
Riparian Ecological Infrastructures: Potential for Biodiversity-Related Ecosystem Services in Mediterranean Human-Dominated Landscapes


 , ,
, ,  , ,
, ,  and
and
Abstract
1. Introduction
- Characterize the structural attributes of existing Riparian Ecological Infrastructures (REIs) in three distinct Mediterranean human-dominated landscapes: Intensive Agriculture (IA), Extensive Agriculture (EA), and Forest Production (FP);
- Develop a new suitability metric, based on the floristic composition of riparian vegetation, and use it as a proxy to evaluate the potential of REIs in supporting seed dispersal and pollination Ecosystem Services (ES) provided by three biological dispersal groups: short-range dispersers (represented by ants), medium-range dispersers (represented by pollinators), and long-range dispersers (represented by birds and non-flying small mammals);
- Derive a novel multimetric index, termed Habitat Ecological Infrastructure’s Diversity Index (HEIDI), by integrating metrics related to the structure and management of riparian vegetation with the new suitability metric, and use it to estimate the potential of REIs in supporting biodiversity and promoting the ES provided by the three biological dispersal groups in each landscape.
2. Materials and Methods
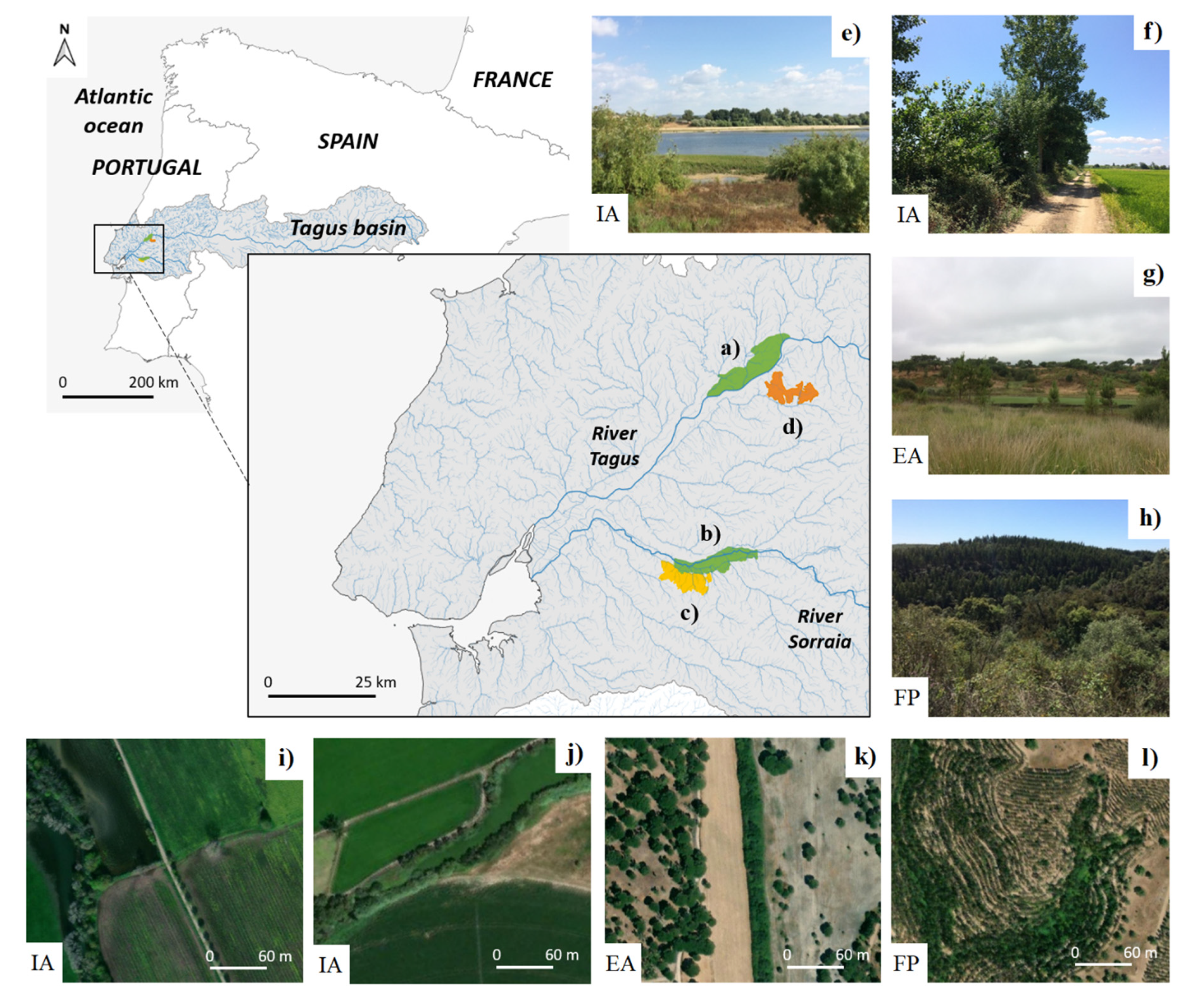
2.1. Study Area
2.2. Riparian Ecological Infrastructures (REIs)
2.3. REI’s Structural Attributes
2.4. Field Sampling
2.5. Habitat Ecological Infrastructure’s Diversity Index (HEIDI)
2.6. HEIDI Calculation
2.6.1. HEIDI Scoring System
2.6.2. HEIDI Estimation
2.7. Statistical Analysis
3. Results
3.1. REI’s Structural Attributes
3.2. Global HEIDI Results
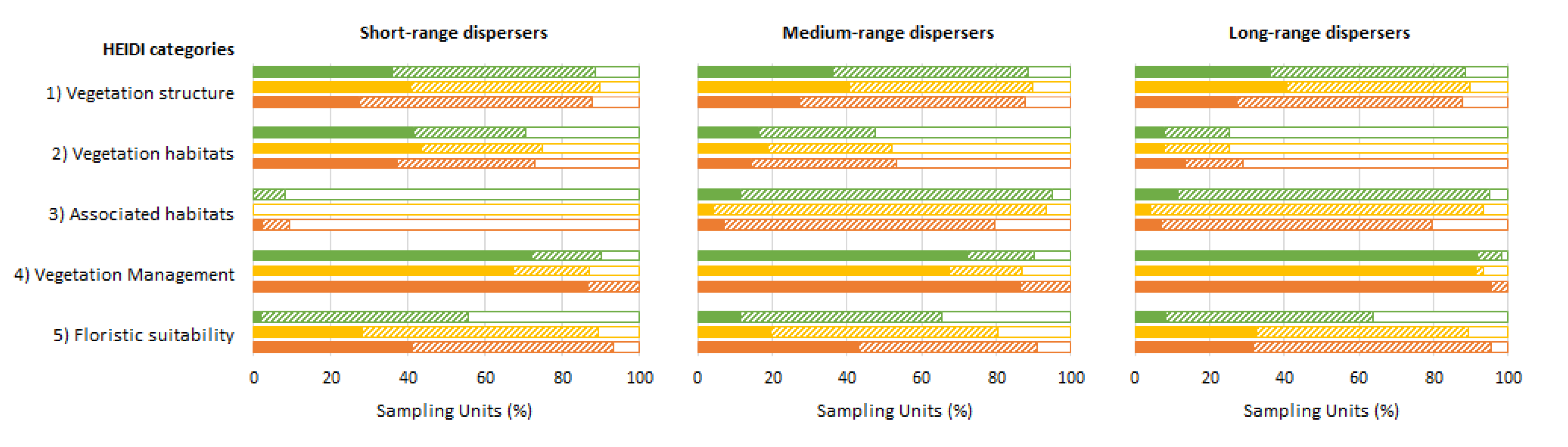
3.3. HEIDI Results by Category
3.3.1. Vegetation Structure, Vegetation Habitats, Associated Habitats, and Vegetation Management
3.3.2. Floristic Suitability
4. Discussion
4.1. Relevance of REI’s Structural Attributes for Biodiversity
4.2. Floristic Suitability to Support Biodiversity-Related ES
4.2.1. Seed Dispersal by Short-Range Dispersers
4.2.2. Pollination by Medium-Range Dispersers
4.2.3. Seed Dispersal by Long-Range Dispersers
4.3. REI’s Potential for Biological Dispersal in the Three Human-Dominated Mediterranean Landscapes
4.4. HEIDI Synthesis and Applications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ward, J.V.; Tockner, K.; Schiemer, F. Biodiversity of floodplain river ecosystems: Ecotones and connectivity. Regul. Rivers Res. Manag. 1999, 15, 125–139. [Google Scholar] [CrossRef]
- Naiman, R.J.; Decamps, H.; Pollock, M. The role of riparian corridors in maintaining regional biodiversity. Ecol. Appl. 1993, 3, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Sabo, J.L.; Sponseller, R.; Dixon, M.; Gade, K.; Harms, T.; Heffernan, J.; Jani, A.; Katz, G.; Soykan, C.; Watts, J.; et al. Riparian zones increase regional species richness by harboring different, not more, species. Ecology 2005, 86, 56–62. [Google Scholar] [CrossRef]
- Capon, S.J.; Chambers, L.E.; Mac Nally, R.; Naiman, R.J.; Davies, P.; Marshall, N.; Pittock, J.; Reid, M.; Capon, T.; Douglas, M.; et al. Riparian ecosystems in the 21st century: Hotspots for climate change adaptation? Ecosystems 2013, 16, 359–381. [Google Scholar] [CrossRef]
- Riis, T.; Kelly-Quinn, M.; Aguiar, F.C.; Manolaki, P.; Bruno, D.; Bejarano, M.D.; Clerici, N.; Fernandes, M.R.; Franco, J.C.; Pettit, N.; et al. Global overview of ecosystem services provided by riparian vegetation. BioScience 2020, 70, 501–514. [Google Scholar] [CrossRef]
- Naiman, R.J.; Décamps, H. The ecology of interfaces: Riparian zones. Annu. Rev. Ecol. Syst. 1997, 28, 621–658. [Google Scholar] [CrossRef]
- Corbacho, C.; Sánchez, J.M.; Costillo, E. Patterns of structural complexity and human disturbance of riparian vegetation in agricultural landscapes of a Mediterranean area. Agric. Ecosyst. Environ. 2003, 95, 495–507. [Google Scholar] [CrossRef]
- Tockner, K.; Bunn, S.; Gordon, C.; Naiman, J.; Robert, P.Q.; Gerry, A.; Stanford, J. Flood plains: Critically threatened ecosystems. In Aquatic Ecosystems. Trends and Global Prospects; Cambridge University Press: Cambridge, UK, 2008; pp. 45–61. [Google Scholar]
- Fielding, K.S.; Terry, D.J.; Masser, B.M.; Bordia, P.; Hogg, M.A. Explaining landholders’ decisions about riparian zone management: The role of behavioural, normative, and control beliefs. J. Environ. Manag. 2005, 77, 12–21. [Google Scholar] [CrossRef]
- Pereira, M.; Rodríguez, A. Conservation value of linear woody remnants for two forest carnivores in a Mediterranean agricultural landscape. J. Appl. Ecol. 2010, 47, 611–620. [Google Scholar] [CrossRef]
- Fremier, A.K.; Kiparsky, M.; Gmur, S.; Aycrigg, J.; Craig, R.K.; Svancara, L.K.; Goble, D.D.; Cosens, B.; Davis, F.W.; Scott, J.M. A riparian conservation network for ecological resilience. Biol. Conserv. 2015, 191, 29–37. [Google Scholar] [CrossRef]
- Van Looy, K.; Tormos, T.; Souchon, Y.; Gilvear, D. Analyzing riparian zone ecosystem services bundles to instruct river management. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2017, 13, 330–341. [Google Scholar] [CrossRef]
- Capon, S.J.; Pettit, N.E. Turquoise is the new green: Restoring and enhancing riparian function in the Anthropocene. Ecol. Manag. Restor. 2018, 19, 44–53. [Google Scholar] [CrossRef]
- Feld, C.K.; Fernandes, M.R.; Ferreira, M.T.; Hering, D.; Ormerod, S.J.; Venohr, M.; Gutiérrez-Cánovas, C. Evaluating riparian solutions to multiple stressor problems in river ecosystems—A conceptual study. Water Res. 2018, 139, 381–394. [Google Scholar] [CrossRef]
- Cole, L.J.; Stockan, J.; Helliwell, R. Managing riparian buffer strips to optimise ecosystem services: A. review. Agric. Ecosyst. Environ. 2020, 296, 106891. [Google Scholar] [CrossRef]
- Jennings, M.K.; Haeuser, E.; Foote, D.; Lewison, R.L.; Conlisk, E. Planning for dynamic connectivity: Operationalizing robust decision-making and prioritization across landscapes experiencing climate and land-use change. Land 2020, 9, 341. [Google Scholar] [CrossRef]
- Lee, J.A.; Chon, J.; Ahn, C. Planning landscape corridors in ecological infrastructure using least-cost path methods based on the value of ecosystem services. Sustainability 2014, 6, 7564–7585. [Google Scholar] [CrossRef]
- Sun, S.; Jiang, Y.; Zheng, S. Research on ecological infrastructure from 1990 to 2018: A bibliometric analysis. Sustainability 2020, 12, 2304. [Google Scholar] [CrossRef]
- Stella, J.C.; Rodríguez-González, P.M.; Dufour, S.; Bendix, J. Riparian vegetation research in Mediterranean-climate regions: Common patterns, ecological processes, and considerations for management. Hydrobiologia 2013, 719, 291–315. [Google Scholar] [CrossRef]
- Aguiar, F.C.; Martins, M.J.; Silva, P.C.; Fernandes, M.R. Riverscapes downstream of hydropower dams: Effects of altered flows and historical land-use change. Landsc. Urban Plan. 2016, 153, 83–98. [Google Scholar] [CrossRef]
- Aguiar, F.C.; Ferreira, M.T. Human-disturbed landscapes: Effects on composition and integrity of riparian woody vegetation in the Tagus River basin, Portugal. Environ. Conserv. 2005, 32, 30–41. [Google Scholar] [CrossRef]
- Cooper, S.D.; Lake, P.S.; Sabater, S.; Melack, J.M.; Sabo, J.L. The effects of land use changes on streams and rivers in Mediterranean climates. Hydrobiologia 2013, 719, 383–425. [Google Scholar] [CrossRef]
- Fernandes, M.R.; Aguiar, F.C.; Martins, M.J.; Rivaes, R.; Ferreira, M.T. Long-term human-generated alterations of Tagus River: Effects of hydrological regulation and land-use changes in distinct river zones. Catena 2020, 188, 104466. [Google Scholar] [CrossRef]
- Plieninger, T.; Wilbrand, C. Land use, biodiversity conservation, and rural development in the dehesas of Cuatro Lugares, Spain. Agrofor. Syst. 2001, 51, 23–34. [Google Scholar] [CrossRef]
- Fonseca, A.; Ugille, J.-P.; Michez, A.; Rodríguez-González, P.M.; Duarte, G.; Ferreira, M.T.; Fernandes, M.R. Assessing the connectivity of Riparian Forests across a gradient of human disturbance: The potential of Copernicus “Riparian Zones” in two hydroregions. Forests 2021, 12, 674. [Google Scholar] [CrossRef]
- Langer, L.E.R.; Steward, G.A.; Kimberley, M.O. Vegetation structure, composition and effect of pine plantation harvesting on riparian buffers in New Zealand. For. Ecol. Manag. 2008, 256, 949–957. [Google Scholar] [CrossRef]
- Tererai, F.; Gaertner, M.; Jacobs, S.M.; Richardson, D.M. Eucalyptus invasions in riparian forests: Effects on native vegetation community diversity, stand structure and composition. For. Ecol. Manag. 2013, 297, 84–93. [Google Scholar] [CrossRef]
- Dufour, S.; Rodríguez-González, P.M.; Laslier, M. Tracing the scientific trajectory of riparian vegetation studies: Main topics, approaches and needs in a globally changing world. Sci. Total Environ. 2019, 653, 1168–1185. [Google Scholar] [CrossRef]
- Santos, M.J.; Rosalino, L.M.; Matos, H.M.; Santos-Reis, M. Riparian ecosystem configuration influences mesocarnivores presence in Mediterranean landscapes. Eur. J. Wildl. Res. 2016, 62, 251–261. [Google Scholar] [CrossRef]
- Gasith, A.; Resh, V.H. Streams in Mediterranean climate regions: Abiotic influences and biotic responses to predictable seasonal events. Annu. Rev. Ecol. Syst. 1999, 30, 51–81. [Google Scholar] [CrossRef]
- Cole, L.J.; Brocklehurst, S.; Robertson, D.; Harrison, W.; McCracken, D.I. Exploring the interactions between resource availability and the utilisation of semi-natural habitats by insect pollinators in an intensive agricultural landscape. Agric. Ecosyst. Environ. 2017, 246, 157–167. [Google Scholar] [CrossRef]
- Santos, A.; Fernandes, M.R.; Aguiar, F.C.; Branco, M.R.; Ferreira, M.T. Effects of riverine landscape changes on pollination services: A case study on the River Minho, Portugal. Ecol. Indic. 2018, 89, 656–666. [Google Scholar] [CrossRef]
- Kalcounis, M.C.; Hobson, K.A.; Brigham, R.M.; Hecker, K.R. Bat activity in the boreal forest: Importance of stand type and vertical strata. J. Mammal. 1999, 80, 673–682. [Google Scholar] [CrossRef]
- Gil-Tena, A.; Saura, S.; Brotons, L. Effects of forest composition and structure on bird species richness in a Mediterranean context: Implications for forest ecosystem management. For. Ecol. Manag. 2007, 242, 470–476. [Google Scholar] [CrossRef]
- Saura, S.; Rubio, L. A common currency for the different ways in which patches and links can contribute to habitat availability and connectivity in the landscape. Ecography 2010, 33, 523–537. [Google Scholar] [CrossRef]
- Taylor, P.D.; Fahrig, L.; Henein, K.; Merriam, G. Connectivity is a vital element of landscape structure. Oikos 1993, 68, 571–573. [Google Scholar] [CrossRef]
- Regnery, B.; Couvet, D.; Kubarek, L.; Julien, J.F.; Kerbiriou, C. Tree microhabitats as indicators of bird and bat communities in Mediterranean forests. Ecol. Indic. 2013, 34, 221–230. [Google Scholar] [CrossRef]
- Paillet, Y.; Archaux, F.; du Puy, S.; Bouget, C.; Boulanger, V.; Debaive, N.; Glig, O.; Gosselin, F.; Guilbert, E. The indicator side of tree microhabitats: A multi-taxon approach based on bats, birds and saproxylic beetles. J. Appl. Ecol. 2018, 55, 2147–2159. [Google Scholar] [CrossRef]
- Harrison, P.A.; Berry, P.M.; Simpson, G.; Haslett, J.R.; Blicharska, M.; Bucur, M.; Dunford, R.; Egoh, B.; Garcia-LIorente, M.; Geamana, N.; et al. Linkages between biodiversity attributes and ecosystem services: A systematic review. Ecosyst. Serv. 2014, 9, 191–203. [Google Scholar] [CrossRef]
- Folgarait, P.J. Ant biodiversity and its relationship to ecosystem functioning: A review. Biodivers. Conserv. 1998, 7, 1221–1244. [Google Scholar] [CrossRef]
- Del Toro, I.; Ribbons, R.R.; Pelini, S.L. The little things that run the world revisited. Myrmecol. News 2012, 14, 133–146. [Google Scholar]
- Gómez, C.; Espadaler, X. An update of the world survey of myrmecochorous dispersal distances. Ecography 2013, 36, 1193–1201. [Google Scholar] [CrossRef]
- Potts, S.G.; Petanidou, T.; Roberts, S.; O’Toole, C.; Hulbert, A.; Willmer, P. Plant-pollinator biodiversity and pollination services in a complex Mediterranean landscape. Biol. Conserv. 2006, 129, 519–529. [Google Scholar] [CrossRef]
- Klein, A.M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Howe, F.; Smallwood, J. Ecology of seed dispersal. Annu. Rev. Ecol. Syst. 1982, 13, 201–228. [Google Scholar] [CrossRef]
- Rosalino, L.M.; Rosa, S.; Santos-Reis, M. The role of carnivores as Mediterranean seed dispersers. Ann. Zool. Fenn. 2010, 47, 195–205. [Google Scholar] [CrossRef]
- Costa, J.M.; Ramos, J.A.; da Silva, L.P.; Timoteo, S.; Araújo, P.M.; Felgueiras, M.S.; Rosa, A.; Matos, C.; Encarnação, P.; Tenreiro, P.Q.; et al. Endozoochory largely outweighs epizoochory in migrating passerines. J. Avian Biol. 2014, 45, 59–64. [Google Scholar] [CrossRef]
- Boyles, J.G.; Cryan, P.M.; McCracken, G.F.; Kunz, T.H. Economic importance of bats in agriculture. Science 2011, 332, 41–42. [Google Scholar] [CrossRef]
- Kunz, T.H.; de Torrez, E.B.; Bauer, D.; Lobova, T.; Fleming, T.H. Ecosystem services provided by bats. Ann. N. Y. Acad. Sci. 2011, 1223, 1–38. [Google Scholar] [CrossRef]
- Santolini, R.; Morri, E.; Pasini, G.; Giovagnoli, G.; Morolli, C.; Salmoiraghi, G. Assessing the quality of riparian areas: The case of river ecosystem quality index applied to the Marecchia river (Italy). Int. J. River Basin Manag. 2015, 13, 1–16. [Google Scholar] [CrossRef]
- de Sosa, L.L.; Glanville, H.C.; Marshall, M.R.; Prysor Williams, A.; Jones, D.L. Quantifying the contribution of riparian soils to the provision of ecosystem services. Sci. Total Environ. 2018, 624, 807–819. [Google Scholar] [CrossRef]
- Hanna, D.E.L.; Tomscha, S.A.; Ouellet Dallaire, C.; Bennett, E.M. A review of riverine ecosystem service quantification: Research gaps and recommendations. J. Appl. Ecol. 2018, 55, 1299–1311. [Google Scholar] [CrossRef]
- Oliver, I.; Dorrough, J.; Seidel, J. A new vegetation integrity metric for trading losses and gains in terrestrial biodiversity value. Ecol. Indic. 2021, 124, 107341. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef]
- Vogt, J.; Soille, P.; De Jager, A.; Rimavičiūtė, E.; Mehl, W.; Foisneau, S.; Bodis, K.; Dusart, J.; Paracchini, M.L.; Haastrup, P.; et al. A Pan-European River and Catchment Database; OPOCE: Luxembourg, Luxemburg, 2007. [Google Scholar] [CrossRef]
- Fernandes, M.R.; Aguiar, F.C.; Ferreira, M.T. Assessing riparian vegetation structure and the influence of land use using landscape metrics and geostatistical tools. Landsc. Urban Plan. 2011, 99, 166–177. [Google Scholar] [CrossRef]
- Raunkiær, C. The Life Forms of Plants and Statistical Plant Geography, Being the Collected Papers of C. Raunkiær. History of Ecology Series; Frank, N.E., Ed.; Oxford University Press: Oxford, UK, 1934; ISBN 0-405-10418-9. [Google Scholar]
- Wasser, L.; Chasmer, L.; Day, R.; Taylor, A. Quantifying land use effects on forested riparian buffer vegetation structure using LiDAR data. Ecosphere 2015, 6, 1–17. [Google Scholar] [CrossRef]
- McGarigal, K.; Cushman, S.A.; Ene, E. FRAGSTATS v4: Spatial Pattern Analysis Program for Categorical and Continuous Maps. 2012. Available online: http://www.umass.edu/landeco/research/fragstats/fragstats.html (accessed on 21 September 2021).
- Larrieu, L.; Gonin, P. L’lndice de biodiversité potentielle (IBP): Une méthode simple et rapide pour évaluer la biodiversité potentielle des peuplements forestiers. Rev. For. Fr. 2008, 60, 727–748. [Google Scholar] [CrossRef]
- Johnson, R.R.; Steven, W.C.; Simpson, J.M. A riparian classification system. In California Riparian Systems: Ecology, Conservation, and Productive Management; Warner, R.E., Hendrix, K.M., Eds.; University of California Press: Berkeley, CA, USA, 1984. [Google Scholar]
- Lengyel, S.; Gove, A.D.; Latimer, A.M.; Majer, J.D.; Dunn, R.R. Convergent evolution of seed dispersal by ants, and phylogeny and biogeography in flowering plants: A global survey. Perspect. Plant. Ecol. Evol. Syst. 2010, 12, 43–55. [Google Scholar] [CrossRef]
- Herrera, C.M. Plant-vertebrate seed dispersal systems in the Mediterranean: Ecological, evolutionary, and historical determinants. Annu. Rev. Ecol. Syst. 1995, 26, 705–727. [Google Scholar] [CrossRef]
- Karr, J.R. Assessment of biotic integrity using fish communities. Fisheries 1981, 6, 21–27. [Google Scholar] [CrossRef]
- Silva, P.S.D.; Bieber, A.G.D.; Corrêa, M.M.; Leal, I.R. Do leaf-litter attributes affect the richness of leaf-litter ants? Neotrop. Entomol. 2011, 40, 542–547. [Google Scholar] [CrossRef]
- Lopes, J.F.S.; Hallack, N.M.D.R.; Sales, T.A.; De Brugger, M.S.; Ribeiro, L.F.; Hastenreiter, I.N.; Camargo, R.D.S. Comparison of the ant assemblages in three phytophysionomies: Rocky field, secondary forest, and riparian forest—A case study in the state park of Ibitipoca, Brazil. Psyche 2012. [Google Scholar] [CrossRef]
- Ives, C.D.; Taylor, M.P.; Nipperess, D.A.; Hose, G.C. Effect of catchment urbanization on ant diversity in remnant riparian corridors. Landsc. Urba. Plan. 2013, 110, 155–163. [Google Scholar] [CrossRef]
- Welch, J.N.; Leppanen, C. The threat of invasive species to bats: A review. Mammal. Rev. 2017, 47, 277–290. [Google Scholar] [CrossRef]
- Badalamenti, E.; Cusimano, D.; La Mantia, T.; Pasta, S.; Romano, S.; Troia, A.; Ilardi, V. The ongoing naturalisation of Eucalyptus spp. in the Mediterranean Basin: New threats to native species and habitats. Aust. For. 2018, 81, 239–249. [Google Scholar] [CrossRef]
- Quinn, L.D.; Holt, J.S. Ecological correlates of invasion by Arundo donax in three southern California riparian habitats. Biol. Invasions 2008, 10, 591–601. [Google Scholar] [CrossRef]
- Andersen, A.N. Responses of ant communities to disturbance: Five principles for understanding the disturbance dynamics of a globally dominant faunal group. J. Anim. Ecol. 2019, 88, 350–362. [Google Scholar] [CrossRef]
- Nooten, S.S.; Schultheiss, P.; Rowe, R.C.; Facey, S.L.; Cook, J.M. Habitat complexity affects functional traits and diversity of ant assemblages in urban green spaces (Hymenoptera: Formicidae). Myrmecol. News 2019, 29, 67–77. [Google Scholar] [CrossRef]
- Gonçalves, P.; Alcobia, S.; Simões, L.; Santos-Reis, M. Effects of management options on mammal richness in a Mediterranean agro-silvo-pastoral system. Agrofor. Syst. 2012, 85, 383–395. [Google Scholar] [CrossRef]
- Galkowski, C.L.C.; Wegnez, R.B.P. Guía de Campo de las Hormigas de Europa Occidental, Edición Española; Omega: Barcelona, Spain, 2017. [Google Scholar]
- da Rocha-Filho, L.C.; Montagnana, P.C.; Boscolo, D.; Garófalo, C.A. Green patches among a grey patchwork: The importance of preserving natural habitats to harbour cavity-nesting bees and wasps (Hymenoptera) and their natural enemies in urban areas. Biodivers. Conserv. 2020, 29, 2487–2514. [Google Scholar] [CrossRef]
- Glaser, F. Ants (Hymenoptera, Formicidae) in alpine floodplains—Ecological notes and conservation aspects. In Riverine Landscapes—Restoration—Flood Protection–Conservation, Proceedings of the Internationales LIFE-Symposium, Reutte-Breitenwang, Austria, 26–29 September 2005; Füreder, L., Sint, D., Vorauer, A., Eds.; Springer: Berlin, Germany, 2005; pp. 147–163. [Google Scholar]
- Rebelo, H.; Rainho, A. Bat conservation and large dams: Spatial changes in habitat use caused by Europe’s largest reservoir. Endanger. Species Res. 2008, 8, 61–68. [Google Scholar] [CrossRef]
- Amorim, F.; Jorge, I.; Beja, P.; Rebelo, H. Following the water? Landscape-scale temporal changes in bat spatial distribution in relation to Mediterranean summer drought. Ecol. Evol. 2018, 8, 5801–5814. [Google Scholar] [CrossRef]
- Perakis, M.; Maravelakis, P.E.; Psarakis, S.; Xekalaki, E.; Panaretos, J. On certain indices for ordinal data with unequally weighted classes. Qual. Quant. 2005, 39, 515–536. [Google Scholar] [CrossRef]
- Feio, M.J.; Aguiar, F.C.; Almeida, S.F.P.; Ferreira, J.; Ferreira, M.T.; Elias, C.; Serra, S.R.Q.; Buffagni, A.; Cambra, J.; Chauvin, C.; et al. Least disturbed condition for European Mediterranean rivers. Sci. Total Environ. 2014, 476, 745–756. [Google Scholar] [CrossRef] [PubMed]
- JASP Team. JASP (Version 0.14.1) [Computer software]. Amsterdam, The Netherlands. 2020. Available online: https://jasp-stats.org (accessed on 21 September 2021).
- Kontsiotis, V.; Zaimes, G.N.; Tsiftsis, S.; Kiourtziadis, P.; Bakaloudis, D. Assessing the influence of riparian vegetation structure on bird communities in agricultural Mediterranean landscapes. Agrofor. Syst. 2019, 93, 675–687. [Google Scholar] [CrossRef]
- Forio, M.A.E.; De Troyer, N.; Lock, K.; Witing, F.; Baert, L.; De Saeyer, N.; Rîșnoveanu, G.; Popescu, C.; Burdon, F.J.; Kupilas, B.; et al. Small patches of riparian woody vegetation enhance biodiversity of invertebrates. Water 2020, 12, 3070. [Google Scholar] [CrossRef]
- Fahrig, L. Effects of habitat fragmentation on biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef]
- Clerici, N.; Paracchini, M.L.; Maes, J. Land-cover change dynamics and insights into ecosystem services in European stream riparian zones. Ecohydrol. Hydrobiol. 2014, 14, 107–120. [Google Scholar] [CrossRef]
- Torralba, M.; Fagerholm, N.; Burgess, P.J.; Moreno, G.; Plieninger, T. Do European agroforestry systems enhance biodiversity and ecosystem services? A meta-analysis. Agric. Ecosyst. Environ. 2016, 230, 150–161. [Google Scholar] [CrossRef]
- Ferreira, V.; Boyero, L.; Calvo, C.; Correa, F.; Figueroa, R.; Gonçalves, J.F., Jr.; Graça, M.A.S.; Hepp, L.U.; Goyenola, G.; Kariuki, S.; et al. A global assessment of the effects of eucalyptus plantations on stream ecosystem functioning. Ecosystems 2019, 22, 629–642. [Google Scholar] [CrossRef]
- Azcárate, F.M.; Arqueros, L.; Sánchez, A.M.; Peco, B. Seed and fruit selection by harvester ants, Messor barbarus, in Mediterranean grassland and scrubland. Funct. Ecol. 2005, 19, 273–283. [Google Scholar] [CrossRef]
- Bastida, F.; Talavera, S.; Ortiz, P.L.; Arista, M. The interaction between Cistaceae and a highly specific seed-harvester ant in a Mediterranean scrubland. Plant Biol. 2009, 11, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Valero, E.; Álvarez, X.; Picos, J. An assessment of river habitat quality as an indicator of conservation status. A case study in the northwest of Spain. Ecol. Indic. 2015, 57, 131–138. [Google Scholar] [CrossRef]
- Zaimes, G.N.; Iakovoglou, V. Assessing riparian areas of Greece—An overview. Sustainability 2021, 13, 309. [Google Scholar] [CrossRef]
- Latsiou, A.; Kouvarda, T.; Stefanidis, K.; Papaioannou, G.; Gritzalis, K.; Dimitriou, E. Pressures and status of the riparian vegetation in Greek rivers: Overview and preliminary assessment. Hydrology 2021, 8, 55. [Google Scholar] [CrossRef]
- Pereira, P.; Godinho, C.; Gomes, M.; Rabaça, J.E. The importance of the surroundings: Are bird communities of riparian galleries influenced by agroforestry landscapes in SW Iberian Peninsula? Ann. For. Sci. 2014, 71, 33–41. [Google Scholar] [CrossRef][Green Version]
- Stephan, J.; Issa, D. Anthropogenic impacts on riparian trees and shrubs in an eastern Mediterranean stream. Plant. Sociol. 2017, 54, 43–50. [Google Scholar] [CrossRef]
- Carrilho, M.; Teixeira, D.; Santos-Reis, M.; Rosalino, L.M. Small mammal abundance in Mediterranean Eucalyptus plantations: How shrub cover can really make a difference. For. Ecol. Manag. 2017, 391, 256–263. [Google Scholar] [CrossRef]
- Guiomar, N.; Godinho, S.; Fernandes, P.M.; Machado, R.; Neves, N.; Fernandes, J.P. Wildfire patterns and landscape changes in Mediterranean oak woodlands. Sci. Total Environ. 2015, 536, 338–352. [Google Scholar] [CrossRef] [PubMed]
- Carlson, E.A.; Cooper, D.J.; Merritt, D.M.; Kondratieff, B.C.; Waskom, R.M. Irrigation canals are newly created streams of semi-arid agricultural regions. Sci. Total Environ. 2019, 646, 770–781. [Google Scholar] [CrossRef]
- Cunha, N.S.; Magalhães, M.R. Methodology for mapping the national ecological network to mainland Portugal: A planning tool towards a green infrastructure. Ecol. Indic. 2019, 104, 802–818. [Google Scholar] [CrossRef]
- Arroyo-Rodríguez, V.; Fahrig, L.; Tabarelli, M.; Watling, J.I.; Tischendorf, L.; Benchimol, M.; Cazetta, E.; Faria, D.; Leal, I.R.; Melo, F.P.L.; et al. Designing optimal human-modified landscapes for forest biodiversity conservation. Ecol. Lett. 2020, 23, 1404–1420. [Google Scholar] [CrossRef] [PubMed]
- Daily, G.C.; Polasky, S.; Goldstein, J.; Kareiva, P.M.; Mooney, H.A.; Pejchar, L.; Ricketts, T.H.; Salzman, J.; Shallenberger, R. Ecosystem services in decision making: Time to deliver. Front. Ecol. Environ. 2009, 7, 21–28. [Google Scholar] [CrossRef]
- Liedtke, J.; Fromhage, L. The joint evolution of learning and dispersal maintains intraspecific diversity in metapopulations. Oikos 2021, 130, 808–818. [Google Scholar] [CrossRef]
- Stephens, P.A.; Pettorelli, N.; Barlow, J.; Whittingham, M.J.; Cadotte, M.W. Management by proxy? The use of indices in applied ecology. J. Appl. Ecol. 2015, 52, 1–6. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| HEIDI Categories and Metrics | Short-Range Dispersers’ Scores | Medium-Range Dispersers’ Scores | Long-Range Dispersers’ Scores | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Low | Fair | High | References | Low | Fair | High | References | Low | Fair | High | References | |
| 1. Vegetation structure | ||||||||||||
| Native tree species (N°) | 0 | 1 to 3 | ≥4 | [67] | 0 | 1 to 3 | ≥4 | [32] | 0 | 1 to 3 | ≥4 | [10,68] |
| Invasive species cover (%) | ≥30 | ]0, 30[ | 0 | [67,69,70] | ≥30 | ]0, 30[ | 0 | [32,69,70] | ≥30 | ]0, 30[ | 0 | [68,69,70] |
| Vertical strata (N°) | 1 | 2 or 3 | 4 | [66,71,72] | 1 | 2 or 3 | 4 | [32] | 1 | 2 or 3 | 4 | [33,66,73] |
| 2. Vegetation habitats | ||||||||||||
| Trees with microhabitats above 3 m (N°) | 0 | 1 or 2 | ≥3 | [34,37] | ||||||||
| Trees with microhabitats below 3 m (N°) | 0 | 1 or 2 | ≥3 | [74] | ||||||||
| Standing dead trees (N°) | 0 | 1 or 2 | ≥3 | [75] | 0 | 1 or 2 | ≥3 | [34,38] | ||||
| Dead wood trunks on the ground (N°) | 0 | 1 or 2 | ≥3 | [74,76] | 0 | 1 or 2 | ≥3 | [68] | ||||
| Large living trees (N°) | 0 | 1 to 4 | ≥5 | [33,34,38] | ||||||||
| Leaf litter cover (%) | 0 | ]0, 50[ | ≥50 | [65,66] | ≥50 | ]0, 50[ | 0 | [32] | ||||
| 3. Associated habitats | ||||||||||||
| Rocky habitat types (N°) | 0 | 1 | ≥2 | [66,74] | ||||||||
| Aquatic habitat types (N°) | 0 | 1 | ≥2 | [32] | 0 | 1 | ≥2 | [77,78] | ||||
| 4. Vegetation management | ||||||||||||
| Understory clearing (%) | ≥60 | [20, 60[ | <20 | [66] | ≥60 | [20, 60[ | <20 | [32] | ||||
| Tree clearing (%) | ≥60 | [20, 60[ | <20 | [33,34,38] | ||||||||
| 5. Floristic suitability | ||||||||||||
| Seed production suitability (initial scores) | <3 | [3, 16] | >16 | * | ||||||||
| Pollen production suitability (initial scores) | <20 | [20, 38] | >38 | ** | ||||||||
| Fruit production suitability (initial scores) | <10 | [10, 23] | >23 | *** | ||||||||
| Landscape | River | Landscape Area (ha) | REI Class | NP | CA (ha) | MPS (ha) | CC (%) | MNN (m) | MSI |
|---|---|---|---|---|---|---|---|---|---|
| IA | Sorraia | 5455.10 | Woody | 173 | 162.85 | 0.94 (±3.45) | 2.98 | 34.15 (±58.40) | 3.07 (±2.38) |
| Non-woody | 165 | 30.70 | 0.19 (±0.33) | 0.56 | 93.54 (±194.27) | 2.79 (±1.20) | |||
| Tagus | 8473.59 | Woody | 109 | 237.74 | 2.18 (±8.27) | 2.81 | 49.53 (±73.03) | 3.42 (±2.63) | |
| Non-woody | 91 | 10.26 | 0.11 (±0.18) | 0.12 | 243.40 (±405.34) | 2.30 (±0.76) | |||
| EA | Sorraia | 4427.40 | Woody | 296 | 78.98 | 0.27 (±0.60) | 1.78 | 47.37 (±62.85) | 2.49 (±1.28) |
| Non-woody | 295 | 63.57 | 0.22 (±0.43) | 1.44 | 52.14 (±115.69) | 3.02 (±1.32) | |||
| FP | Tagus | 4204.00 | Woody | 379 | 283.83 | 0.75 (±4.12) | 6.75 | 39.57 (±52.54) | 3.32 (±2.51) |
| Non-woody | 441 | 178.62 | 0.41 (±0.80) | 4.25 | 40.17 (±47.60) | 3.78 (±2.37) |
| HEIDI Estimated Values | IA | EA | FP | H(2) | p |
|---|---|---|---|---|---|
| Short-range dispersers | 0.95 (±0.43) | 1.26 (±0.66) | 1.43 (±0.71) | 13.89 | <0.001 |
| Medium-range dispersers | 1.28 (±0.60) | 1.44 (±0.78) | 1.63 (±0.75) | 6.98 | 0.031 |
| Long-range dispersers | 1.25 (±0.81) | 1.61 (±1.33) | 1.54 (±1.02) | 5.51 | 0.064 |
| Global | 1.16 (±0.45) | 1.43 (±0.79) | 1.53 (±0.69) | 9.41 | 0.009 |
| HEIDI Estimated Values | Woody REIs | Non-Woody REIs | W | p |
|---|---|---|---|---|
| Short-range dispersers | 1.40 (±0.60) | 0.86 (±0.52) | 1097.50 | <0.001 |
| Medium-range dispersers | 1.46 (±0.67) | 1.37 (±0.77) | 2368.00 | 0.167 |
| Long-range dispersers | 1.72 (±1.25) | 1.03 (±0.40) | 1266.00 | <0.001 |
| Global | 1.53 (±0.68) | 1.09 (±0.51) | 1372.00 | <0.001 |
| HEIDI Categories and Metrics | IA | EA | FP | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Low | Fair | High | Low | Fair | High | Low | Fair | High | |
| 1. Vegetation structure | |||||||||
| Native tree species | 16.39 | 77.05 | 6.56 | 17.39 | 69.57 | 13.04 | 13.64 | 86.36 | 0.00 |
| Invasive species cover | 9.84 | 22.95 | 67.21 | 2.17 | 8.70 | 89.13 | 13.64 | 27.27 | 59.09 |
| Vertical strata | 8.20 | 57.38 | 34.43 | 10.87 | 69.57 | 19.57 | 9.09 | 68.18 | 22.73 |
| 2. Vegetation habitats | |||||||||
| Trees with microhabitats above 3 m | 77.05 | 13.11 | 9.84 | 67.39 | 13.04 | 19.57 | 56.82 | 18.18 | 25.00 |
| Trees with microhabitats below 3 m | 34.43 | 4.92 | 60.66 | 32.61 | 10.87 | 56.52 | 40.91 | 11.36 | 47.73 |
| Standing dead trees | 86.89 | 11.48 | 1.64 | 78.26 | 17.39 | 4.35 | 77.27 | 20.45 | 2.27 |
| Dead wood trunks on the ground | 44.26 | 18.03 | 37.70 | 34.78 | 21.74 | 43.48 | 34.09 | 31.82 | 34.09 |
| Large living trees | 60.66 | 27.87 | 11.48 | 78.26 | 21.74 | 0.00 | 79.55 | 6.82 | 13.64 |
| Leaf litter cover (short-range) | 9.84 | 63.93 | 26.23 | 8.70 | 60.87 | 30.43 | 6.82 | 63.64 | 29.55 |
| Leaf litter cover (medium-range) | 26.23 | 63.93 | 9.84 | 30.43 | 60.87 | 8.70 | 29.55 | 63.64 | 6.82 |
| 3. Associated habitats | |||||||||
| Rocky habitat types | 91.80 | 8.20 | 0.00 | 100.00 | 0.00 | 0.00 | 90.91 | 6.82 | 2.27 |
| Aquatic habitat types | 4.92 | 83.61 | 11.48 | 6.52 | 89.13 | 4.35 | 20.45 | 72.73 | 6.82 |
| 4. Vegetation management | |||||||||
| Understory clearing | 9.84 | 18.03 | 72.13 | 13.04 | 19.57 | 67.39 | 0.00 | 13.64 | 86.36 |
| Tree clearing | 1.64 | 6.56 | 91.80 | 6.52 | 2.17 | 91.30 | 0.00 | 4.55 | 95.45 |
| 5. Floristic suitability | |||||||||
| Seed production suitability | 44.26 | 54.10 | 1.64 | 10.87 | 60.87 | 28.26 | 6.82 | 52.27 | 40.91 |
| Pollen production suitability | 34.43 | 54.10 | 11.48 | 19.57 | 60.87 | 19.57 | 9.09 | 47.73 | 43.18 |
| Fruit production suitability | 36.07 | 55.74 | 8.20 | 10.87 | 56.52 | 32.61 | 4.55 | 63.64 | 31.82 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fonseca, A.; Zina, V.; Duarte, G.; Aguiar, F.C.; Rodríguez-González, P.M.; Ferreira, M.T.; Fernandes, M.R. Riparian Ecological Infrastructures: Potential for Biodiversity-Related Ecosystem Services in Mediterranean Human-Dominated Landscapes. Sustainability 2021, 13, 10508. https://doi.org/10.3390/su131910508
Fonseca A, Zina V, Duarte G, Aguiar FC, Rodríguez-González PM, Ferreira MT, Fernandes MR. Riparian Ecological Infrastructures: Potential for Biodiversity-Related Ecosystem Services in Mediterranean Human-Dominated Landscapes. Sustainability. 2021; 13(19):10508. https://doi.org/10.3390/su131910508
Chicago/Turabian StyleFonseca, André, Vera Zina, Gonçalo Duarte, Francisca C. Aguiar, Patricia María Rodríguez-González, Maria Teresa Ferreira, and Maria Rosário Fernandes. 2021. "Riparian Ecological Infrastructures: Potential for Biodiversity-Related Ecosystem Services in Mediterranean Human-Dominated Landscapes" Sustainability 13, no. 19: 10508. https://doi.org/10.3390/su131910508
APA StyleFonseca, A., Zina, V., Duarte, G., Aguiar, F. C., Rodríguez-González, P. M., Ferreira, M. T., & Fernandes, M. R. (2021). Riparian Ecological Infrastructures: Potential for Biodiversity-Related Ecosystem Services in Mediterranean Human-Dominated Landscapes. Sustainability, 13(19), 10508. https://doi.org/10.3390/su131910508