A System Dynamics Model for Assessing the Efficacy of Lethal Control for Sustainable Management of Ochotona curzoniae on Tibetan Plateau




Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Natural Population Demography
2.3. Lethal Control Population Demography
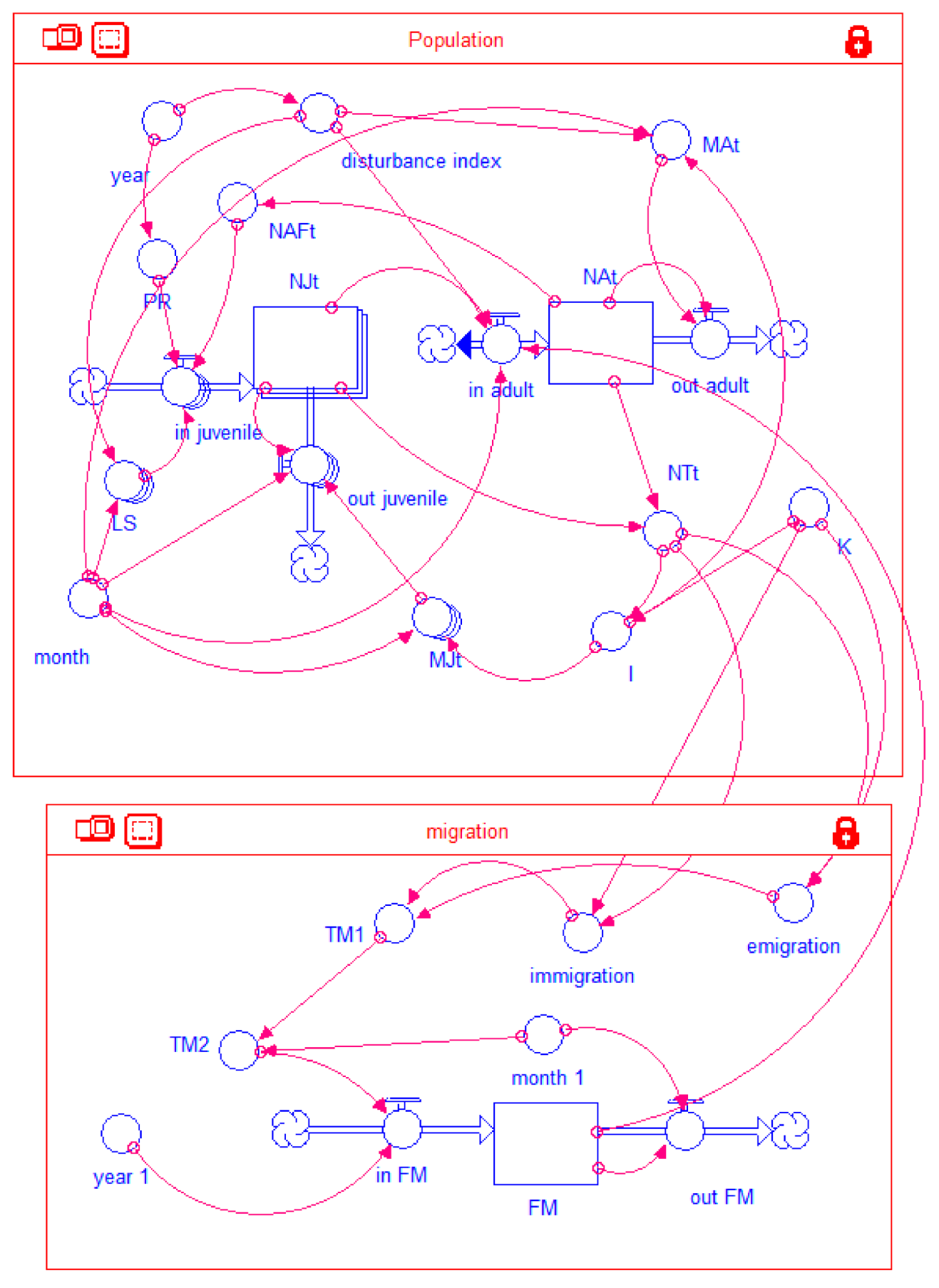
2.4. Model Structure and Parameterization
3. Results
3.1. Plateau Pika Population Trend under Natural Conditions
3.2. Model of Plateau Pika Population
4. Discussion
4.1. Model Evaluation
4.2. Pikas Managements
4.3. Management Implication
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singleton, G.R. Rodent Outbreaks: Ecology and Impacts; International Rice Researh Institute: Los Banos, Philippines, 2010. [Google Scholar]
- Redlich, S.; Martin, E.A.; Steffan-Dewenter, I. Landscape-level crop diversity benefits biological pest control. J. Appl. Ecol. 2018, 55, 2419–2428. [Google Scholar] [CrossRef]
- Fan, N.; Zhou, W.; Wei, W.; Wang, Q.; Jiang, Y. Rodent pest management in the Qinghai-Tibet alpine meadow ecosystem. In Ecologically-Based Rodent Management; Singleton, G., Hinds, L., Leirs, H., Zhang, Z.B., Eds.; Australian Centre for International Agricultural Research: Canberra, Australia, 1999; pp. 285–304. [Google Scholar]
- Russell, J.C.; Jones, H.P.; Armstrong, D.P.; Courchamp, F.; Kappes, P.J.; Seddon, P.J.; Oppel, S.; Rauzon, M.J.; Cowan, P.E.; Rocamora, G.; et al. Importance of lethal control of invasive predators for island conservation. Conserv. Biol. 2016, 30, 670–672. [Google Scholar] [CrossRef] [PubMed]
- Krebs, C.J. Population cycles revisited. J. Mammal. 1996, 77, 8–24. [Google Scholar] [CrossRef]
- Ericsson, R.J. Male antifertility compounds: U-5897 as a rat chemosterilant. J. Reprod. Fertil. 1970, 22, 213–222. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Stenseth, N.C.; Leirs, H.; Mercelis, S.; Mwanjabe, P. Comparing strategies for controlling an African pest rodent: An empirically based theoretical study. J. Appl. Ecol. 2001, 38, 1020–1031. [Google Scholar] [CrossRef]
- Shi, D.Z.; Wan, X.R.; Davis, S.A.; Pech, R.P.; Zhang, Z.B. Simulation of lethal control and fertility control in a demographic model for Brandt’s vole Microtus brandti. J. Appl. Ecol. 2002, 39, 337–348. [Google Scholar] [CrossRef]
- Shang, Z.H.; Long, R.J. Formation causes and recovery of the “Black Soil Type” degraded alpine grassland in Qinghai-Tibetan Plateau. Front. Agric. China 2007, 1, 197–202. [Google Scholar] [CrossRef]
- Harris, R.B. Rangeland degradation on the Qinghai-Tibetan plateau: A review of the evidence of its magnitude and causes. J. Arid Environ. 2010, 74, 1–12. [Google Scholar] [CrossRef]
- Xin, H. A green fervor sweeps the Qinghai-Tibetan Plateau. Science 2008, 321, 633–635. [Google Scholar] [CrossRef]
- Wu, G.; Du, G. Discussion on ecological construction and sustainable development of degraded alpine grassland ecosystem of the qinghai-tibetan plateau. Chin. J. Nat. 2007, 29, 159–164. [Google Scholar]
- Smith, A.T.; Xie, Y. A Guide to the Mammals of China; Princeton University Press: Princeton, NJ, USA, 2008. [Google Scholar]
- Koju, N.P.; He, K.; Chalise, M.K.; Ray, C.; Chen, Z.; Zhang, B.; Wan, T.; Chen, S.; Jiang, X. Multilocus approaches reveal underestimated species diversity and inter-specific gene flow in pikas (Ochotona) from southwestern China. Mol. Phylogenet. Evol. 2017, 107, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Yang, M.; Li, W.; Li, K.; Zhang, Y.; Smith, A.T. Seasonal variation of family group structure of plateau pikas (Ochotona curzoniae). Acta Theriol. Sin. 2008, 28, 144–150. [Google Scholar]
- Qu, J.; Li, W.; Yang, M.; Ji, W.; Zhang, Y. Life history of the plateau pika (Ochotona curzoniae) in alpine meadows of the Tibetan Plateau. Mamm. Biol. 2013, 78, 68–72. [Google Scholar] [CrossRef]
- Lai, C.H.; Smith, A.T. Keystone status of plateau pikas (Ochotona curzoniae): Effect of control on biodiversity of native birds. Biodivers. Conserv. 2003, 12, 1901–1912. [Google Scholar] [CrossRef]
- Harris, R.B.; Jiake, Z.; Yinqiu, J.; Kai, Z.; Chunyan, Y.; Yu, D.W. Evidence that the Tibetan fox is an obligate predator of the plateau pika: Conservation implications. J. Mamm. 2014, 95, 1207–1221. [Google Scholar] [CrossRef]
- Yu, C.; Zhang, J.; Pang, X.P.; Wang, Q.; Zhou, Y.P.; Guo, Z.G. Soil disturbance and disturbance intensity: Response of soil nutrient concentrations of alpine meadow to plateau pika bioturbation in the Qinghai-Tibetan Plateau, China. Geoderma 2017, 307, 98–106. [Google Scholar] [CrossRef]
- Wang, Q.; Yu, C.; Pang, X.P.; Jin, S.H.; Zhang, J.; Guo, Z.G. The disturbance and disturbance intensity of small and semi-fossorial herbivores alter the belowground bud density of graminoids in alpine meadows. Ecol. Eng. 2018, 113, 35–42. [Google Scholar] [CrossRef]
- Qin, Y.; Chen, J.; Yi, S. Plateau pikas burrowing activity accelerates ecosystem carbon emission from alpine grassland on the Qinghai-Tibetan Plateau. Ecol. Eng. 2015, 84, 287–291. [Google Scholar] [CrossRef]
- Yu, C.; Pang, X.P.; Wang, Q.; Jin, S.H.; Shu, C.C.; Guo, Z.G. Soil nutrient changes induced by the presence and intensity of plateau pika (Ochotona curzoniae) disturbances in the Qinghai-Tibet Plateau, China. Ecol. Eng. 2017, 106, 1–9. [Google Scholar] [CrossRef]
- Liu, Y.; Fan, J.; Shi, Z.; Yang, X.; Harris, W. Relationships between plateau pika (Ochotona curzoniae) densities and biomass and biodiversity indices of alpine meadow steppe on the Qinghai–Tibet Plateau China. Ecol. Eng. 2017, 102, 509–518. [Google Scholar] [CrossRef]
- Wu, L.; Wang, H. Poisoning the pika: Must protection of grasslands be at the expense of biodiversity? Sci. China Life Sci. 2017, 60, 545. [Google Scholar] [CrossRef] [PubMed]
- Pech, R.P.; Jiebu; Anthony, A.D.; Zhang, Y.M.; Lin, H. Population dynamics and responses to management of plateau pikas (Ochotona curzoniae). J. Appl. Ecol. 2007, 44, 615–624. [Google Scholar] [CrossRef]
- Köhler, H.R.; Triebskorn, R. Wildlife ecotoxicology of pesticides: Can we track effects to the population level and beyond? Science 2013, 341, 759–765. [Google Scholar] [CrossRef]
- Qu, J.; Russell, J.C.; Ji, W.; Yang, M.; Chen, Q.; Li, W.; Zhang, Y. Five-year population dynamics of plateau pikas (Ochotona curzoniae) on the east of Tibetan Plateau. Eur. J. Wildlife Res. 2017, 63, 51. [Google Scholar] [CrossRef]
- Qu, J.; Liu, M.; Yang, M.; Zhang, Y.; Ji, W. Reproduction of plateau pika (Ochotona curzoniae) on the Qinghai-Tibetan plateau. Eur. J. Wildlife Res. 2012, 58, 269–277. [Google Scholar] [CrossRef]
- Qu, J.; Li, W.; Yang, M.; Li, K.; Zhang, Y. Methods for large scale assessment of small mammal abundance in open habitats: Plateau pika (Ochotona curzoniae) in alpine grassland. Pol. J. Ecol. 2011, 59, 829–833. [Google Scholar]
- Qu, J. Life History Characteristics and Response to Fertility Control of Plateau Pika Ochotona curzoniae Populations. Ph.D. Thesis, University of Chinese Academy of Sciences, Beijing, China, 2011. [Google Scholar]
- Zhang, Y.M.; Fan, N.C.; Wang, Q.Y.; Jing, Z.C. The changing ecological process of rodent communities during rodent pest managements in alpine meadow. Acta Theriol. Sin. 1998, 18, 137–143. [Google Scholar]
- DeMaso, S.J.; Grant, W.E.; Hernández, F.; Brennan, L.A.; Silvy, N.J.; Wu, X.B.; Bryant, F.C. A population model to simulate northern bobwhite population dynamics in southern Texas. J. Wildl. Manag. 2011, 75, 319–332. [Google Scholar] [CrossRef]
- Weclaw, P.; Hudson, R.J. Simulation of conservation and management of woodland caribou. Ecol. Model. 2004, 177, 75–94. [Google Scholar] [CrossRef]
- Weller, F.; Sherley, R.B.; Waller, L.J.; Ludynia, K.; Geldenhuys, D.; Shannon, L.J.; Jarre, A. System dynamics modelling of the Endangered African penguin populations on Dyer and Robben islands, South Africa. Ecol. Model. 2016, 327, 44–56. [Google Scholar] [CrossRef]
- Dobson, F.S.; Smith, A.T.; Wang, X.G. Social and ecological influences on dispersal and philopatry in the plateau pika (Ochotona curzoniae). Behav. Ecol. 1998, 9, 622–635. [Google Scholar] [CrossRef]
- Wang, X.; Dai, K. Studies on the population reproduction ecology of plateau pika. Zool. Res. 1991, 12, 155–161. [Google Scholar]
- Frederiksen, M.; Bregnballe, T. Evidence for density-dependent survival in adult cormorants from a combined analysis of recoveries and resightings. J. Anim. Ecol. 2000, 69, 737–752. [Google Scholar] [CrossRef]
- Norman, D.; Peach, W.J. Density-dependent survival and recruitment in a long-distance Palaearctic migrant, the Sand Martin Riparia riparia. Ibis 2013, 155, 284–296. [Google Scholar] [CrossRef]
- Bunnefeld, N.; Börger, L.; van Moorter, B.; Rolandsen, C.M.; Dettki, H.; Solberg, E.J.; Ericsson, G. A model-driven approach to quantify migration patterns: Individual, regional and yearly differences. J. Anim. Ecol. 2011, 80, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Arditi, R.; Lobry, C.; Sari, T. Is dispersal always beneficial to carrying capacity? New insights from the multi-patch logistic equation. Theor. Popul. Biol. 2015, 106, 45–59. [Google Scholar] [CrossRef]
- McLaren, I.A. Natural Regulation of Animal Populations; Routledge: New York, NY, USA, 1971. [Google Scholar]
- Fordham, D.; Georges, A.; Brook, B. Experimental evidence for density-dependent responses to mortality of snake-necked turtles. Oecologia 2009, 159, 271–281. [Google Scholar] [CrossRef]
- Taylor Caz, M.; Norris, D.R. Predicting conditions for migration: Effects of density dependence and habitat quality. Biol. Lett. 2007, 3, 280–284. [Google Scholar] [CrossRef]
- Fay, R.; Weimerskirch, H.; Delord, K.; Barbraud, C. Population density and climate shape early-life survival and recruitment in a long-lived pelagic seabird. J. Anim. Ecol. 2015, 84, 1423–1433. [Google Scholar] [CrossRef]
- Qu, J.; Chen, Q.; Zhang, Y. Behaviour and reproductive fitness of postdispersal in plateau pikas (Ochotona curzoniae) on the Tibetan Plateau. Mamm. Res. 2018, 63, 151–159. [Google Scholar] [CrossRef]
- Dickman, C.R.; Greenville, A.C.; Beh, C.L.; Tamayo, B.; Wardle, G.M. Social organization and movements of desert rodents during population “booms” and “busts” in central Australia. J. Mamm. 2010, 91, 798–810. [Google Scholar] [CrossRef]
- Zipkin, E.F.; Sullivan, P.J.; Cooch, E.G.; Kraft, C.E.; Shuter, B.J.; Weidel, B.C. Overcompensatory response of a smallmouth bass (Micropterus dolomieu) population to harvest: Release from competition? Can. J. Fish. Aquat. Sci. 2008, 65, 2279–2292. [Google Scholar] [CrossRef]
- Zhang, Z.B. Mathematical models of wildlife management by contraception. Ecol. Model. 2000, 132, 105–113. [Google Scholar] [CrossRef]
- Zipkin, E.F.; Kraft, C.E.; Cooch, E.G.; Sullivan, P.J. When can efforts to control nuisance and invasive species backfire? Ecol. Appl. 2009, 19, 1585–1595. [Google Scholar] [CrossRef] [PubMed]
- Byrom, A.E.; Innes, J.; Binny, R.N. A review of biodiversity outcomes from possum-focused pest control in New Zealand. Wildl. Res. 2016, 43, 228–253. [Google Scholar] [CrossRef]
- Pearson, D.E.; Callaway, R.M. Indirect effects of host-specific biological control agents. Trends Ecol. Evol. 2003, 18, 456–461. [Google Scholar] [CrossRef]
- Bald, J.; Sinquin, A.; Borja, A.; Caill-Milly, N.; Duclercq, B.; Dang, C.; de Montaudouin, X. A system dynamics model for the management of the Manila clam, Ruditapes philippinarum (Adams and Reeve, 1850) in the Bay of Arcachon (France). Ecol. Model. 2009, 220, 2828–2837. [Google Scholar] [CrossRef]
- Boiteau, G.; Vernon, R.S. Physical barriers for the control of insect pests. In Physical Control Methods in Plant Protection; Vincent, C., Panneton, B., Fleurat-Lessard, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2001; pp. 224–247. [Google Scholar]
- Otto, C.R.V.; Roth, C.L.; Carlson, B.L.; Smart, M.D. Land-use change reduces habitat suitability for supporting managed honey bee colonies in the Northern Great Plains. Proc. Natl. Acad. Sci. USA 2016, 113, 10430–10435. [Google Scholar] [CrossRef]
- Su, Q.Q.; Chen, Y.; Qin, J.; Wang, T.L.; Wang, D.H.; Liu, Q.S. Responses in reproductive organs, steroid hormones and CYP450 enzymes in female Mongolian gerbil (Meriones unguiculatus) over time after quinestrol treatment. Pestic. Biochem. Phys. 2017, 143, 122–126. [Google Scholar] [CrossRef]
- Chen, X.; Li, F.; Feng, L.; Yu, L.; Ding, J. An injectable thermogel containing levonorgestrel for long-acting contraception and fertility control of animals. J. Biomed. Nanotechnol. 2017, 13, 1357–1368. [Google Scholar] [CrossRef]
- Altieri, M.A.; Farrell, J.G.; Hecht, S.B.; Liebman, M.; Magdoff, F.; Murphy, B.; Norgaard, R.B.; Sikor, T.O. Integrated pest management. In Agroecology, 2nd ed.; Altieri, M.A., Ed.; CRC Press: Boca Raton, FL, USA, 2018; pp. 267–281. [Google Scholar]



{kind=link}
{kind=link}
| In juvenile = NAFt * PR * LS[L1] + NAFt + 1 * PR * LS[L2]. |
| Jt = in juvenilet − our juvenilet |
| NAFt = Nat * 0.51 |
| Out juvenile (L1) = if month = 8 then NJt[NL1] else NJt[NL1] * MJt[NL1] |
| Out juvenile (L2) = if month = 8 then NJt[NL2] else NJt[NL2] * MJt[NL2] |
| In adult = if month = 8 then (NJt[NL1] + NJt[NL2]) * 0.9 ELSE IF month = 5 OR month = 6 THEN 0 ELSE IF month = 7 THEN FM ELSE 0 |
| Out adult = Mat * NAt |
| MA (Aug. to Apr.) = 0.04 * I |
| I = if NTt < K then 1 else 3 |
| NTt = Nat + NJt [NL1] + NJt [NL2] |
| NAt = 33 NJt = 0 Immigration = if NTt < 88 and NTt ≥ 70 then NTt * 0.3 ELSE if NTt < 70 THEN NTt * 1 ELSE if NTt > K then NTt * 0.01 ELSE NTt * 0.05 |
| Emigration = if NTt < 88 then 0 ELSE if NTt > K THEN NTt * 0.1 ELSE NTt * 0.05 |
| TM1 = immigration-emigration |
| TM2# = if month_1 = 1 OR month_1 = 2 OR month_1 = 3 OR month_1 = 4 THEN TM1 ELSE 0 |
| Month 1 = time MOD 12 |
| Out FM = IF month_1 = 8 THEN FM ELSE 0 |
| Source of Variation | Estimate | SE | t Value | p Value |
|---|---|---|---|---|
| Intercept | 120.582 | 12.024 | 10.028 | <0.001 |
| Case 1 | −56.527 | 17.005 | −3.324 | 0.001 |
| Case 2 | −26.875 | 17.005 | −1.580 | 0.115 |
| Case 3 | −19.733 | 17.005 | −1.160 | 0.246 |
| Case 4 | −27.388 | 17.005 | −1.599 | 0.108 |
| Case 5 | −62.887 | 17.005 | −3.698 | <0.001 |
| Year | −0.687 | 1.801 | −0.381 | 0.703 |
| Year: Month | 0.210 | 0.069 | 3.062 | 0.002 |
| Case 1: Year | 0.310 | 2.467 | 0.126 | 0.900 |
| Case 2: Year | 1.626 | 2.467 | 0.659 | 0.510 |
| Case 3: Year | 2.010 | 2.467 | 0.815 | 0.416 |
| Case 4: Year | 4.175 | 2.467 | 1.692 | 0.091 |
| Case 5: Year | 0.307 | 2.467 | 0.124 | 0.901 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qu, J.; Liu, Z.; Guo, Z.; Li, Y.; Zhou, H. A System Dynamics Model for Assessing the Efficacy of Lethal Control for Sustainable Management of Ochotona curzoniae on Tibetan Plateau. Sustainability 2021, 13, 543. https://doi.org/10.3390/su13020543
Qu J, Liu Z, Guo Z, Li Y, Zhou H. A System Dynamics Model for Assessing the Efficacy of Lethal Control for Sustainable Management of Ochotona curzoniae on Tibetan Plateau. Sustainability. 2021; 13(2):543. https://doi.org/10.3390/su13020543
Chicago/Turabian StyleQu, Jiapeng, Zelin Liu, Zhenggang Guo, Yikang Li, and Huakun Zhou. 2021. "A System Dynamics Model for Assessing the Efficacy of Lethal Control for Sustainable Management of Ochotona curzoniae on Tibetan Plateau" Sustainability 13, no. 2: 543. https://doi.org/10.3390/su13020543
APA StyleQu, J., Liu, Z., Guo, Z., Li, Y., & Zhou, H. (2021). A System Dynamics Model for Assessing the Efficacy of Lethal Control for Sustainable Management of Ochotona curzoniae on Tibetan Plateau. Sustainability, 13(2), 543. https://doi.org/10.3390/su13020543