation of Species Invasiveness: A Case Study with Acacia dealbata Link. on the Slopes of Cabeça (Seia-Portugal). Sustainability, 13(20), 11233. https://doi.org/10.3390/su132011233