4.1. DM Yields
Broad experimental research has been conducted in the past to analyse growth dynamics of grassland species [
22,
32,
33,
34], and a wide variety of studies have examined the effect of specific defoliation frequencies or varying regrowth intervals [
35,
36,
37,
38]. Focusing on simple grass clover mixtures or different grass species, the studies have consistently found that a shortening of regrowth intervals leads to lower herbage yields, especially at the start of the vegetation period, whereas extended regrowth intervals of up to eight weeks increase total annual DM yields.
The results of the present study confirm these observations for a multi-species sward as well. Irrespective of the seed mixture, the total herbage yield decreased significantly with an increase in defoliation frequency. The yields of RC and the group of forage herbs were affected most. A comparison of the proportions that the different species contributed to the total DM yield showed that the highest proportion from RC occurred in the 6W system, whereas the highest proportions from all the other species occurred with more frequent defoliation. Nevertheless, in the 3W and 4W treatments too, RC was the highest-yielding species despite its low tolerance of frequent defoliation. According to [
39], a decrease in yield from RC should be expected after the first year of cultivation, making RC more suitable as a mixture component in short-term leys rather than in permanent pastures. Belesky et al. [
40] found a 26% higher herbage yield at longer regrowth intervals when comparing three-weekly and six-weekly defoliation of pure chicory stands, which is consistent with the effect of defoliation frequency on the group of forage herbs in the present experiment, with chicory as the dominating species. The DM yields of PR did not differ significantly between defoliation frequencies, in contrast to the results of [
41] who found an increase in DM yields with a decrease in defoliation frequency when analysing pure PR stands. In the current experiment it was assumed that the strong growth of RC reduced the production potential of PR under infrequent defoliation. The DM yield of white clover was not affected by defoliation frequency, thus confirming its high suitability for pasturing.
The compilation of the seed mixtures aimed to achieve appropriate sowing rates for all species in both mixtures while maintaining a similar ratio of leguminous to non-leguminous species. Hence the sowing rates of PR, RC, and WC were 50% lower in the complex mixture than in the simple mixture. The comparison of DM yields between the mixtures showed that the difference in sowing rate affected the three species to different extents. In the complex mixture, the yields of RC and PR were significantly lower, but represented more than 50% of their yields in the simple mixture. This indicates that in the simple mixture the growth of these species was restricted by inter-species or intra-species competition, which appeared to be lower in the complex mixture. The DM yields of the forage herbs and BT totalled just 18 to 21% of the total DM yield of the complex mixture. The DM production of WC did not differ between seed mixtures; the stoloniferous growth pattern allowed high dispersion even at the lower sowing rate. The growth curves of WC show that in the complex mixture a slower DM increment at the start of the season was compensated by maintaining high growth rates later in the season, when maximum growth of RC was limited by too few plants due to the lower sowing rate.
Very low DM production was observed for BT despite the beneficial conditions for legumes in the experimental design. The experiment was included in the five-year crop rotation on the experimental farm and each experimental year represented the first year of a two-year ley, which was established by undersowing a winter cereal in the May of the previous year. Due to the farm being managed in line with organic farming principles, no mineral N fertiliser was applied, and the N supply of the leys relied mainly on the symbiotic nitrogen fixation (SNF) of the legumes. This means that in each of the experimental years, the soils were N depleted and the legumes had a competitive advantage over the non-leguminous species. Plantain and chicory were the key species for determining the DM yield of the group of forage herbs, whereas caraway and burnet persisted only at a low occurrence in the swards of the complex mixture. Like RC, chicory had a competitive advantage through its deep taproot, which allows it to reach and utilise nutrients in soil layers below the rooting zone of the shallow-rooting species PR and WC [
42].
Sanderson et al. [
43] found that overyielding of low-diversity treatments is reasonable when they contain few species, but ones that are well adapted to the given environment. This applies to the current experiment, except that both seed mixtures were composed to ensure high herbage production by including highly productive species and provided similar DM yields. In particular, RC has been shown to increase the DM yields of grass clover swards [
39]. Despite the inclusion of less productive species in M
2, DM yields were similar for both mixtures. Due to the specific seed mixture composition and the fact that none of the species were cultivated in pure stands, the experiment was not designed to allow quantification of a diversity effect. Thus, it is unclear whether the average monoculture yield would have been higher or lower than the observed DM yields. Considering the growth limitations for the non-leguminous species (low soil N contents and the absence of fertilisation during the experimental periods), it can be assumed that the average monoculture yield would have been lower due to anticipated lower yields of the non-leguminous species when grown in pure stands. SNF and adjacent transfer of fixed N
2 from the legumes to the non-leguminous species, as described by [
44], was enabled in both mixtures. Conversely, some species with a low occurrence in the mixed sward might have developed better in pure stands due to less shading from other plant species. Further complementarity between species, irrespective of their ability to fix atmospheric N, as found by [
45,
46] in grass clover mixtures and by [
47,
48] in mixtures without legumes, might have occurred. Since the increase in species diversity did not increase DM yields, in contrast to the findings of previous studies [
49,
50], it is likely that the species that were abundant in M
1 offered functional traits being crucial for herbage production at the experimental site and that the species added in M
2 did not enhance positive complementary effects and are thus characterized by trait redundancy.
An explanation for the lack of overyielding from the simple mixture might be that with the admixture of species, the replacement of species with similar functional traits outweighed a potential complementation of functional traits. The group of forage herbs partially replaced PR as a non-leguminous species. In terms of growth morphology, chicory with its deep taproot and erect growth habit offered similar traits to RC. Simultaneously, BT offered the ability of SNF but was only able to replace RC in this trait to a negligible extent due to its low competitiveness for light.
The similar DM yields of both mixtures indicated that the number of species within a mixture was of minor importance and supports the findings of [
51] that functional group composition is more important in productive grasslands than species diversity. Similarly, [
23], who retrospectively analysed multiple biodiversity experiments, and [
52], who compared a wide variety of simple and complex mixtures, conclude that the composition of the mixture might be more important than its complexity.
4.2. Herbage Quality
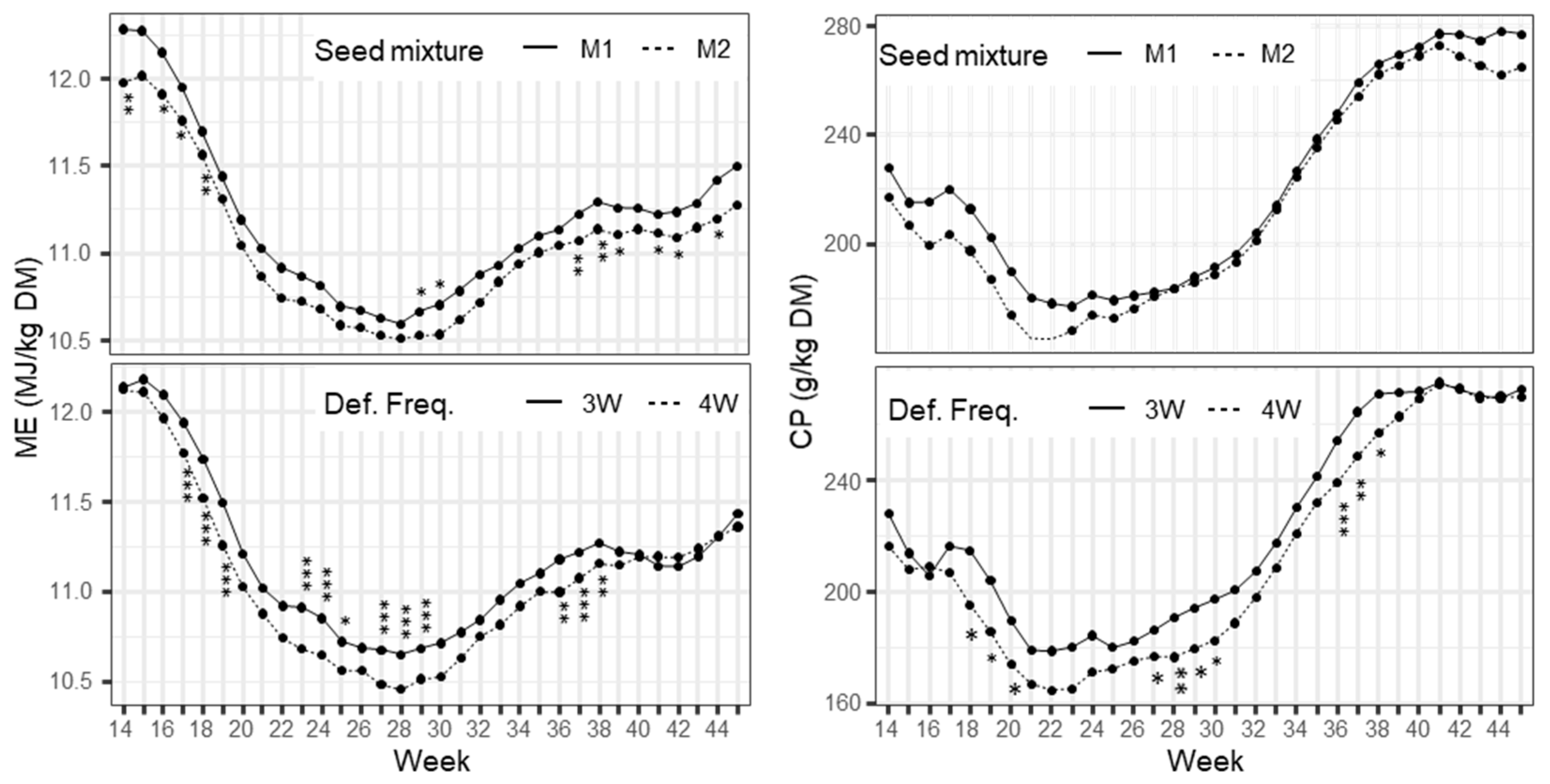
The results showed that the seed mixture had a significant effect on the ME content of the total herbage, but not on the ME content of individual species. This indicates similar sward densities in both mixtures, not affecting growth morphology of individual species. The low ME content of the group of herbs, in particular narrow-leafed plantain and chicory, caused the difference in the ME content of the total herbage. The CP content of PR was significantly lower in the complex mixture, which indicates a lower N supply to PR. Reasons for this could have been a smaller transfer of N from legumes due to their lower occurrence or higher competition for the available soil N due to the concomitance of other non-leguminous species.
According to the model developed by [
53], SNF averaged 40.51 g/m
2 in the simple mixture and 29.32 g/m
2 in the complex mixture. The respective N yields of the total herbage were 31.83 and 31.29 g/m
2. This might be an indication that higher competition for symbiotically fixed N in M
2 due to the lower legume proportion associated with lower SNF contributed to the lower CP content of PR in M
2. Additionally, in M
2 non-leguminous species other than PR competed for the plant-available N.
However, it should be noted that the model was not explicitly designed for mixed pasture swards containing non-leguminous plants other than PR. The highest yielding species in the group of forage herbs was chicory, which potentially reaches N deposited in deeper soil layers than PR [
54], but due to differences in the root morphology receives less N from legumes [
44], which makes a higher transfer of fixed N
2 to PR than to chicory likely. Høgh-Jensen et al. [
54] found a higher exploitation of soil N by chicory, which leads to higher accumulation of herbage N compared to PR. Their results from experiments using the
15N plant-labelling technique indicated that chicory is not a good competitor for N in upper soil layers, but can access N sources in deeper soil layers that are less accessible for other species. This suggests that chicory did not solely rely on the same N sources as PR, and contributed to the total N yield by a higher accumulation of N from deeper soil layers.
As grazing dairy cows are often exposed to excess N [
2] and as milk production might be limited not by a lack of CP but rather by a surplus of CP in relation to the ME supply of a pasture [
55], a lower CP content, as found in the complex mixture in the present experiment, might be more of an advantage than a drawback. For example, Totty et al. [
2] showed that grazing a diverse pasture, including chicory, plantain, and big trefoil besides high-sugar PR and WC, is beneficial for animal metabolism and the environment as it avoids the large urinary N losses that can occur when simple grass clover swards are grazed. However, in this regard it should be noted that since the present experiment resembled a rotational grazing system only in terms of high defoliation frequencies, no account was taken of pasture-specific effects such as nutrient return or trampling and, in terms of nutritional aspects, to the selective foraging behaviour of animals that influences the botanical and chemical composition of the actual intake.
Consideration of the total CP yields, which were similar for both mixtures, showed that the differences observed in the concentrations did not have an impact on yields due to a slightly higher DM production (n.s.) of M
2. This means that despite the lower yield of legumes, a similar amount of N was accumulated in the herbage overall. As described above, the deep-rooting trait of chicory might have contributed to the N supply of the mixed sward. Furthermore, Nyfeler et al. [
56] found that grasses stimulated the amount of symbiotically fixed N, with the effect being greatest at legume proportions of 40–60%. Although in the present experiment the yield of PR was not higher in M
2 than in M
1, the yield of all non-leguminous species together was higher, which might have induced a similar effect.
Unlike morphological traits such as deep rooting, the ability to increase the N content of the soils via SNF is offered by leguminous species only. Irrespective of defoliation frequency, the proportion of legumes at 75.5% of DM was significantly higher in M
1 than in M
2 at 61.9% of DM. Thus, the admixture of BT (as the only additional leguminous species) did not fully compensate for the lower SNF by RC in M
2, which resulted in a lower CP content of the total herbage. These results coincide with the findings of [
52] that the proportion of legumes, rather than species diversity, determines the CP content of the herbage.
The feeding quality of the herbage in terms of ME content and CP content was negatively affected by extending the regrowth interval. As expected, this effect was consistent across all species since the ongoing maturation at long regrowth intervals leads to an increase in the proportion of stem and a decreasing digestibility in all plant parts [
57]. The effect of defoliation frequency on the average annual ME yield was masked by the difference in DM yields. Despite the decrease in ME content at the extension of the regrowth interval, the yield of ME was significantly higher in the 6W system than in the simulated grazing treatments. The average annual CP yields, however, were significantly higher in the simulated grazing treatments than in the 6W system. This indicates that the effect of defoliation frequency on the CP content was, in contrast to the effect on the ME content, too great to be masked by the DM yield differences. These results show that despite differences in the ME content and CP content, both mixtures achieved similar annual DM, ME, and CP yields. The effect of defoliation frequency on annual ME and CP yields, however, was adverse. Whereas higher CP yields occurred at high defoliation frequencies, the highest ME yields occurred at 6W defoliation.
4.3. Yield Stability
The coefficient of variation is a well-established metric to describe the temporal stability of yields [
23,
51]. Nevertheless, various authors point out that, depending on the research question being addressed, CV is not necessarily the best parameter for describing yield stability [
42,
58]. Carnus et al. [
58] showed that CV can be misleading under certain circumstances, e.g., when low levels of the functional response are desired, or the level of the functional response is not relevant at all. In the present study, the functional responses were the yields of DM, ME, and CP for inter-annual CV and weekly growth rates for intra-annual CV. Thus, the desired state of the functional response is a high mean at a low standard deviation. Therefore, CV was considered to be a meaningful parameter when evaluated in combination with the respective mean and SD.
The seasonal distribution of the variability of forage growth is relevant for both grazing and zero-grazing systems. Late summer months are particularly prone to forage shortages. The widest variation in growth rate occurs during the generative phase in spring, which is favourable because it means a rapid increase in growth rate which can be managed by conservation for winter-feed production. The opposite situation, however, occurs in summer where there is a greater risk of shortages due to lower vegetative growth and higher risks of irregular precipitation. If there are no trade-offs in terms of lower average growth rates, an even yield distribution is desired in grazing systems. A high yield stability between years is important for planning the annual feed budget and is achieved when the mixture shows a low yield response to variations in the weather conditions. This is reflected in a low inter-annual CV. In the present experiment, the inter-annual CV was not affected by the seed mixture, which is in agreement with the results of [
23] who analysed herbage yield variability of mixtures of grasses and legumes from three experiments and found no consistent relationship between the number of species and the inter-annual CV.
The results from [
42] showed that a higher species richness can potentially increase yield stability during drought events, and several studies have found a higher yield potential during warm and dry periods for mixtures containing chicory or plantain [
13,
59]. During drought conditions, species increase the nutrient utilisation from deeper soil layers, which suits deep-rooting species that can access these layers more easily [
14]. However, due to the reduced yields of RC in M
2, the difference between the seed mixtures’ proportion of deep-rooting species was small in the present study and the absence of major weather events, such as drought, within the experimental years did not allow conclusions to be drawn on the different drought tolerance of the seed mixtures.
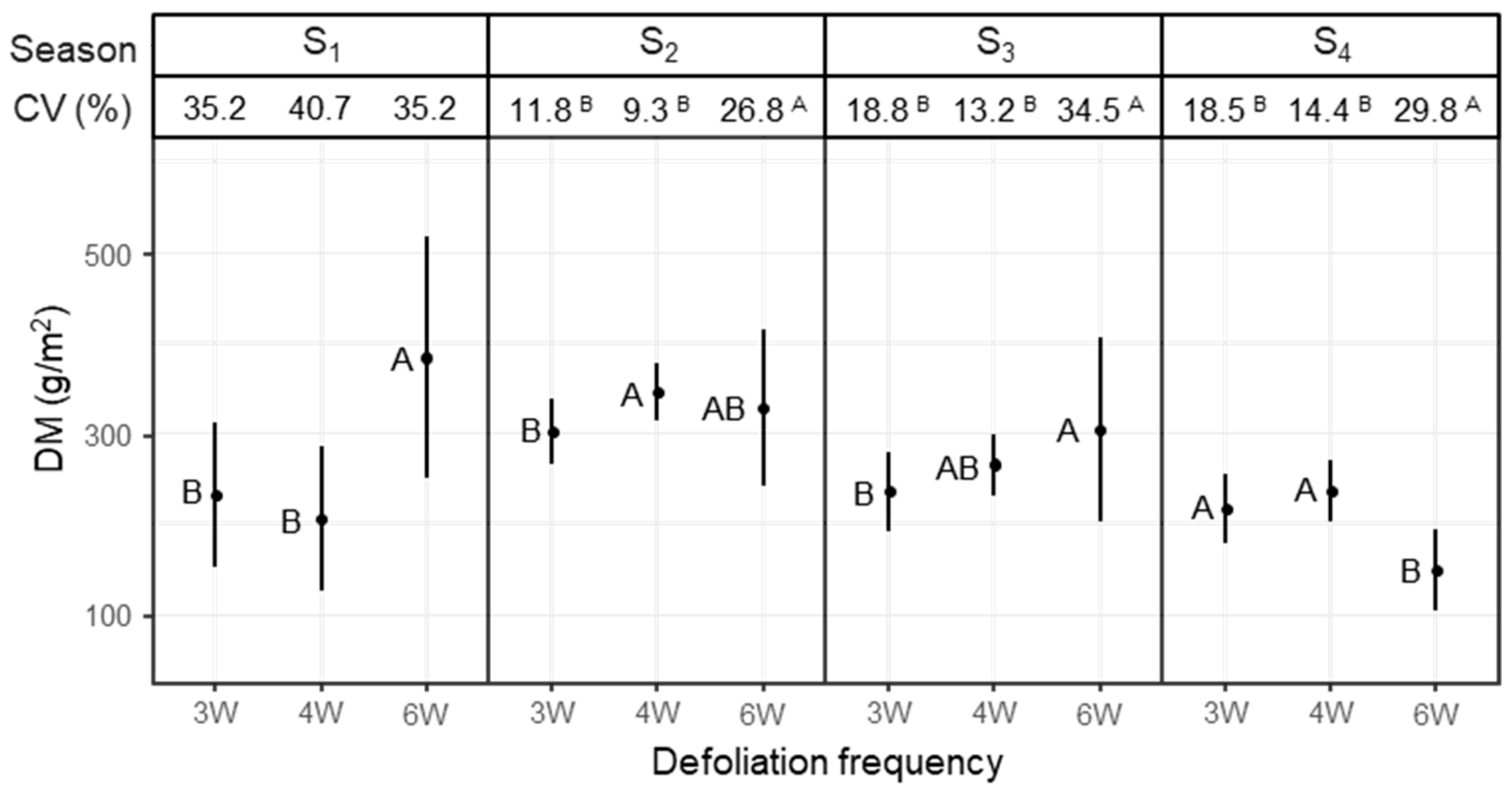
The seasonal fragmentation of the growing period was firstly based on the distribution of the harvests in the cutting-for-conservation system, but secondly was separated between different growth stages of PR. Whereas the reproductive growth phase was covered by season S1, the transition to vegetative growth occurred in S2, and the vegetative growth phase was split into summer and autumn growth in S3 and S4. The inter-annual CV of seasonal DM yields was higher in the 6W system than in the 3W and 4W systems in seasons S2 to S4, thus between the end of May and mid-November. In these seasons the average DM yields in the 3W and 4W systems were high and the SDs were small, which led to particularly low CVs. In the 6W system, the highest DM yield in season S1 co-occurred with a large SD, which eventually led to a large CV.
These results indicate that in all seasons except S1, a higher inter-annual yield stability was achieved by using high defoliation frequencies that were not tied to any trade-offs regarding DM production. In season S1, the SDs were also clearly lower in the simulated grazing systems, but so were the DM yields. Since in seasons S2 and S3 the inter-annual CVs of the 3W and 4W systems were a combination of high means and low SDs, which was considered favourable against the backdrop of this study, CV has a high informative value in these cases.
The inter-annual CV of the annual DM yields was higher with the frequent defoliation of the simulated grazing treatments despite the smaller DM yields. In the 6W system, the DM yields were high but so were the SDs, which means that variations between years and also the effects of weather events were greater under less frequent defoliation, which led to comparatively large CVs.
The analysis of the stability between weekly growth rates in the simulated grazing systems, described by the intra-annual CV, showed a significant effect of seed mixture, with CV being significantly higher in the simple mixture M
1. Due to similar annual DM yields, this indicates a more even distribution of growth rates in the complex mixture M
2. Since the greatest variation between growth rates occurred during the generative phase, when growth rates rapidly increase from very low to high, a separate analysis (not presented) was carried out for calendar weeks 25 to 47 only. The significant difference between the seed mixtures remained following the exclusion of the generative phase, which showed that the effect on intra-annual stability did not come from a lower peak in spring, but from a more even distribution of growth rates throughout the growing season, primarily driven by good performance of WC in seasons S
2 and S
3 (end of May to middle of August, see
Figure 2). The reduction of RC seed density in M
2 in combination with stoloniferous growth of WC supported strong competitiveness of WC during summer months. Additionally, in comparison with PR, the observed growth rates of the group of forage herbs showed a less distinct peak production in spring and a second peak during the growth depression in PR after the conversion to vegetative growth. This indicates that in terms of seasonal growth distribution, the forage herbs did not replace PR, but with their divergent growth pattern contributed to a more even distribution of herbage growth.








{kind=link}
{kind=link}
{kind=link}
{kind=link}