
Effects of Exogenous Application of Plant Growth Regulators (SNP and GA3) on Phytoextraction by Switchgrass (Panicum virgatum L.) Grown in Lead (Pb) Contaminated Soil


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil
2.2. Plant Growth Conditions
2.3. Selection of Exogenous NO Donor
2.4. Phytoextraction by Switchgrass Enhanced by Coordinated Application of SNP, Chelate and Soil Fungicide
2.5. Phytoextraction by Switchgrass Enhanced by Coordinated Application of SNP, GA3, Chelate and Soil Fungicide
2.6. Acid Digestion & Chemical Analysis of Plant Samples
2.7. Trypan Blue Staining of Roots for AMF Assessment
2.8. Statistical Analysis and Remediation Calculation
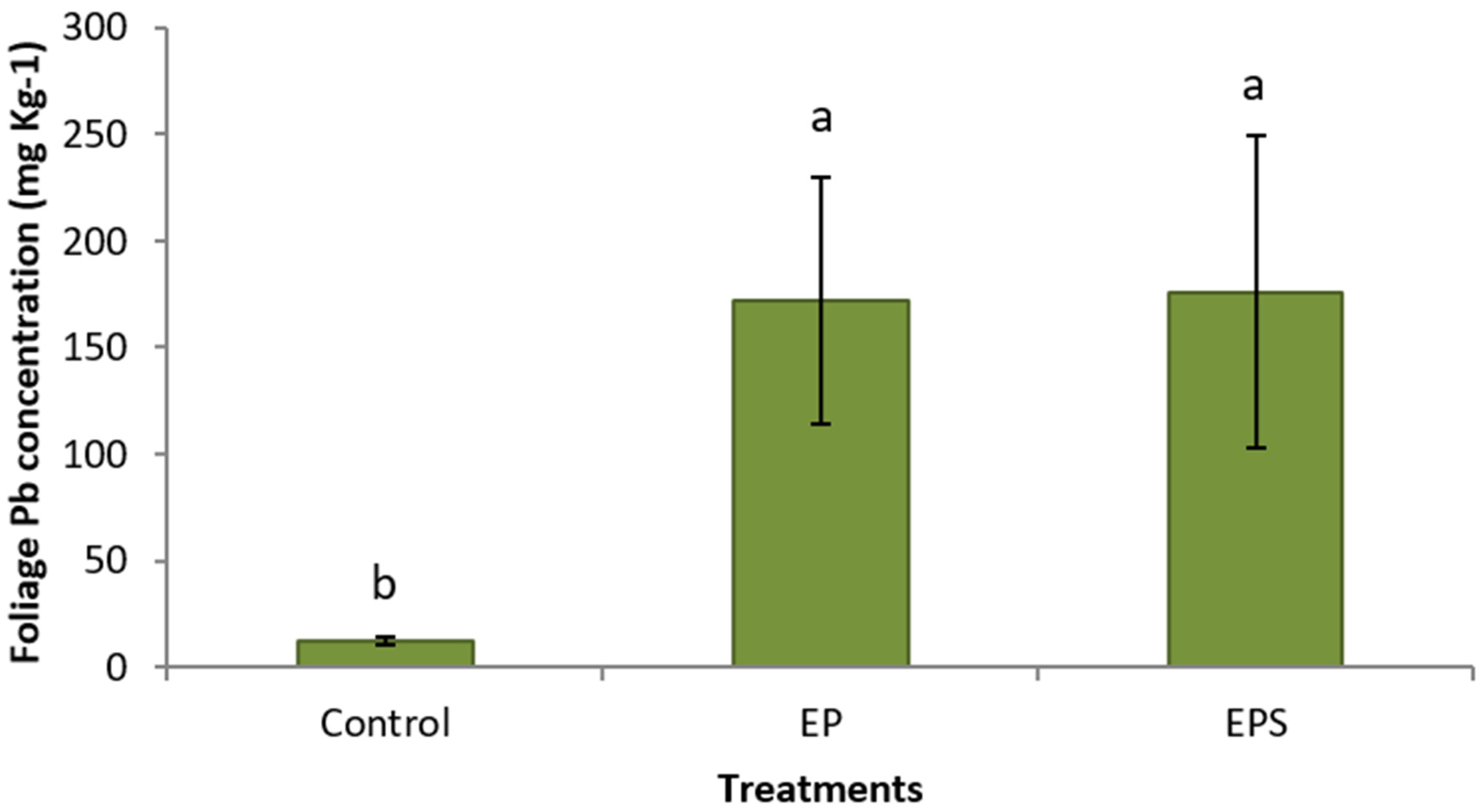
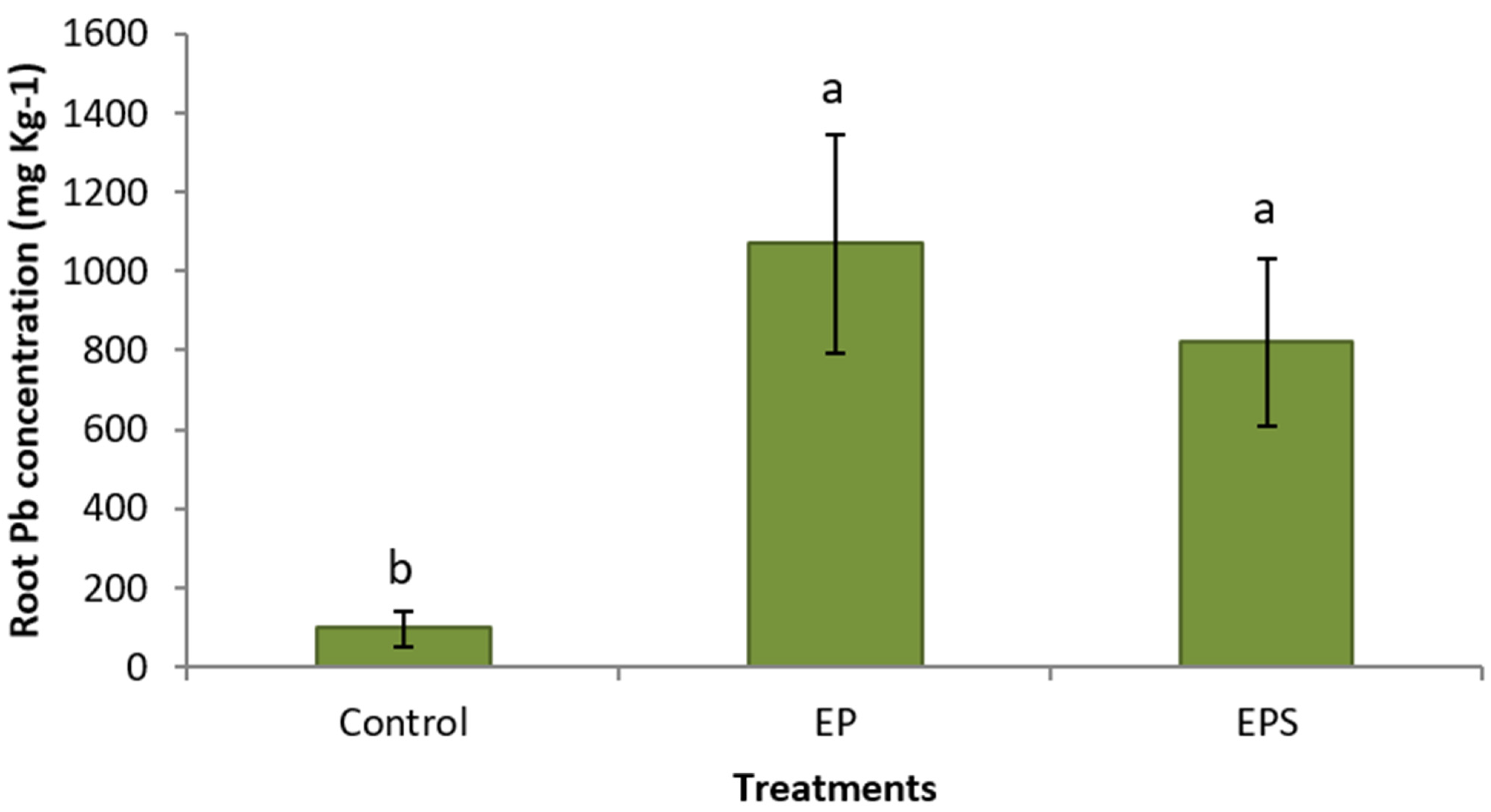
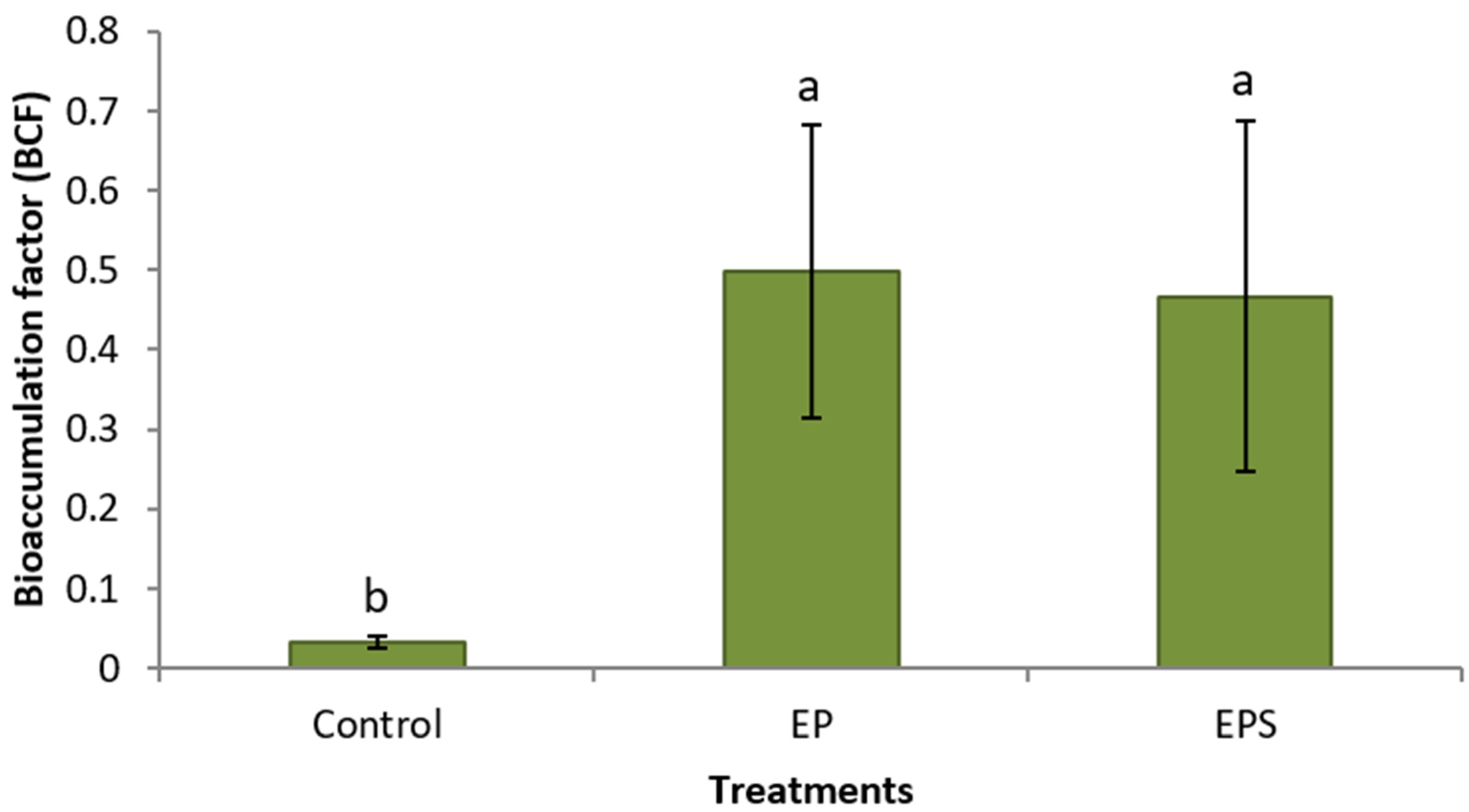
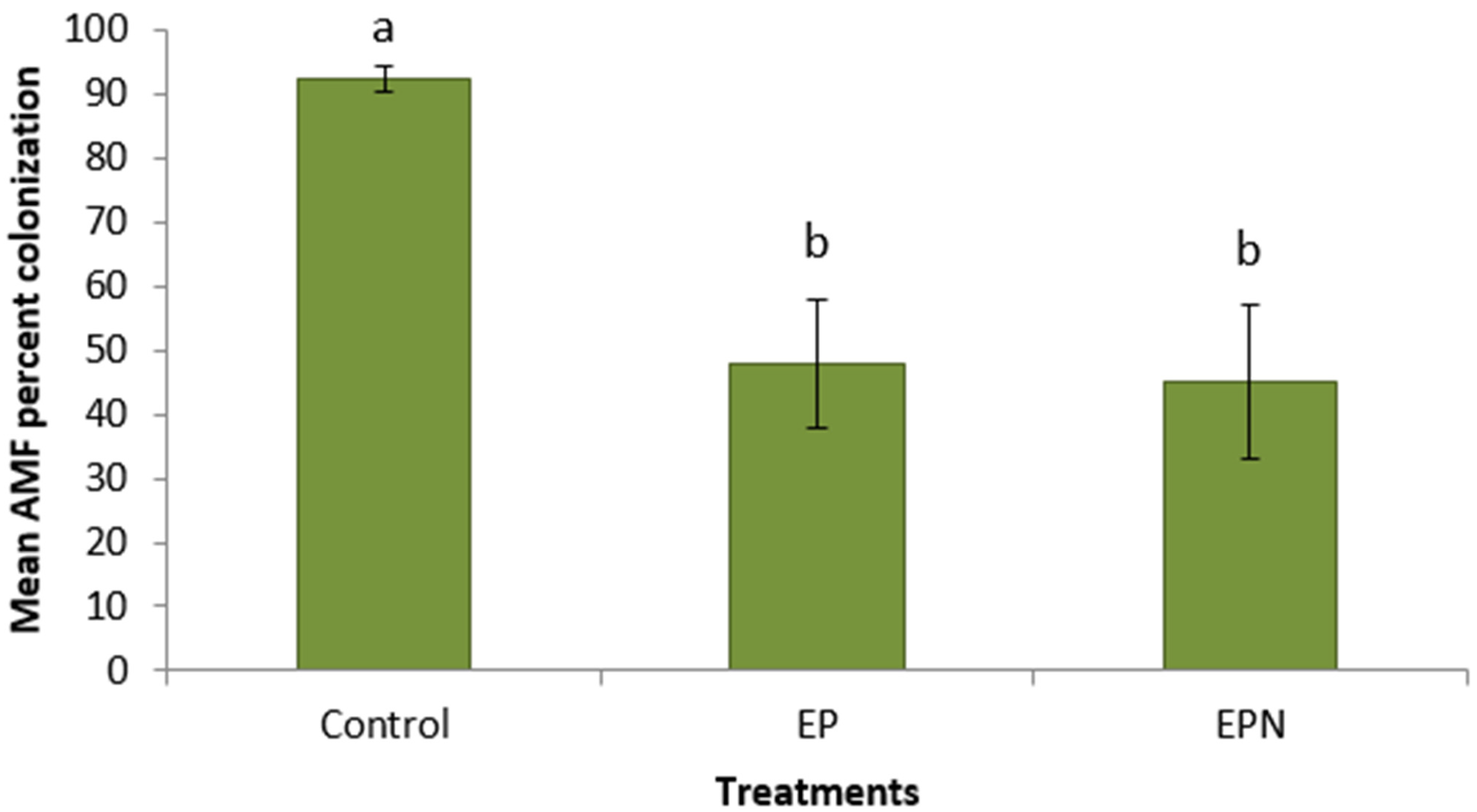
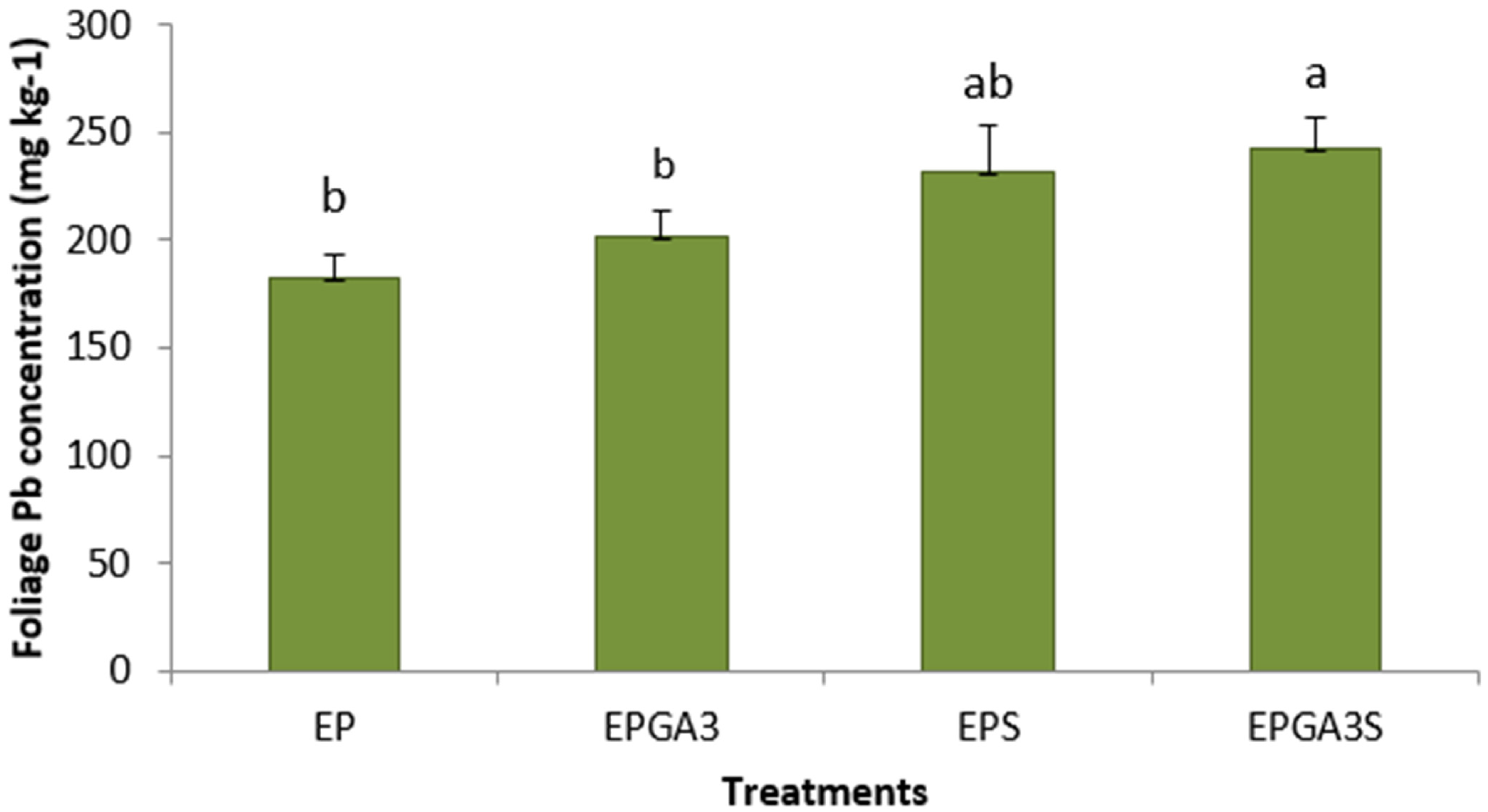
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lagerwerff, J.V.; Specht, A.W. Contamination of roadside soil and vegetation with cadmium, nickel, lead, and zinc. Environ. Sci. Technol. 1970, 4, 583–586. [Google Scholar] [CrossRef]
- Gulson, B.L.; Davis, J.J.; Mizon, K.J.; Korsch, M.J.; Bawden-Smith, J. Sources of lead in soil and dust and the use of dust fallout as a sampling medium. Sci. Total. Environ. 1995, 166, 245–262. [Google Scholar] [CrossRef]
- Mielke, H.W.; Reagan, P.L. Soil is an important pathway of human lead exposure. Environ. Health Perspect. 1998, 106 (Suppl. S1), 217. [Google Scholar] [PubMed] [Green Version]
- Johnson, A.W.; Gutierrez, M.; Gouzie, D.; McAliley, M.L. State of remediation and metal toxicity in the tri-state mining district, USA. Chemosphere 2016, 144, 1132–1141. [Google Scholar] [CrossRef] [PubMed]
- Nacke, H.; Gonçalves, A.; Schwantes, D.; Nava, I.; Strey, L.; Coelho, G. Availability of heavy metals (Cd, Pb, and Cr) in agriculture from commercial fertilizers. Arch. Environ. Contam. Toxicol. 2013, 64, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Deocampo, D.M.; Reed, P.J.; Kalenuik, A.P. Road dust lead (Pb) in two neighborhoods of urban Atlanta, (GA, USA). Int. J. Environ. Res. Public Health 2012, 9, 2020–2030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solt, M.J.; Deocampo, D.M.; Norris, M. Spatial distribution of lead in Sacramento, California, USA. Int. J. Environ. Res. Public Health 2015, 12, 3174–3187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mielke, H.W.; Conzales, C.R.; Powell, E.T.; Mielke, P.W. Spatiotemporal dynamic transformations of soil lead and children’s blood lead ten years after Hurricane Katrina: New grounds for primary prevention. Environ. Int. 2016, 94, 567–575. [Google Scholar] [CrossRef]
- EPA (Environmental Protection Agency). Cleaning Up the Nation’s Waste Sites: Markets and Technology Trends; EPA/542/R-96/005; Office of Solid Waste and Emergency Response: Washington, DC, USA, 1997.
- Nevin, R. How lead exposure relates to temporal changes in IQ, violent crime, and unwed pregnancy. Environ. Res. 2000, 83, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Canfield, R.L.; Henderson, R.; Cory-Slechta, D.A.; Cox, C.; Jusko, T.A.; Lanphear, B.P. Intellectual impairment in children with blood lead concentrations below 10 μg per deciliter. N. Engl. J. Med. 2003, 348, 1517–1526. [Google Scholar] [CrossRef] [Green Version]
- Skerfving, S.; Löfmark, L.; Lundh, T.; Mikoczy, Z.; Strömberg, U. Late effects of low blood lead concentrations in children on school performance and cognitive functions. Neurotoxicology 2015, 49, 114–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakulski, K.M.; Rozek, L.S.; Dolinoy, D.C.; Paulson, H.L.; Hu, H. Alzheimer’s disease and environmental exposure to lead: The epidemiologic evidence and potential role of epigenetics. Curr. Alzheimer Res. 2012, 9, 563–573. [Google Scholar] [CrossRef]
- Chen, H.; Kwong, J.C.; Copes, R.; Tu, K.; Villeneuve, P.J.; van Donkelaar, A.; Hystad, P.; Martin, R.V.; Murray, B.J.; Jessiman, B.; et al. Living near major roads and the incidence of dementia, Parkinson’s disease, and multiple sclerosis: A population-based cohort study. Lancet 2017, 389, 718–726. [Google Scholar] [CrossRef]
- Fathabadi, B.; Dehghanifiroozabadi, M.; Aaseth, J.; Sharifzadeh, G.; Nakhaee, S.; Rajabpour-Sanati, A.; Amirabadizadeh, A.R.; Mehrpour, O. Comparison of blood lead levels in patients with Alzheimer’s Disease and healthy people. Am. J. Alzheimer’s Dis. Other Dement. 2018, 33, 541–547. [Google Scholar] [CrossRef]
- Malcoe, L.H.; Lynch, R.A.; Keger, M.C.; Skaggs, V.J. Lead sources, behaviors, and socioeconomic factors in relation to blood lead of Native American and white children: A community-based assessment of a former mining area. Environ. Health Perspect. 2002, 110, 221–231. [Google Scholar] [CrossRef]
- Levin, R.; Brown, M.J.; Kashtock, M.E.; Jacobs, D.E.; Whelan, E.A.; Rodman, J.; Schock, M.R.; Padilla, A.; Sinks, T. Lead exposures in US children, 2008: Implications for prevention. Environ. Health Perspect. 2008, 116, 1285–1293. [Google Scholar] [CrossRef] [Green Version]
- Sen, A.; Heredia, N.; Senut, M.C.; Land, S.; Hollocher, K.; Lu, X.; Dereski, M.O.; Ruden, D.M. Multigenerational epigenetic inheritance in humans: DNA methylation changes associated with maternal exposure to lead can be transmitted to the grandchildren. Sci. Rep. 2015, 5, 14466. [Google Scholar] [CrossRef] [Green Version]
- Paulson, J.A.; Brown, M.J. The CDC blood lead reference value for children: Time for a change. Environ. Health 2019, 18, 16. [Google Scholar] [CrossRef] [Green Version]
- Jabeen, R.; Ahmad, A.; Iqbal, M. Phytoremediation of heavy metals: Physiological and molecular mechanisms. Bot. Rev. 2009, 75, 339–364. [Google Scholar] [CrossRef]
- Morgano, M.A.; Teixeira Martins, M.C.; Rabonato, L.C.; Milani, R.F.; Yotsuyanagi, K.; Rodriguez-Amaya, D.B. Inorganic contaminants in bee pollen from Southeastern Brazil. J. Agric. Food Chem. 2010, 58, 6876–6883. [Google Scholar] [CrossRef] [PubMed]
- Lambert, O.; Piroux, M.; Puyo, S.; Thorin, C.; Larhantec, M.; Delbac, F.; Pouliquen, H. Bees, honey and pollen as sentinels for lead environmental contamination. Environ. Pollut. 2012, 170, 254–259. [Google Scholar] [CrossRef]
- Van der Merwe, D.; Carpenter, J.W.; Nietfeld, J.C.; Miesner, J.F. Adverse health effects in Canada geese (Branta canadensis) associated with waste from zinc and lead mines in the Tri-State Mining District (Kansas, Oklahoma, and Missouri, USA). J. Wildl. Dis. 2011, 47, 650–660. [Google Scholar] [CrossRef] [PubMed]
- Beyer, W.N.; Franson, J.C.; French, J.B.; May, T.; Rattner, B.A.; Shearn-Bochsler, V.I.; Warner, S.E.; Weber, J.; Mosby, D. Toxic exposure of songbirds to lead in the southeast Missouri lead mining district. Arch. Environ. Contam. Toxicol. 2013, 65, 598–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greipsson, S. Phytoremediation. Nat. Educ. Knowl. 2011, 13, 7. [Google Scholar]
- Peer, W.; Baxter, I.; Richards, E.; Freeman, J.; Murphy, A. Phytoremediation and hyperaccumulator plants. In Molecular Biology of Metal Homeostasis and Detoxification; Topics in Current Genetics; Tamas, M.J., Martinoia, E., Klomp, L.W.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 14, pp. 299–340. [Google Scholar]
- Evangelou, M.W.; Ebel, M.; Schaeffer, A. Chelate assisted phytoextraction of heavy metals from soil, effect, mechanism, toxicity, and fate of chelating agents. Chemosphere 2007, 68, 989–1003. [Google Scholar] [CrossRef]
- Zimdahl, R.L.; Skogerboe, R.K. Behavior of lead in soil. Environ. Sci. Technol. 1977, 11, 1202–1207. [Google Scholar] [CrossRef]
- Schooley, T.; Weaver, M.; Mullins, D.; Eick, M. The history of lead arsenate use in apple production: Comparison of its impact in Virginia with other states. J. Pestic. Saf. Educ. 2009, 10, 22–53. [Google Scholar]
- Hovsepyan, A.; Greipsson, S. EDTA-enhanced phytoremediation of lead contaminated soil by corn. J. Plant. Nutr. 2005, 28, 2037–2048. [Google Scholar] [CrossRef]
- López, M.L.; Peralta-Videa, J.R.; Benitez, T.; Gardea-Torresdey, J.L. Enhancement of lead uptake by alfalfa (Medicago sativa) using EDTA and a plant growth promoter. Chemosphere 2005, 61, 595–598. [Google Scholar] [CrossRef]
- Bolan, N.; Kunhikrishnan, A.; Thangarajan, R.; Kumpiene, J.; Park, J.; Makino, T.; Kirkham, M.B.; Scheckel, K. Remediation of heavy metal(oid)s contaminated soil—To mobilize or to immobilize? J. Hazard. Mater. 2014, 266, 141–166. [Google Scholar] [CrossRef]
- Jeffries, P.; Gianinazzi, S.; Perotto, S.; Turnau, K.; Barea, J.M. The contribution of arbuscular mycorrhizal fungi in sustainable maintenance of plant health and soil fertility. Biol. Fertil. Soils 2003, 37, 1–16. [Google Scholar] [CrossRef]
- Reynolds, H.L.; Vogelsang, K.M.; Hartley, A.E.; Bever, J.D.; Schultz, P.A. Variable responses of old-field perennials to arbuscular mycorrhizal fungi and phosphorus source. Oecologia 2006, 147, 348–358. [Google Scholar] [CrossRef] [PubMed]
- Leung, H.-M.; Wang, Z.-W.; Ye, Z.-H.; Yung, K.-L.; Peng, X.-L.; Cheung, K.-C. Interactions between arbuscular mycorrhizae and plants in phytoremediation of metal-contaminated soils: A review. Pedosphere 2013, 23, 549–563. [Google Scholar] [CrossRef]
- Van der Heijden, M.G.A.; Martin, F.M.; Selosse, M.; Sanders, I.R. Mycorrhizal ecology and evolution: The past, the present, and the future. New Phytol. 2015, 205, 1406–1423. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.B. Differences among mycorrhizal fungi for mineral uptake per root length of switchgrass grown in acidic soil. J. Plant. Nutr. 2002, 25, 1753–1772. [Google Scholar] [CrossRef]
- Cabral, L.; Soares, C.R.F.S.; Giachini, A.J.; Siqueira, J.O. Arbuscular mycorrhizal fungi in phytoremediation of contaminated areas by trace elements: Mechanisms and major benefits of their applications. World J. Microbiol. Biotechnol. 2015, 31, 1655–1664. [Google Scholar] [CrossRef] [PubMed]
- Wang, F. Occurrence of arbuscular mycorrhizal fungi in mining impacted sites and their contribution to ecological restoration: Mechanisms and application. Crit. Rev. Environ. Sci. Technol. 2017, 47, 1901–1957. [Google Scholar] [CrossRef]
- Xu, Z.; Ban, Y.; Li, Z.; Chen, H.; Yang, R.; Tang, M. Arbuscular mycorrhizal fungi play a role in protecting roots from Pb damage associated with increased phytochelatin synthase gene expression. Environ. Sci. Pollut. Res. 2014, 22, 12671–12683. [Google Scholar] [CrossRef] [PubMed]
- Calonne, M.; Fontaine, J.; Debiane, D.; Laruelle, F.; Grandmougin, A.; Lounes-Hadj, S.A. Side effects of the sterol biosynthesis inhibitor fungicide, propiconazole, on a beneficial arbuscular mycorrhizal fungus. Commun. Agric. Appl. Biol. Sci. 2010, 76, 891–902. [Google Scholar]
- Hovsepyan, A.; Greipsson, S. Effect of arbuscular mycorrhizal fungi on phytoextraction by corn (Zea mays) of lead-contaminated soil. Int. J. Phytoremediat. 2004, 6, 305–321. [Google Scholar] [CrossRef]
- Perry, V.R.; Krogstad, E.J.; El-Mayas, H.; Greipsson, S. Chemically enhanced phytoextraction of lead-contaminated soils. Int. J. Phytoremediat. 2012, 14, 703–713. [Google Scholar] [CrossRef]
- Namdjoyan, S.; Kermanian, H. Exogenous nitric oxide (as sodium nitroprusside) ameliorates arsenic-induced oxidative stress in watercress (Nasturtium officinale R. Br.) plants. Sci. Hortic. 2013, 161, 350–356. [Google Scholar] [CrossRef]
- Hadi, F.; Bano, A.; Fuller, M.P. The improved phytoextraction of lead (Pb) and the growth of maize (Zea mays L.): The role of plant growth regulators (GA3 and IAA) and EDTA alone and in combinations. Chemosphere 2010, 80, 457–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neill, S.J.; Desikan, R.; Clarke, A.; Hurst, R.D.; Hancock, J.T. Hydrogen peroxide and nitric oxide as signaling molecules in plants. J. Exp. Bot. 2002, 53, 1237–1247. [Google Scholar] [CrossRef] [PubMed]
- Khan, J.; Malangisha, G.K.; Ali, A.; Mahmoud, A.; Yang, J.; Zhang, M.; Hu, Z. Nitric oxide alleviates lead toxicity by inhibiting lead translocation and regulating root growth in watermelon seedlings. Hortic. Environ. Biotechnol. 2021. [Google Scholar] [CrossRef]
- Graziano, M.; Beligni, M.V.; Lamattina, L. Nitric oxide improves internal iron availability in plants. Plant. Physiol. 2002, 130, 1852–1859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graziano, M.; Lamattina, L. Nitric oxide and iron in plants: An emerging and converging story. Trends Plant. Sci. 2005, 10, 4–8. [Google Scholar] [CrossRef] [PubMed]
- Besson-Bard, A.; Gravot, A.; Richaud, P.; Auroy, P.; Duc, C.; Gaymard, F.; Taconnat, L.; Renou, J.P.; Pugin, A.; Wendehenne, D. Nitric oxide contributes to cadmium toxicity in Arabidopsis by promoting cadmium accumulation in roots and by up-regulating genes related to iron uptake. Plant. Physiol. 2009, 149, 1302–1315. [Google Scholar] [CrossRef] [Green Version]
- Lehotai, N.; Petô, A.; Weisz, M.; Erdei, L.; Kolbert, Z. Generation of reactive oxygen and nitrogen species in pea cultivars under copper exposure. Acta Bioligica. Szeged. 2011, 55, 273–278. [Google Scholar]
- Phang, C.; Leung, D.W.; Taylor, H.H.; Burritt, D.J. The protective effect of sodium nitroprusside (SNP) treatment on Arabidopsis thaliana seedlings exposed to toxic levels of Pb is not linked to avoidance of Pb uptake. Ecotoxicol. Environ. Saf. 2011, 74, 1310–1315. [Google Scholar] [CrossRef]
- Mihailovic, N.; Drazic, G. Incomplete alleviation of nickel toxicity in bean by nitric oxide supplementation. Plant. Soil Environ. 2011, 57, 396–401. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, C.; Vandenberghe, L.P.S.; de Oliveira, J.; Soccol, C.R. New perspectives of gibberellic acid production: A review. Crit. Rev. Biotechnol. 2012, 32, 263–273. [Google Scholar] [CrossRef] [PubMed]
- He, S.; He, Z.; Wu, Q.; Lei, W.; Zhang, X. Effects of GA3 on plant physiological properties, extraction, subcellular distribution and chemical forms of Pb in Lolium perenne. Int. J. Phytoremediat. 2015, 17, 1153–1159. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, A.M. Phytoextraction of Lead from Contaminated Soil by Panicum virgatum L. and Associated Growth Responses. Master’s Thesis, Department of Biology, Queen’s University, Kingston, ON, Canada, 2007. [Google Scholar]
- Parrish, D.J.; Fike, J.H. The biology and agronomy of switchgrass for biofuels. Crit. Rev. Plant. Sci. 2005, 24, 423–459. [Google Scholar] [CrossRef]
- Briske, D.D. Developmental morphology and physiology of grasses. In Grazing Management: An Ecological Perspective; Timber Press: Portland, OR, USA, 1991; pp. 85–108. [Google Scholar]
- Levy, D.B.; Redente, E.F.; Uphoff, G.D. Evaluating the phytotoxicity of Pb-Zn tailings to big bluestem (Andropogon gerardii Vitman) and switchgrass (Panicum virgatum L.). Soil Sci. 1999, 164, 363–375. [Google Scholar] [CrossRef]
- Smith, L.L.; Allen, D.J.; Barney, J.N. Yield potential and stand establishment for 20 candidate bioenergy feedstocks. Biomass Bioenergy 2015, 73, 145–154. [Google Scholar] [CrossRef]
- Lewandowski, I.; Scurlock, J.M.; Lindvall, E.; Christou, M. The development and current status of perennial rhizomatous grasses as energy crops in the US and Europe. Biomass Bioenergy 2003, 25, 335–361. [Google Scholar] [CrossRef]
- Balsamo, R.A.; Kelly, W.J.; Satrio, J.A.; Ruiz-Felix, M.N.; Fetterman, M.; Wynn, R.; Hagel, K. Utilization of grasses for potential biofuel production and phytoremediation of heavy metal contaminated soils. Int. J. Phytoremediat. 2014, 17, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.G.; Li, X.D.; Wang, C.C.; Chen, H.M.; Chua, H. Lead phytoextraction from contaminated soil with high-biomass plant species. J. Environ. Qual. 2002, 31, 1893–1900. [Google Scholar] [CrossRef] [PubMed]
- Van Ginneken, L.; Meers, E.; Guisson, R.; Ruttens, A.; Elst, K.; Tack, F.M.G.; Vangronsveld, J.; Diels, L.; Dejonghe, W. Phytoremediation for heavy metal-contaminated soils combined with bioenergy production. J. Environ. Eng. Landsc. Manag. 2007, 15, 227–236. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Allica, J.; Becerril, J.M.; Garbisu, C. Assessment of the phytoextraction potential of high biomass crop plants. Environ. Pollut. 2008, 152, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.C.; Lai, H.Y.; Juang, K.W. Model evaluation of plant metal content and biomass yield for the phytoextraction of heavy metals by switchgrass. Ecotoxicol. Environ. Saf. 2012, 80, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Oh, K.; Li, T.; Cheng, H.; Hu, X.; He, C.; Yan, L.; Shinichi, Y. Development of profitable phytoremediation of contaminated soils with biofuel crops. J. Environ. Prot. 2013, 4, 58–64. [Google Scholar] [CrossRef]
- Qin, Z.; Zhaung, Q.; Cai, X. Bioenergy crop productivity and potential climate change mitigation from marginal lands in the United States: An ecosystem modeling perspective. GCB Bioenergy 2015, 7, 1211–1221. [Google Scholar] [CrossRef] [Green Version]
- McLaughlin, S.B.; Kszos, L.A. Development of switchgrass (Panicum virgatum) as a bioenergy feedstock in the United States. Biomass Bioenergy 2005, 28, 515–535. [Google Scholar] [CrossRef]
- Hancock, D.W. The Management and Use of Switch-Grass in Georgia; Extension Service Bulletin 1358; University of Georgia, College of Agricultural and Environmental Sciences: Athens, Georgia, 2009. [Google Scholar]
- Rose, S. Anion adsorption and desorption characteristics of a piedmont ultisol: Some implications for the fate of sulfate deposition. Water Air Soil Pollut. 1998, 101, 333–347. [Google Scholar] [CrossRef]
- Franklin, D.H.; West, L.T.; Radcliffe, D.E.; Hendrix, P.F. Characteristics and genesis of preferential flow paths in a piedmont ultisol. Soil Sci. Soc. Am. J. 2007, 71, 752–758. [Google Scholar] [CrossRef]
- Northcott, G.L.; Jones, K.C. Experimental approaches and analytical techniques for determining organic compound bound residues in soil and sediment. Environ. Pollut. 2000, 108, 19–43. [Google Scholar] [CrossRef]
- Ötvös, K.; Pasternak, T.P.; Miskolczi, P.; Domoki, M.; Dorjgotov, D.; Bottka, S.; Dudits, D.; Fehér, A. Nitric oxide is required for, and promotes auxin-mediated activation of, cell division and embryogenic cell formation but does not influence cell cycle progression in alfalfa cell cultures. Plant. J. 2005, 43, 849–860. [Google Scholar] [CrossRef]
- Gupta, D.K.; Huang, H.G.; Corpas, F.J. Lead tolerance in plants: Strategies for phytoremediation. Environ. Sci. Pollut. Res. 2013, 20, 2150–2161. [Google Scholar] [CrossRef]
- Saxena, I.; Shekhawat, G.S. Nitric oxide (NO) in alleviation of heavy metal induced phytotoxicity and its role in protein nitration. Nitric Oxide 2013, 32, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Koske, R.E.; Gemma, J.N. A modified procedure for staining roots to detect VA mycorrhizas. Mycol. Res. 1989, 92, 486–488. [Google Scholar] [CrossRef]
- Sun, X.G.; Tang, M. Comparison of four routinely used methods for assessing root colonization by arbuscular mycorrhizal fungi. Botany 2012, 90, 1073–1083. [Google Scholar] [CrossRef]
- McGrath, S.P.; Zhao, F.J. Phytoextraction of metals and metalloids from contaminated soils. Curr. Opin. Biotechnol. 2003, 14, 277–282. [Google Scholar] [CrossRef]
- Sun, Y.; Zhou, Q.; An, J.; Liu, W.; Liu, R. Chelator-enhanced phytoextraction of heavy metals from contaminated soil irrigated by industrial wastewater with the hyperaccumulator plant (Sedum alfredii Hance). Geoderma 2009, 150, 106–112. [Google Scholar] [CrossRef]
- Ramirez, L.; Simontacchi, M.; Murgia, I.; Zabaleta, E.; Lamattina, L. Nitric oxide, nitrosyl iron complexes, ferritin and frataxin: A well-equipped team to preserve plant iron homeostasis. Plant. Sci. 2011, 181, 582–592. [Google Scholar] [CrossRef]
- Kopyra, M.; Gwóźdź, E.A. Nitric oxide stimulates seed germination and counter active inhibitory effect of heavy metals and salinity on root growth of Lupinus luteus. Plant. Physiol. Biochem. 2003, 41, 1011–1017. [Google Scholar] [CrossRef]
- Sarath, G.; Bethke, P.C.; Jones, R.; Baird, L.M.; Hou, G.; Mitchell, R.B. Nitric oxide accelerates seed germination in warm-season grasses. Planta 2006, 223, 1154–1164. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.; Deocampo, D.; El-Mayas, H.; Greipsson, S. Induced phytoextraction of lead through chemical manipulation of switchgrass and corn; role of iron supplement. Int. J. Phytoremediat. 2016, 17, 1192–1203. [Google Scholar] [CrossRef]
- Aderholt, M.; Vogelien, D.L.; Koether, M.; Greipsson, S. Phytoextraction of contaminated urban soil by Panicum virgatum L. enhanced with application of a plant growth regulator (BAP) and citric acid. Chemosphere 2017, 175, 85–96. [Google Scholar] [CrossRef]
- Bucheli-Witschel, M.; Egli, T. Environmental fate and microbial degradation of aminopolycarboxylic acids. FEMS Microbiol. Rev. 2001, 25, 69–106. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Zhao, F.J.; Lombi, E.; McGrath, S.P. Leaching of heavy metals from contaminated soils using EDTA. Environ. Pollut. 2001, 113, 111–120. [Google Scholar] [CrossRef]
- Oviedo, C.; Rodríguez, J. EDTA: The chelating agent under environmental scrutiny. Quim. Nova 2003, 26, 901–905. [Google Scholar] [CrossRef] [Green Version]
- de Araújo, J.D.C.T.; do Nascimento, C.W.A. Phytoextraction of lead from soil from a battery recycling site: The use of citric acid and NTA. Water Air Soil Pollut. 2010, 211, 113–120. [Google Scholar] [CrossRef]
- Geebelen, W.; Vangronsveld, J.; Adriano, D.C.; Van Poucke, L.C.; Clijsters, H. Effects of Pb-EDTA and EDTA on oxidative stress reactions and mineral uptake in Phaseolus vulgaris. Physiol. Plant. 2002, 115, 377–384. [Google Scholar] [CrossRef]
- Hasegawa, H.; Rahman, M.A.; Saitou, K.; Kobayashi, M.; Okumura, C. Influence of chelating ligands on bioavailability and mobility of iron in plant growth media and their effect on radish growth. Environ. Exp. Bot. 2011, 71, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Hart, G.; Koether, M.; McElroy, T.; Greipsson, S. Evaluation of chelating agents used in phytoextraction by switchgrass of lead (Pb) contaminated soils. J. Environ. Chem. Eng. 2021, submitted. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beavers, A.; Koether, M.; McElroy, T.; Greipsson, S. Effects of Exogenous Application of Plant Growth Regulators (SNP and GA3) on Phytoextraction by Switchgrass (Panicum virgatum L.) Grown in Lead (Pb) Contaminated Soil. Sustainability 2021, 13, 10866. https://doi.org/10.3390/su131910866
Beavers A, Koether M, McElroy T, Greipsson S. Effects of Exogenous Application of Plant Growth Regulators (SNP and GA3) on Phytoextraction by Switchgrass (Panicum virgatum L.) Grown in Lead (Pb) Contaminated Soil. Sustainability. 2021; 13(19):10866. https://doi.org/10.3390/su131910866
Chicago/Turabian StyleBeavers, Adrianne, Marina Koether, Thomas McElroy, and Sigurdur Greipsson. 2021. "Effects of Exogenous Application of Plant Growth Regulators (SNP and GA3) on Phytoextraction by Switchgrass (Panicum virgatum L.) Grown in Lead (Pb) Contaminated Soil" Sustainability 13, no. 19: 10866. https://doi.org/10.3390/su131910866
APA StyleBeavers, A., Koether, M., McElroy, T., & Greipsson, S. (2021). Effects of Exogenous Application of Plant Growth Regulators (SNP and GA3) on Phytoextraction by Switchgrass (Panicum virgatum L.) Grown in Lead (Pb) Contaminated Soil. Sustainability, 13(19), 10866. https://doi.org/10.3390/su131910866