From Nucleotides to Satellite Imagery: Approaches to Identify and Manage the Invasive Pathogen Xylella fastidiosa and Its Insect Vectors in Europe

 , ,
, ,  ,
,  and
and
Abstract
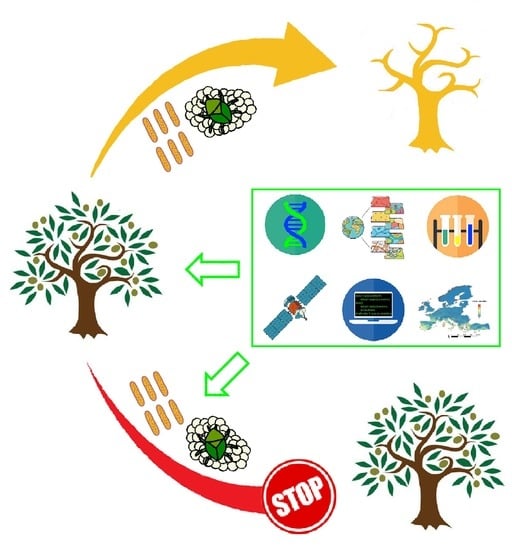
1. Introduction
2. Genetic Approaches
2.1. Sequencing Data
| Term | Definition |
| Accessory genome | Part of genes that are not present in all strains of a species; typically includes strain specific adaptation such as antibiotic resistance. |
| Clade | A group of organisms that include a common ancestor and all its descendants. |
| Competence | Ability of a microorganism to incorporate exogenous DNA in its genome. |
| Core genome | Part of genes that are present in all strains of a species; typically includes housekeeping genes for cell envelope or regulatory functions. |
| Genetic vector | Vehicles (e.g., DNA plasmids, viruses, artificial chromosomes) for delivering foreign DNA into recipient cells. |
| Genome assembly | A computational representation of a genome sequence. Genomes are not sequenced along the complete length of a chromosome but fragmented: these fragments are sequenced and then put back together (assembled). |
| Epigenetic variation | A hereditary but reversible change in gene expression not involving modifications in the DNA sequence. |
| Isolate | A sample or culture of microorganisms isolated for study. |
| Multi-locus sequence typing (MLST) | A standardized typing method based on Sanger sequencing of seven housekeeping genes not under positive selection; each allelic combination defines a Sequence Type (ST). |
| Pangenome | The entire gene set of all strains of a species, including both the core and accessory genome. |
| Pathogen confusion | Alteration of the in planta levels of diffusible signaling factors that regulate pathogen virulence to reduce infections. |
| Plasmid | A small, extrachromosomal DNA molecule within a cell that is physically separated from chromosomal DNA and can replicate independently. |
| Prophage | A bacteriophage genome that is inserted and integrated into the circular bacterial DNA chromosome or exists as an extrachromosomal plasmid. |
| Repeated genomic regions | Sequence patterns that occur in multiple copies throughout the genome. |
| Sequence type (ST) | A group of individuals sharing the same multi-locus sequence typing (MLST) profile. |
| Strain | A group of microorganisms that are genetically distinct from other groups of the same species or subspecies. |
2.2. Genetic Diversity
2.3. Diagnostic Protocols
2.4. Gene Expression Profiles
2.5. Genetic Engineering
3. Spatial Ecology Approaches
3.1. Remote Sensing
| Term | Definition |
| Area Under Curve (AUC) | A measurement of the discriminatory ability of classification models. The closer the AUC to 1, the better the predictive ability of the model. |
| Bayesian inference approach | A method of statistical inference in which the Bayes’ theorem is used to update the probability for a hypothesis as more evidence or information becomes available. |
| biomod2 | An R package for ensemble forecasting of species distributions, enabling the treatment of a range of methodological uncertainties in models and the examination of species-environment associations. |
| Ensemble modeling | It is a process where multiple diverse models are created to predict an outcome, either by using many different modeling algorithms or using different training datasets. |
| Hyperspectral image | Differs from the multispectral image (Table S3) because it consists of hundreds of spectral bands of limited width. |
| Leaf Area Index (LAI) | Total area of green elements (one side) in the canopy per unit horizontal ground area (m2/m2). |
| Leaf chlorophyll content | The amount of chlorophyll per unit leaf area. |
| Leaf water content | The amount of water present in a leaf. |
| Maxent | A software to model species niches and distributions by applying a machine-learning technique called maximum entropy modeling. |
| Multispectral image | A stack of several digital images, each one corresponding to the radiation intensity measured in a specific wavelength interval (spectral band, Table S3), which has been reflected or emitted by ground surface elements, represented by the image pixels. |
| Normalized Difference Vegetation Index (NDVI) | Commonly used index calculated as (NIR-RED)/(NIR+RED), with NIR representing the reflectance measured at the near-infrared wavelength (750–950 nm, strongly reflected by vegetation, Table S3) and RED the corresponding at the red wavelength (620–700 nm, absorbed by vegetation, Table S3). |
| Phenocam | A digital camera set up at a fixed location to capture time-lapse images. |
| Solar induced chlorophyll fluorescence (SIF) | The reemitted solar light in the 650–850 nm range (Table S3) from the chlorophyll-a pigment, which is linked to the initial steps in photosynthesis. |
| Spectral reflectance | Generic term to express the reflecting ability of a surface at a given wavelength of the electromagnetic spectrum. The distribution of spectral reflectance along the spectrum is called “reflectance curve” which has a characteristic shape for different kinds of surfaces (i.e., water, soil, vegetation, etc.). |
| Thermal image | Digital image representing the amount of radiation emitted by a surface in the thermal infrared range (7000–13,000 nm, Table S3), which is related with the temperature of the surface itself and other characteristics (i.e., emissivity). |
| True Skill Statistics (TSS) | It compares the number of correct forecasts minus those attributable to random guessing to that of a hypothetical set of ideal forecasts. It considers both omission and commission errors and success as a result of random guessing; its values range from −1 to +1, where +1 corresponds to perfect agreement and zero or less to a performance no better than random. |
| Vegetation index | It is a spectral transformation of two or more bands designed to enhance the contribution of vegetation properties and allow reliable spatial and temporal inter-comparisons of terrestrial photosynthetic activity and canopy structural variations. |
3.2. Geographic Information Systems
3.3. Spatially Explicit Models
4. Research Perspectives
4.1. The Impact of Climate Change
4.2. Genetic Approaches
4.3. Spatial Ecology
5. Integrating Genetic and Ecological Approaches
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Williamson, M.; Griffiths, B. Biological Invasions; Springer Science & Business Media: London, UK, 1996; ISBN 0-412-59190-1. [Google Scholar]
- Levine, J.M. Biological invasions. Curr. Biol. 2008, 18, R57–R60. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, B.D.; Broadhurst, L.M. The economic cost of managing invasive species in Australia. NeoBiota 2016, 31, 1. [Google Scholar] [CrossRef]
- Hulme, P.E.; Pyšek, P.; Nentwig, W.; Vilà, M. Will threat of biological invasions unite the European Union? Science 2009, 324, 40–41. [Google Scholar] [CrossRef] [PubMed]
- Charles, H.; Dukes, J.S. Impacts of Invasive Species on Ecosystem Services. In Biological Invasions; Nentwig, W., Ed.; Ecological Studies; Springer: Berlin/Heidelberg, Germany, 2008; pp. 217–237. ISBN 978-3-540-36920-2. [Google Scholar]
- Bertolino, S.; Sciandra, C.; Bosso, L.; Russo, D.; Lurz, P.W.; Di Febbraro, M. Spatially explicit models as tools for implementing effective management strategies for invasive alien mammals. Mammal Rev. 2020. [Google Scholar] [CrossRef]
- Thakur, M.P.; van der Putten, W.H.; Cobben, M.M.; van Kleunen, M.; Geisen, S. Microbial invasions in terrestrial ecosystems. Nat. Rev. Microbiol. 2019, 17, 621–631. [Google Scholar] [CrossRef]
- Van der Putten, W.H.; Klironomos, J.N.; Wardle, D.A. Microbial ecology of biological invasions. ISME J. 2007, 1, 28–37. [Google Scholar] [CrossRef]
- Litchman, E. Invisible invaders: Non-pathogenic invasive microbes in aquatic and terrestrial ecosystems. Ecol. Lett. 2010, 13, 1560–1572. [Google Scholar] [CrossRef]
- Peeri, N.C.; Shrestha, N.; Rahman, M.S.; Zaki, R.; Tan, Z.; Bibi, S.; Baghbanzadeh, M.; Aghamohammadi, N.; Zhang, W.; Haque, U. The SARS, MERS and novel coronavirus (COVID-19) epidemics, the newest and biggest global health threats: What lessons have we learned? Int. J. Epidemiol. 2020. [Google Scholar] [CrossRef]
- Ghelardini, L.; Luchi, N.; Pecori, F.; Pepori, A.L.; Danti, R.; Della Rocca, G.; Capretti, P.; Tsopelas, P.; Santini, A. Ecology of invasive forest pathogens. Biol. Invasions 2017, 19, 3183–3200. [Google Scholar] [CrossRef]
- Wells, J.M.; Raju, B.C.; Hung, H.-Y.; Weisburg, W.G.; Mandelco-Paul, L.; Brenner, D.J. Xylella fastidiosa gen. nov., sp. nov: Gram-Negative, Xylem-Limited, Fastidious Plant Bacteria Related to Xanthomonas spp. Int. J. Syst. Evol. Microbiol. 1987, 37, 136–143. [Google Scholar] [CrossRef]
- Bragard, C.; Dehnen-Schmutz, K.; Di Serio, F.; Gonthier, P.; Jacques, M.; Miret, J.A.J.; Justesen, A.F.; MacLeod, A.; Magnusson, C.S.; Milonas, P. Update of the Scientific Opinion on the risks to plant health posed by Xylella fastidiosa in the EU territory. EFSA J. 2019, 17. [Google Scholar] [CrossRef]
- Schneider, K.; van der Werf, W.; Cendoya, M.; Mourits, M.; Navas-Cortés, J.A.; Vicent, A.; Oude Lansink, A. Impact of Xylella fastidiosa subspecies pauca in European olives. Proc. Natl. Acad. Sci. USA 2020, 201912206. [Google Scholar] [CrossRef] [PubMed]
- European and Mediterranean Plant Protection Organization. EPPO PM 7/24 (4) Xylella fastidiosa. EPPO Bull. 2019, 49, 175–227. [Google Scholar] [CrossRef]
- Janse, J.; Obradovic, A. Xylella fastidiosa: Its biology, diagnosis, control and risks. J. Plant Pathol. 2010, S35–S48. [Google Scholar] [CrossRef]
- Chatterjee, S.; Almeida, R.P.P.; Lindow, S. Living in two worlds: The plant and insect lifestyles of Xylella fastidiosa. Annu. Rev. Phytopathol. 2008, 46, 243–271. [Google Scholar] [CrossRef]
- Sabella, E.; Aprile, A.; Genga, A.; Siciliano, T.; Nutricati, E.; Nicolì, F.; Vergine, M.; Negro, C.; De Bellis, L.; Luvisi, A. Xylem cavitation susceptibility and refilling mechanisms in olive trees infected by Xylella fastidiosa. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Saponari, M.; Boscia, D.; Nigro, F.; Martelli, G. Identification of DNA sequences related to Xylella fastidiosa in oleander, almond and olive trees exhibiting leaf scorch symptoms in Apulia (Southern Italy). J. Plant Pathol. 2013, 95. [Google Scholar] [CrossRef]
- European and Mediterranean Plant Protection Organization. EPPO Annual report and Council recommendations 2017. EPPO Bull. 2018, 48, 609–619. [Google Scholar] [CrossRef]
- Landa, B.B.; Castillo, A.I.; Giampetruzzi, A.; Kahn, A.; Román-Écija, M.; Velasco-Amo, M.P.; Navas-Cortés, J.A.; Marco-Noales, E.; Barbé, S.; Moralejo, E. Emergence of a plant pathogen in Europe associated with multiple intercontinental introductions. Appl. Environ. Microbiol. 2020, 86. [Google Scholar] [CrossRef]
- Frazier, N.W. Xylem viruses and their insect vectors. In Proceedings of the Proceedings International Conference on Virus and Vectors on Perennial Hosts, with Special Reference to Vitis, Davis, CA, USA, 6–10 September 1965. [Google Scholar]
- Almeida, R.P.P.; Blua, M.J.; Lopes, J.R.S.; Purcell, A.H. Vector Transmission of Xylella fastidiosa: Applying Fundamental Knowledge to Generate Disease Management Strategies. Ann. Entomol. Soc. Am. 2005, 98, 775–786. [Google Scholar] [CrossRef]
- Redak, R.A.; Purcell, A.H.; Lopes, J.R.S.; Blua, M.J.; Mizell, R.F., III; Andersen, P.C. The Biology of Xylem Fluid–Feeding Insect Vectors of Xylella Fastidiosa and Their Relation to Disease Epidemiology. Annu. Rev. Entomol. 2004, 49, 243–270. [Google Scholar] [CrossRef] [PubMed]
- Cornara, D.; Marra, M.; Tedone, B.; Cavalieri, V.; Porcelli, F.; Fereres, A.; Purcell, A.; Saponari, M. No evidence for cicadas’ implication in Xylella fastidiosa epidemiology. Entomol. Gen. 2020, 125–132. [Google Scholar] [CrossRef]
- European Food Safety Authority Panel on Plant Health (PLH). EFSA Scientific Opinion on the risks to plant health posed by Xylella fastidiosa in the EU territory, with the identification and evaluation of risk reduction options. EFSA J. 2015, 13, 3989. [Google Scholar] [CrossRef]
- Severin, H. Spittle-insect vectors of Pierce’s disease virus: II. Life history and virus transmission. Hilgardia 1950, 19, 357–382. [Google Scholar] [CrossRef]
- Purcell, A.H. Almond Leaf Scorch: Leafhopper and Spittlebug Vectors. J. Econ. Entomol. 1980, 73, 834–838. [Google Scholar] [CrossRef]
- Sanderlin, R.S.; Melanson, R.A. Insect Transmission of Xylella fastidiosa to Pecan. Plant Dis. 2010, 94, 465–470. [Google Scholar] [CrossRef]
- Cornara, D.; Saponari, M.; Zeilinger, A.R.; de Stradis, A.; Boscia, D.; Loconsole, G.; Bosco, D.; Martelli, G.P.; Almeida, R.P.P.; Porcelli, F. Spittlebugs as vectors of Xylella fastidiosa in olive orchards in Italy. J. Pest Sci. 2017, 90, 521–530. [Google Scholar] [CrossRef]
- Jeger, M.; Caffier, D.; Candresse, T.; Chatzivassiliou, E.; Dehnen-Schmutz, K.; Gilioli, G.; Grégoire, J.-C.; Miret, J.A.J.; MacLeod, A.; Navarro, M.N.; et al. Updated pest categorisation of Xylella fastidiosa. EFSA J. 2018, 16, e05357. [Google Scholar] [CrossRef]
- Yurtsever, S. On the Polymorphic Meadow Spittlebug, Philaenus spumarius (L.) (Homoptera: Cercopidae). Turk. J. Zool. 2000, 24, 447–460. [Google Scholar]
- Cavalieri, V.; Altamura, G.; Fumarola, G.; di Carolo, M.; Saponari, M.; Cornara, D.; Bosco, D.; Dongiovanni, C. Transmission of Xylella fastidiosa Subspecies Pauca Sequence Type 53 by Different Insect Species. Insects 2019, 10, 324. [Google Scholar] [CrossRef]
- Cornara, D.; Morente, M.; Markheiser, A.; Bodino, N.; Tsai, C.-T.; Fereres, A.; Redak, R.A.; Perring, T.M.; Spotti Lopes, J.R. An overview on the worldwide vectors of Xylella fastidiosa. Entomol. Gen. 2019, 39, 157–181. [Google Scholar] [CrossRef]
- European and Mediterranean Plant Protection Organization. EPPO PM 7/141 (1) Philaenus spumarius, Philaenus italosignus and Neophilaenus campestris. EPPO Bull. 2020, epp.12610. [Google Scholar] [CrossRef]
- Elbeaino, T.; Yaseen, T.; Valentini, F.; Ben Moussa, I.E.; Mazzoni, V.; D’onghia, A.M. Identification of three potential insect vectors of Xylella fastidiosa in southern Italy. Phytopathol. Mediterr. 2014, 53, 328–332. [Google Scholar] [CrossRef]
- EFSA Panel on Plant Health (EFSA PLH Panel); Bragard, C.; Dehnen-Schmutz, K.; Di Serio, F.; Gonthier, P.; Jacques, M.-A.; Jaques Miret, J.A.; Justesen, A.F.; MacLeod, A.; Magnusson, C.S.; et al. Effectiveness of in planta control measures for Xylella fastidiosa. EFSA J. 2019, 17, e05666. [Google Scholar] [CrossRef]
- Liccardo, A.; Fierro, A.; Garganese, F.; Picciotti, U.; Porcelli, F. A biological control model to manage the vector and the infection of Xylella fastidiosa on olive trees. PLoS ONE 2020, 15, e0232363. [Google Scholar] [CrossRef]
- Catalano, L.; Shoki, A.-D.; Boscia, D.; Martelli, G.P. Guidelines for the Prevention, Eradication and Containment of Xylella Fastidiosa in Olive-Growing Areas; Food and Agriculture Organization of the United Nations: Cairo, Egypt, 2019; p. 64. [Google Scholar]
- Lawson Handley, L.-J.; Estoup, A.; Evans, D.M.; Thomas, C.E.; Lombaert, E.; Facon, B.; Aebi, A.; Roy, H.E. Ecological genetics of invasive alien species. BioControl 2011, 56, 409–428. [Google Scholar] [CrossRef]
- Bouyer, J.; Lancelot, R. Using genetic data to improve species distribution models. Infect. Genet. Evol. 2018, 63, 292–294. [Google Scholar] [CrossRef]
- Yamasaki, E.; Altermatt, F.; Cavender-Bares, J.; Schuman, M.C.; Zuppinger-Dingley, D.; Garonna, I.; Schneider, F.D.; Guillén-Escribà, C.; van Moorsel, S.J.; Hahl, T.; et al. Genomics meets remote sensing in global change studies: Monitoring and predicting phenology, evolution and biodiversity. Curr. Opin. Environ. Sustain. 2017, 29, 177–186. [Google Scholar] [CrossRef]
- Burger, M. Applying GIS and Remote Sensing to Landscape Genetics and Genome Size Research. Available online: https://pdfs.semanticscholar.org/3054/4c8c85b5bfc7e3f07725329376120473178a.pdf (accessed on 28 May 2020).
- Waldvogel, A.-M.; Feldmeyer, B.; Rolshausen, G.; Exposito-Alonso, M.; Rellstab, C.; Kofler, R.; Mock, T.; Schmid, K.; Schmitt, I.; Bataillon, T.; et al. Evolutionary genomics can improve prediction of species’ responses to climate change. Evol. Lett. 2020, 4, 4–18. [Google Scholar] [CrossRef]
- Searle, J.B. The genetics of mammalian invasions: A review. Wildl. Res. 2008, 35, 185–192. [Google Scholar] [CrossRef]
- Barrett, S.C.H. Foundations of invasion genetics: The Baker and Stebbins legacy. Mol. Ecol. 2015, 24, 1927–1941. [Google Scholar] [CrossRef]
- Welles, S.R.; Dlugosch, K.M. Population Genomics of Colonization and Invasion. In Population Genomics: Concepts, Approaches and Applications; Rajora, O.P., Ed.; Population Genomics; Springer International Publishing: Cham, Switzerland, 2019; pp. 655–683. ISBN 978-3-030-04589-0. [Google Scholar]
- Reznick, D.N.; Losos, J.; Travis, J. From low to high gear: There has been a paradigm shift in our understanding of evolution. Ecol. Lett. 2019, 22, 233–244. [Google Scholar] [CrossRef]
- Dupuis, J.R.; Ruiz-Arce, R.; Barr, N.B.; Thomas, D.B.; Geib, S.M. Range-wide population genomics of the Mexican fruit fly: Toward development of pathway analysis tools. Evol. Appl. 2019, 12, 1641–1660. [Google Scholar] [CrossRef]
- Makino, T.; Kawata, M. Invasive invertebrates associated with highly duplicated gene content. Mol. Ecol. 2019, 28, 1652–1663. [Google Scholar] [CrossRef]
- Hamelin, R.C.; Roe, A.D. Genomic biosurveillance of forest invasive alien enemies: A story written in code. Evol. Appl. 2020, 13, 95–115. [Google Scholar] [CrossRef]
- Hammond, A.; Galizi, R.; Kyrou, K.; Simoni, A.; Siniscalchi, C.; Katsanos, D.; Gribble, M.; Baker, D.; Marois, E.; Russell, S.; et al. A CRISPR-Cas9 gene drive system targeting female reproduction in the malaria mosquito vector Anopheles gambiae. Nat. Biotechnol. 2016, 34, 78–83. [Google Scholar] [CrossRef]
- Kyrkou, I.; Pusa, T.; Ellegaard-Jensen, L.; Sagot, M.-F.; Hansen, L.H. Pierce’s Disease of Grapevines: A Review of Control Strategies and an Outline of an Epidemiological Model. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, Y.; Zeng, B.; Wang, Y.; James, A.A.; Gurr, G.M.; Yang, G.; Lin, X.; Huang, Y.; You, M. CRISPR/Cas9 mediated knockout of the abdominal-A homeotic gene in the global pest, diamondback moth (Plutella xylostella). Insect Biochem. Mol. Biol. 2016, 75, 98–106. [Google Scholar] [CrossRef]
- Wu, N.; Zhang, S.; Li, X.; Cao, Y.; Liu, X.; Wang, Q.; Liu, Q.; Liu, H.; Hu, X.; Zhou, X.J.; et al. Fall webworm genomes yield insights into rapid adaptation of invasive species. Nat. Ecol. Evol. 2019, 3, 105–115. [Google Scholar] [CrossRef]
- Simpson, A.J.G.; Reinach, F.C.; Arruda, P.; Abreu, F.A.; Acencio, M.; Alvarenga, R.; Alves, L.M.C.; Araya, J.E.; Baia, G.S.; Baptista, C.S.; et al. The genome sequence of the plant pathogen Xylella fastidiosa. Nature 2000, 406, 151–157. [Google Scholar] [CrossRef]
- Retchless, A.C.; Labroussaa, F.; Shapiro, L.; Stenger, D.C.; Lindow, S.E.; Almeida, R.P.P. Genomic insights into Xylella fastidiosa interactions with plant and insect hosts. In Genomics of Plant-Associated Bacteria; Springer: Berlin/Heidelberg, Germany, 2014; pp. 177–202. ISBN 978-3-642-55378-3. [Google Scholar]
- Vanhove, M.; Retchless, A.C.; Sicard, A.; Rieux, A.; Coletta-Filho, H.D.; Fuente, L.D.L.; Stenger, D.C.; Almeida, R.P.P. Genomic Diversity and Recombination among Xylella fastidiosa Subspecies. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef]
- Potnis, N.; Kandel, P.P.; Merfa, M.V.; Retchless, A.C.; Parker, J.K.; Stenger, D.C.; Almeida, R.P.P.; Bergsma-Vlami, M.; Westenberg, M.; Cobine, P.A.; et al. Patterns of inter- and intrasubspecific homologous recombination inform eco-evolutionary dynamics of Xylella fastidiosa. ISME J. 2019, 13, 2319–2333. [Google Scholar] [CrossRef]
- Bhattacharyya, A.; Stilwagen, S.; Reznik, G.; Feil, H.; Feil, W.S.; Anderson, I.; Bernal, A.; D’Souza, M.; Ivanova, N.; Kapatral, V.; et al. Draft Sequencing and Comparative Genomics of Xylella fastidiosa Strains Reveal Novel Biological Insights. Genome Res. 2002, 12, 1556–1563. [Google Scholar] [CrossRef]
- Chen, J.; Xie, G.; Han, S.; Chertkov, O.; Sims, D.; Civerolo, E.L. Whole Genome Sequences of Two Xylella fastidiosa Strains (M12 and M23) Causing Almond Leaf Scorch Disease in California. J. Bacteriol. 2010, 192, 4534. [Google Scholar] [CrossRef]
- Giampetruzzi, A.; Chiumenti, M.; Saponari, M.; Donvito, G.; Italiano, A.; Loconsole, G.; Boscia, D.; Cariddi, C.; Martelli, G.P.; Saldarelli, P. Draft genome sequence of the Xylella fastidiosa CoDiRO strain. Genome Announc. 2015, 3, e01538-14. [Google Scholar] [CrossRef]
- Giampetruzzi, A.; D’Attoma, G.; Zicca, S.; Abou Kubaa, R.; Rizzo, D.; Boscia, D.; Saldarelli, P.; Saponari, M. Draft Genome Sequence Resources of Three Strains (TOS4, TOS5, and TOS14) of Xylella fastidiosa Infecting Different Host Plants in the Newly Discovered Outbreak in Tuscany, Italy. Phytopathology 2019, 109, 1516–1518. [Google Scholar] [CrossRef]
- Giampetruzzi, A.; Saponari, M.; Almeida, R.P.; Essakhi, S.; Boscia, D.; Loconsole, G.; Saldarelli, P. Complete genome sequence of the olive-infecting strain Xylella fastidiosa subsp. pauca De Donno. Genome Announc. 2017, 5, e00569-17. [Google Scholar] [CrossRef]
- Guan, W.; Shao, J.; Zhao, T.; Huang, Q. Genome sequence of a Xylella fastidiosa strain causing mulberry leaf scorch disease in Maryland. Genome Announc. 2014, 2, e00916-13. [Google Scholar] [CrossRef]
- Ramazzotti, M.; Cimaglia, F.; Gallo, A.; Ranaldi, F.; Surico, G.; Mita, G.; Bleve, G.; Marchi, G. Insights on a founder effect: The case of Xylella fastidiosa in the Salento area of Apulia, Italy. Phytopathol. Mediterr. 2018, 57, 8–25. [Google Scholar] [CrossRef]
- Schreiber, H.L.; Koirala, M.; Lara, A.; Ojeda, M.; Dowd, S.E.; Bextine, B.; Morano, L. Unraveling the first Xylella fastidiosa subsp. fastidiosa genome from Texas. Southwest. Entomol. 2010, 35, 479–484. [Google Scholar] [CrossRef]
- Besser, J.; Carleton, H.A.; Gerner-Smidt, P.; Lindsey, R.L.; Trees, E. Next-generation sequencing technologies and their application to the study and control of bacterial infections. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2018, 24, 335–341. [Google Scholar] [CrossRef]
- De Maio, N.; Shaw, L.P.; Hubbard, A.; George, S.; Sanderson, N.D.; Swann, J.; Wick, R.; AbuOun, M.; Stubberfield, E.; Hoosdally, S.J.; et al. Comparison of long-read sequencing technologies in the hybrid assembly of complex bacterial genomes. Microb. Genom. 2019, 5. [Google Scholar] [CrossRef]
- Richardson, E.J.; Watson, M. The automatic annotation of bacterial genomes. Brief. Bioinform. 2013, 14, 1–12. [Google Scholar] [CrossRef]
- Lemos de Macedo, E.G.; Alves, L.M.C.; Campanharo, J.C. Genomics-based design of defined growth media for the plant pathogen Xylella fastidiosa. FEMS Microbiol. Lett. 2003, 219, 39–45. [Google Scholar] [CrossRef]
- Biello, R.; Mathers, T.C.; Mugford, S.T.; Liu, Q.; Rodrigues, A.S.B.; Neto, A.C.; Rebelo, M.T.; Paulo, O.S.; Seabra, S.G.; Hogenhout, S.A. Draft Genome Assembly Version 1 of the Meadow Spittlebug Philaenus spumarius (Linnaeus, 1758) (Hemiptera, Aphrophoridae). Available online: https://zenodo.org/record/3368385#.Xs9ejMARXIU (accessed on 31 January 2020).
- Almeida, R.P.P.; Nunney, L. How Do Plant Diseases Caused by Xylella fastidiosa Emerge? Plant Dis. 2015, 99, 1457–1467. [Google Scholar] [CrossRef]
- Denancé, N.; Briand, M.; Gaborieau, R.; Gaillard, S.; Jacques, M.-A. Identification of genetic relationships and subspecies signatures in Xylella fastidiosa. BMC Genom. 2019, 20, 239. [Google Scholar] [CrossRef]
- Mang, S.M.; Frisullo, S.; Elshafie, H.S.; Camele, I. Diversity Evaluation of Xylella fastidiosa from Infected Olive Trees in Apulia (Southern Italy). Plant Pathol. J. 2016, 32, 102–111. [Google Scholar] [CrossRef]
- Marcelletti, S.; Scortichini, M. Genome-wide comparison and taxonomic relatedness of multiple Xylella fastidiosa strains reveal the occurrence of three subspecies and a new Xylella species. Arch. Microbiol. 2016, 198, 803–812. [Google Scholar] [CrossRef]
- Baldi, P.; La Porta, N. Xylella fastidiosa: Host Range and Advance in Molecular Identification Techniques. Front. Plant Sci. 2017, 8, 944. [Google Scholar] [CrossRef]
- Bergsma-Vlami, M.; van de Bilt, J.L.J.; Tjou-Tam-Sin, N.N.A.; Helderman, C.M.; Gorkink-Smits, P.; Landman, N.M.; van Nieuwburg, J.G.W.; van Veen, E.J.; Westenberg, M. Assessment of the genetic diversity of Xylella fastidiosa in imported ornamental Coffea arabica plants. Plant Pathol. 2017, 66, 1065–1074. [Google Scholar] [CrossRef]
- Bull, C.T.; Boer, S.H.D.; Denny, T.P.; Firrao, G.; Saux, M.F.-L.; Saddler, G.S.; Scortichini, M.; Stead, D.E.; Takikawa, Y. List of New Names of Plant Pathogenic Bacteria (2008–2010). J. Plant Pathol. 2012, 94, 21–27. [Google Scholar] [CrossRef]
- Denancé, N.; Legendre, B.; Briand, M.; Olivier, V.; de Boisseson, C.; Poliakoff, F.; Jacques, M.-A. Several subspecies and sequence types are associated with the emergence of Xylella fastidiosa in natural settings in France. Plant Pathol. 2017, 66, 1054–1064. [Google Scholar] [CrossRef]
- Nunney, L.; Azad, H.; Stouthamer, R. An Experimental Test of the Host-Plant Range of Nonrecombinant Strains of North American Xylella fastidiosa subsp. multiplex. Phytopathology 2019, 109, 294–300. [Google Scholar] [CrossRef]
- Nunney, L.; Ortiz, B.; Russell, S.A.; Sánchez, R.R.; Stouthamer, R. The complex biogeography of the plant pathogen Xylella fastidiosa: Genetic evidence of introductions and subspecific introgression in Central America. PLoS ONE 2014, 9, e112463. [Google Scholar] [CrossRef] [PubMed]
- Nunney, L.; Schuenzel, E.L.; Scally, M.; Bromley, R.E.; Stouthamer, R. Large-scale intersubspecific recombination in the plant-pathogenic bacterium Xylella fastidiosa is associated with the host shift to mulberry. Appl. Environ. Microbiol. 2014, 80, 3025–3033. [Google Scholar] [CrossRef]
- Randall, J.J.; Goldberg, N.P.; Kemp, J.D.; Radionenko, M.; French, J.M.; Olsen, M.W.; Hanson, S.F. Genetic Analysis of a Novel Xylella fastidiosa Subspecies Found in the Southwestern United States. Appl. Environ. Microbiol. 2009, 75, 5631–5638. [Google Scholar] [CrossRef]
- Schuenzel, E.L.; Scally, M.; Stouthamer, R.; Nunney, L. A Multigene Phylogenetic Study of Clonal Diversity and Divergence in North American Strains of the Plant Pathogen Xylella fastidiosa. Appl. Environ. Microbiol. 2005, 71, 3832–3839. [Google Scholar] [CrossRef]
- Cruaud, A.; Gonzalez, A.-A.; Godefroid, M.; Nidelet, S.; Streito, J.-C.; Thuillier, J.-M.; Rossi, J.-P.; Santoni, S.; Rasplus, J.-Y. Using insects to detect, monitor and predict the distribution of Xylella fastidiosa: A case study in Corsica. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Elbaino, T.; Valentini, F.; Abou Kubaa, R.; Moubarak, P.; Yaseen, T.; Digiaro, M. Multilocus sequence typing of Xylella fastidiosa isolated from olive affected by “olive quick decline syndrome” in Italy. Phytopathol. Mediterr. 2014, 53, 533–542. [Google Scholar]
- Jacques, M.-A.; Denancé, N.; Legendre, B.; Morel, E.; Briand, M.; Mississipi, S.; Durand, K.; Olivier, V.; Portier, P.; Poliakoff, F.; et al. New Coffee Plant-Infecting Xylella fastidiosa Variants Derived via Homologous Recombination. Appl. Environ. Microbiol. 2016, 82, 1556. [Google Scholar] [CrossRef]
- Doddapaneni, H.; Francis, M.; Yao, J.; Lin, H.; Civerolo, E.L. Genome-wide analysis of Xylella fastidiosa: Implications for detection and strain relationships. Afr. J. Biotechnol. 2007, 6, 055–066. [Google Scholar]
- Yuan, X.; Morano, L.; Bromley, R.; Spring-Pearson, S.; Stouthamer, R.; Nunney, L. Multilocus Sequence Typing of Xylella fastidiosa Causing Pierce’s Disease and Oleander Leaf Scorch in the United States. Phytopathology 2010, 100, 601–611. [Google Scholar] [CrossRef]
- Burbank, L.P.; Ortega, B.C. Novel amplification targets for rapid detection and differentiation of Xylella fastidiosa subspecies fastidiosa and multiplex in plant and insect tissues. J. Microbiol. Methods 2018, 155, 8–18. [Google Scholar] [CrossRef] [PubMed]
- European and Mediterranean Plant Protection Organization. EPPO PM 7/24 (3) Xylella fastidiosa. EPPO Bull. 2018, 48, 175–218. [Google Scholar] [CrossRef]
- Loconsole, G.; Potere, O.; Boscia, D.; Altamura, G.; Djelouah, K.; Elbeaino, T.; Frasheri, D.; Lorusso, D.; Palmisano, F.; Pollastro, P. Detection of Xylella fastidiosa in olive trees by molecular and serological methods. J. Plant Pathol. 2014, 96, 7–14. [Google Scholar] [CrossRef]
- Loconsole, G.; Saponari, M.; Boscia, D.; D’Attoma, G.; Morelli, M.; Martelli, G.P.; Almeida, R.P.P. Intercepted isolates of Xylella fastidiosa in Europe reveal novel genetic diversity. Eur. J. Plant Pathol. 2016, 146, 85–94. [Google Scholar] [CrossRef]
- Maiden, M.C. Multilocus sequence typing of bacteria. Annu. Rev. Microbiol. 2016, 60, 561–588. [Google Scholar] [CrossRef]
- Horizon 2020. Pest Organisms Threatening Europe. In Proceedings of the XFactors European Conference on Xylella fastidiosa and Project Annual Meetings, Palma de Mallorca, Spain, 13–15 November 2017. [Google Scholar]
- Barbosa, D.; Alencar, V.C.; Santos, D.S.; de Freitas Oliveira, A.C.; de Souza, A.A.; Coletta-Filho, H.D.; Costa de Oliveira, R.; Nunes, L.R. Comparative genomic analysis of coffee-infecting Xylella fastidiosa strains isolated from Brazil. Microbiology 2015, 161, 1018–1033. [Google Scholar] [CrossRef]
- Bleve, G.; Marchi, G.; Rinaldi, F.; Gallo, A.; Cimaglia, F.; Logrieco, A.F.; Mita, G.; Ristori, J.; Surico, G. Molecular characteristics of a strain (Salento-1) of Xylella fastidiosa isolated in Apulia (Italy) from an olive plant with the quick decline syndrome. Phytopathol. Mediterr. 2016, 55, 139–146. [Google Scholar] [CrossRef]
- Parker, J.K.; Havird, J.C.; De La Fuente, L. Differentiation of Xylella fastidiosa Strains via Multilocus Sequence Analysis of Environmentally Mediated Genes (MLSA-E). Appl. Environ. Microbiol. 2012, 78, 1385. [Google Scholar] [CrossRef]
- Cella, E.; Angeletti, S.; Fogolari, M.; Bazzardi, R.; De Gara, L.; Ciccozzi, M. Two different Xylella fastidiosa strains circulating in Italy: Phylogenetic and evolutionary analyses. J. Plant Interact. 2018, 13, 428–432. [Google Scholar] [CrossRef]
- Giampetruzzi, A.; Loconsole, G.; Boscia, D.; Calzolari, A.; Chiumenti, M.; Martelli, G.P.; Saldarelli, P.; Almeida, R.P.; Saponari, M. Draft genome sequence of CO33, a coffee-infecting isolate of Xylella fastidiosa. Genome Announc. 2015, 3, e01472-15. [Google Scholar] [CrossRef] [PubMed]
- Giampetruzzi, A.; Saponari, M.; Loconsole, G.; Boscia, D.; Savino, V.N.; Almeida, R.P.; Zicca, S.; Landa, B.B.; Chacón-Diaz, C.; Saldarelli, P. Genome-wide analysis provides evidence on the genetic relatedness of the emergent Xylella fastidiosa genotype in Italy to isolates from Central America. Phytopathology 2017, 107, 816–827. [Google Scholar] [CrossRef] [PubMed]
- Moralejo, E.; Borràs, D.; Gomila, M.; Montesinos, M.; Adrover, F.; Juan, A.; Nieto, A.; Olmo, D.; Seguí, G.; Landa, B.B. Insights into the epidemiology of Pierce’s disease in vineyards of Mallorca, Spain. Plant Pathol. 2019, 68, 1458–1471. [Google Scholar] [CrossRef]
- Nunney, L.; Yuan, X.; Bromley, R.E.; Stouthamer, R. Detecting Genetic Introgression: High Levels of Intersubspecific Recombination Found in Xylella fastidiosa in Brazil. Appl. Environ. Microbiol. 2012, 78, 4702–4714. [Google Scholar] [CrossRef] [PubMed]
- Nunney, L.; Yuan, X.; Bromley, R.; Hartung, J.; Montero-Astúa, M.; Moreira, L.; Ortiz, B.; Stouthamer, R. Population Genomic Analysis of a Bacterial Plant Pathogen: Novel Insight into the Origin of Pierce’s Disease of Grapevine in the U.S. PLoS ONE 2010, 5, e15488. [Google Scholar] [CrossRef]
- Saponari, M.; Giampetruzzi, A.; Loconsole, G.; Boscia, D.; Saldarelli, P. Xylella fastidiosa in Olive in Apulia: Where We Stand. Phytopathology 2018, 109, 175–186. [Google Scholar] [CrossRef]
- Coletta-Filho, H.D.; Francisco, C.S.; Lopes, J.R.S.; Muller, C.; Almeida, R.P.P. Homologous Recombination and Xylella fastidiosa Host–Pathogen Associations in South America. Phytopathology 2016, 107, 305–312. [Google Scholar] [CrossRef]
- Kung, S.H.; Almeida, R.P.P. Natural Competence and Recombination in the Plant Pathogen Xylella fastidiosa. Appl. Environ. Microbiol. 2011, 77, 5278–5284. [Google Scholar] [CrossRef]
- Scally, M.; Schuenzel, E.L.; Stouthamer, R.; Nunney, L. Multilocus sequence type system for the plant pathogen Xylella fastidiosa and relative contributions of recombination and point mutation to clonal diversity. Appl. Environ. Microbiol. 2005, 71, 8491–8499. [Google Scholar] [CrossRef]
- Chen, H.; De La Fuente, L. Calcium transcriptionally regulates movement, recombination and other functions of Xylella fastidiosa under constant flow inside microfluidic chambers. Microb. Biotechnol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Nunes, L.R.; Rosato, Y.B.; Muto, N.H.; Yanai, G.M.; da Silva, V.S.; Leite, D.B.; Gonçalves, E.R.; de Souza, A.A.; Coletta-Filho, H.D.; Machado, M.A.; et al. Microarray Analyses of Xylella fastidiosa Provide Evidence of Coordinated Transcription Control of Laterally Transferred Elements. Genome Res. 2003, 13, 570–578. [Google Scholar] [CrossRef] [PubMed]
- De Mello Varani, A.; Souza, R.C.; Nakaya, H.I.; de Lima, W.C.; Paula de Almeida, L.G.; Kitajima, E.W.; Chen, J.; Civerolo, E.; Vasconcelos, A.T.R.; Van Sluys, M.-A. Origins of the Xylella fastidiosa Prophage-Like Regions and Their Impact in Genome Differentiation. PLoS ONE 2008, 3. [Google Scholar] [CrossRef] [PubMed]
- Giampetruzzi, A.; Velasco-Amo, M.P.; Marco-Noales, E.; Montes-Borrego, M.; Román-Écija, M.; Navarro, I.; Monterde, A.; Barbé, S.; Almeida, R.P.P.; Saldarelli, P.; et al. Draft Genome Resources of Two Strains (“ESVL” and “IVIA5901”) of Xylella fastidiosa Associated with Almond Leaf Scorch Disease in Alicante, Spain. Phytopathology 2018, 109, 219–221. [Google Scholar] [CrossRef] [PubMed]
- Gomila, M.; Moralejo, E.; Busquets, A.; Segui, G.; Olmo, D.; Nieto, A.; Juan, A.; Lalucat, J. Draft Genome Resources of Two Strains of Xylella fastidiosa XYL1732/17 and XYL2055/17 Isolated from Mallorca Vineyards. Phytopathology 2019, 109, 222–224. [Google Scholar] [CrossRef]
- Van Horn, C.; Wu, F.; Zheng, Z.; Dai, Z.; Chen, J. Detection of a Single-Copy Plasmid, pXFSL21, in Xylella fastidiosa Strain Stag’s Leap with Two Toxin-Antitoxin Systems Using Next-Generation Sequencing. Phytopathology 2018, 109, 240–247. [Google Scholar] [CrossRef]
- Schürch, A.C.; Arredondo-Alonso, S.; Willems, R.J.L.; Goering, R.V. Whole genome sequencing options for bacterial strain typing and epidemiologic analysis based on single nucleotide polymorphism versus gene-by-gene–based approaches. Clin. Microbiol. Infect. 2018, 24, 350–354. [Google Scholar] [CrossRef]
- Yurtsever, S. Population genetics of Philaenus spumaris on the Istrianca mountanins: II. polymorphism and phenotype frequency. Acta Zool. Acad. Sci. Hung. 2004, 50, 25–34. [Google Scholar]
- Rodrigues, A.S.B.; Silva, S.E.; Pina-Martins, F.; Loureiro, J.; Castro, M.; Gharbi, K.; Johnson, K.P.; Dietrich, C.H.; Borges, P.A.V.; Quartau, J.A.; et al. Assessing genotype-phenotype associations in three dorsal colour morphs in the meadow spittlebug Philaenus spumarius (L.) (Hemiptera: Aphrophoridae) using genomic and transcriptomic resources. BMC Genet. 2016, 17, 144. [Google Scholar] [CrossRef]
- Rodrigues, A.S.B.; Silva, S.E.; Marabuto, E.; Silva, D.N.; Wilson, M.R.; Thompson, V.; Yurtsever, S.; Halkka, A.; Borges, P.A.V.; Quartau, J.A.; et al. New Mitochondrial and Nuclear Evidences Support Recent Demographic Expansion and an Atypical Phylogeographic Pattern in the Spittlebug Philaenus spumarius (Hemiptera, Aphrophoridae). PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Campanharo, J.C.; Lemos, M.V.F.; de Lemos, E.G.M. Growth optimization procedures for the phytopathogen Xylella fastidiosa. Curr. Microbiol. 2003, 46, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, M.; Luvisi, A.; Meyer, J.B.; Sabella, E.; De Bellis, L.; Cruz, A.C.; Ampatzidis, Y.; Cherubini, P. Specific Fluorescence in Situ Hybridization (FISH) Test to Highlight Colonization of Xylem Vessels by Xylella fastidiosa in Naturally Infected Olive Trees (Olea europaea L.). Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Djelouah, K.; Frasheri, D.; Valentini, F.; D’Onghia, A.M.; Digiaro, M. Direct tissue blot immunoassay for detection of Xylella fastidiosa in olive trees. Phytopathol. Mediterr. 2014, 53, 559–564. [Google Scholar]
- Potere, O.; Susca, L.; Loconsole, G.; Saponari, M.; Boscia, D.; Savino, V.N.; Martelli, G.P. Survey for the presence of Xylella fastidiosa subsp. pauca strain CoDiRO in some forestry and ornamental species in the Salento peninsula. J. Plant Pathol. 2015, 97, 373–376. [Google Scholar]
- Saponari, M.; Boscia, D.; Altamura, G.; D’Attoma, G.; Cavalieri, V.; Zicca, S.; Morelli, M.; Tavano, D.; Loconsole, G.; Susca, L. Pilot project on Xylella fastidiosa to reduce risk assessment uncertainties. EFSA Support. Publ. 2016, 13, EN-1013. [Google Scholar] [CrossRef]
- Novelli, S.; Gismondi, A.; Di Marco, G.; Canuti, L.; Nanni, V.; Canini, A. Plant defense factors involved in Olea europaea resistance against Xylella fastidiosa infection. J. Plant Res. 2019, 132, 439–455. [Google Scholar] [CrossRef] [PubMed]
- Reisenzein, H. PCR assays for the detection of Xylella fastidiosa: Review and comparison of published protocols. In Xylella Fastidiosa & the Olive Quick Decline Syndrome (OQDS): A Serious Worldwide Challenge for the Safeguard of Olive Trees; CIHEAM: Bari, Italy, 2017; pp. 57–60. [Google Scholar]
- Martinelli, F.; Scalenghe, R.; Davino, S.; Panno, S.; Scuderi, G.; Ruisi, P.; Villa, P.; Stroppiana, D.; Boschetti, M.; Goulart, L.R.; et al. Advanced methods of plant disease detection. A review. Agron. Sustain. Dev. 2015, 35, 1–25. [Google Scholar] [CrossRef]
- Modesti, V.; Pucci, N.; Lucchesi, S.; Campus, L.; Loreti, S. Experience of the Latium region (Central Italy) as a pest-free area for monitoring of Xylella fastidiosa: Distinctive features of molecular diagnostic methods. Eur. J. Plant Pathol. 2017, 148, 557–566. [Google Scholar] [CrossRef]
- Bonants, P.; Griekspoor, Y.; Houwers, I.; Krijger, M.; van der Zouwen, P.; van der Lee, T.A.; van der Wolf, J. Development and Evaluation of a Triplex TaqMan Assay and Next-Generation Sequence Analysis for Improved Detection of Xylella in Plant Material. Plant Dis. 2019, 103, 645–655. [Google Scholar] [CrossRef]
- Dupas, E.; Briand, M.; Jacques, M.-A.; Cesbron, S. Novel tetraplex qPCR assays for simultaneous detection and identification of Xylella fastidiosa subspecies in plant tissues. Front. Plant Sci. 2019, 10, 1732. [Google Scholar] [CrossRef]
- Faino, L.; Scala, V.; Albanese, A.; Modesti, V.; Grottoli, A.; Pucci, N.; L’Aurora, A.; Reverberi, M.; Loreti, S. Nanopore sequencing for the detection and the identification of Xylella fastidiosa subspecies and sequence types from naturally infected plant material. bioRxiv 2019, 810648. [Google Scholar] [CrossRef]
- Chiriacò, M.S.; Luvisi, A.; Primiceri, E.; Sabella, E.; De Bellis, L.; Maruccio, G. Development of a lab-on-a-chip method for rapid assay of Xylella fastidiosa subsp. pauca strain CoDiRO. Sci. Rep. 2018, 8, 7376. [Google Scholar] [CrossRef] [PubMed]
- Schrader, C.; Schielke, A.; Ellerbroek, L.; Johne, R. PCR inhibitors–occurrence, properties and removal. J. Appl. Microbiol. 2012, 113, 1014–1026. [Google Scholar] [CrossRef] [PubMed]
- Harper, S.J.; Ward, L.I.; Clover, G.R.G. Development of LAMP and real-time PCR methods for the rapid detection of Xylella fastidiosa for quarantine and field applications. Phytopathology 2010, 100, 1282–1288. [Google Scholar] [CrossRef]
- Sipahioglu, H.M.; Usta, M.; Ocak, M. Use of dried high-phenolic laden host leaves for virus and viroid preservation and detection by PCR methods. J. Virol. Methods 2006, 137, 120–124. [Google Scholar] [CrossRef]
- Witte, A.K.; Sickha, R.; Mester, P.; Fister, S.; Schoder, D.; Rossmanith, P. Essential role of polymerases for assay performance–Impact of polymerase replacement in a well-established assay. Biomol. Detect. Quantif. 2018, 16, 12–20. [Google Scholar] [CrossRef]
- Martinelli, F.; Marchese, A.; Giovino, A.; Marra, F.P.; Della Noce, I.; Caruso, T.; Dandekar, A.M. In-Field and Early Detection of Xylella fastidiosa Infections in Olive Using a Portable Instrument. Front. Plant Sci. 2019, 9. [Google Scholar] [CrossRef]
- Nicolì, F.; Negro, C.; Nutricati, E.; Vergine, M.; Aprile, A.; Sabella, E.; Damiano, G.; De Bellis, L.; Luvisi, A. Accumulation of Azelaic Acid in Xylella fastidiosa-Infected Olive Trees: A Mobile Metabolite for Health Screening. Phytopathology 2018, 109, 318–325. [Google Scholar] [CrossRef]
- Boncristiani, H.; Li, J.; Evans, J.D.; Pettis, J.; Chen, Y. Scientific note on PCR inhibitors in the compound eyes of honey bees, Apis mellifera. Apidologie 2011, 42, 457–460. [Google Scholar] [CrossRef]
- Shamim, G.; Ranjan, S.K.; Pandey, D.M.; Ramani, R. Biochemistry and biosynthesis of insect pigments. EJE 2014, 111, 149–164. [Google Scholar] [CrossRef]
- European and Mediterranean Plant Protection Organization. EPPO PM 7/129 (1) DNA barcoding as an identification tool for a number of regulated pests. EPPO Bull. 2016, 46, 501–537. [Google Scholar] [CrossRef]
- Rach, J.; Bergmann, T.; Paknia, O.; DeSalle, R.; Schierwater, B.; Hadrys, H. The marker choice: Unexpected resolving power of an unexplored CO1 region for layered DNA barcoding approaches. PLoS ONE 2017, 12, e0174842. [Google Scholar] [CrossRef] [PubMed]
- Marquina, D.; Andersson, A.F.; Ronquist, F. New mitochondrial primers for metabarcoding of insects, designed and evaluated using in silico methods. Mol. Ecol. Resour. 2019, 19, 90–104. [Google Scholar] [CrossRef] [PubMed]
- Crampton-Platt, A.; Yu, D.W.; Zhou, X.; Vogler, A.P. Mitochondrial metagenomics: Letting the genes out of the bottle. GigaScience 2016, 5, 15. [Google Scholar] [CrossRef] [PubMed]
- Cicconardi, F.; Borges, P.A.V.; Strasberg, D.; Oromí, P.; López, H.; Pérez-Delgado, A.J.; Casquet, J.; Caujapé-Castells, J.; Fernández-Palacios, J.M.; Thébaud, C.; et al. MtDNA metagenomics reveals large-scale invasion of belowground arthropod communities by introduced species. Mol. Ecol. 2017, 26, 3104–3115. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Li, Y.; Liu, S.; Yang, Q.; Su, X.; Zhou, L.; Tang, M.; Fu, R.; Li, J.; Huang, Q. Ultra-deep sequencing enables high-fidelity recovery of biodiversity for bulk arthropod samples without PCR amplification. GigaScience 2013, 2, 4. [Google Scholar] [CrossRef]
- Deiner, K.; Bik, H.M.; Mächler, E.; Seymour, M.; Lacoursière-Roussel, A.; Altermatt, F.; Creer, S.; Bista, I.; Lodge, D.M.; de Vere, N.; et al. Environmental DNA metabarcoding: Transforming how we survey animal and plant communities. Mol. Ecol. 2017, 26, 5872–5895. [Google Scholar] [CrossRef]
- Ciraulo, M.B.; Santos, D.S.; de Rodrigues, A.C.F.O.; de Oliveira, M.V.; Rodrigues, T.; de Oliveira, R.C.; Nunes, L.R. Transcriptome Analysis of the Phytobacterium Xylella fastidiosa Growing under Xylem-Based Chemical Conditions. Available online: https://www.hindawi.com/journals/bmri/2010/781365/ (accessed on 4 December 2019).
- Federici, M.T.; Marcondes, J.A.; Picchi, S.C.; Stuchi, E.S.; Fadel, A.L.; Laia, M.L.; Lemos, M.V.F.; Lemos, E.G.M. Xylella fastidiosa: An in vivo system to study possible survival strategies within citrus xylem vessels based on global gene expression analysis. Electron. J. Biotechnol. 2012, 15, 4. [Google Scholar] [CrossRef]
- De Travensolo, R.F.; Costa, M.V.C.G.; Carareto-Alves, L.M.; Carrilho, E.; de Lemos, E.G.M. Production of DNA microarray and expression analysis of genes from Xylella fastidiosa in different culture media. Braz. Arch. Biol. Technol. 2009, 52, 555–566. [Google Scholar] [CrossRef]
- Travensolo, R.F.; Carareto-Alves, L.M.; Costa, M.V.C.G.; Lopes, T.J.S.; Carrilho, E.; Lemos, E.G.M. Xylella fastidiosa gene expression analysis by DNA microarrays. Genet. Mol. Biol. 2009, 32, 340–353. [Google Scholar] [CrossRef]
- Gambetta, G.A.; Matthews, M.A.; Syvanen, M. The Xylella fastidosa RTX operons: Evidence for the evolution of protein mosaics through novel genetic exchanges. BMC Genom. 2018, 19, 329. [Google Scholar] [CrossRef]
- Li, W.B.; Zhou, C.H.; Pria, W.D.; Teixeira, D.C.; Miranda, V.S.; Pereira, E.O.; Ayres, A.J.; He, C.-X.; Costa, P.I.; Hartung, J.S. Citrus and Coffee Strains of Xylella fastidiosa Induce Pierce’s Disease in Grapevine. Plant Dis. 2002, 86, 1206–1210. [Google Scholar] [CrossRef]
- Abou Kubaa, R.; Giampetruzzi, A.; Altamura, G.; Saponari, M.; Saldarelli, P. Infections of the Xylella fastidiosa subsp. pauca Strain “De Donno” in Alfalfa (Medicago sativa) Elicits an Overactive Immune Response. Plants 2019, 8, 335. [Google Scholar] [CrossRef]
- Sabella, E.; Luvisi, A.; Aprile, A.; Negro, C.; Vergine, M.; Nicolì, F.; Miceli, A.; De Bellis, L. Xylella fastidiosa induces differential expression of lignification related-genes and lignin accumulation in tolerant olive trees cv. Leccino. J. Plant Physiol. 2018, 220, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Zaini, P.A.; Nascimento, R.; Gouran, H.; Cantu, D.; Chakraborty, S.; Phu, M.; Goulart, L.R.; Dandekar, A.M. Molecular Profiling of Pierce’s Disease Outlines the Response Circuitry of Vitis vinifera to Xylella fastidiosa Infection. Front. Plant Sci. 2018, 9, 771. [Google Scholar] [CrossRef] [PubMed]
- De Pascali, M.; Vergine, M.; Sabella, E.; Aprile, A.; Nutricati, E.; Nicolì, F.; Buja, I.; Negro, C.; Miceli, A.; Rampino, P.; et al. Molecular Effects of Xylella fastidiosa and Drought Combined Stress in Olive Trees. Plants 2019, 8, 437. [Google Scholar] [CrossRef] [PubMed]
- Rapicavoli, J.; Ingel, B.; Blanco-Ulate, B.; Cantu, D.; Roper, C. Xylella fastidiosa: An examination of a re-emerging plant pathogen. Mol. Plant Pathol. 2018, 19, 786–800. [Google Scholar] [CrossRef]
- Giampetruzzi, A.; Morelli, M.; Saponari, M.; Loconsole, G.; Chiumenti, M.; Boscia, D.; Savino, V.N.; Martelli, G.P.; Saldarelli, P. Transcriptome profiling of two olive cultivars in response to infection by the CoDiRO strain of Xylella fastidiosa subsp. pauca. BMC Genom. 2016, 17, 475. [Google Scholar] [CrossRef]
- D’Attoma, G.; Morelli, M.; Saldarelli, P.; Saponari, M.; Giampetruzzi, A.; Boscia, D.; Savino, V.N.; De La Fuente, L.; Cobine, P.A. Ionomic Differences between Susceptible and Resistant Olive Cultivars Infected by Xylella fastidiosa in the Outbreak Area of Salento, Italy. Pathogens 2019, 8, 272. [Google Scholar] [CrossRef]
- Parker, J.K.; Chen, H.; McCarty, S.E.; Liu, L.Y.; De La Fuente, L. Calcium transcriptionally regulates the biofilm machinery of Xylella fastidiosa to promote continued biofilm development in batch cultures. Environ. Microbiol. 2016, 18, 1620–1634. [Google Scholar] [CrossRef]
- Rodrigues, C.M.; de Souza, A.A.; Takita, M.A.; Kishi, L.T.; Machado, M.A. RNA-Seq analysis of Citrus reticulata in the early stages of Xylella fastidiosa infection reveals auxin-related genes as a defense response. BMC Genom. 2013, 14, 676. [Google Scholar] [CrossRef] [PubMed]
- Workman, R.E.; Tang, A.D.; Tang, P.S.; Jain, M.; Tyson, J.R.; Razaghi, R.; Zuzarte, P.C.; Gilpatrick, T.; Payne, A.; Quick, J.; et al. Nanopore native RNA sequencing of a human poly(A) transcriptome. Nat. Methods 2019, 16, 1297–1305. [Google Scholar] [CrossRef]
- Kukurba, K.R.; Montgomery, S.B. RNA Sequencing and Analysis. Cold Spring Harb. Protoc. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Mathers, T.C.; Chen, Y.; Kaithakottil, G.; Legeai, F.; Mugford, S.T.; Baa-Puyoulet, P.; Bretaudeau, A.; Clavijo, B.; Colella, S.; Collin, O.; et al. Rapid transcriptional plasticity of duplicated gene clusters enables a clonally reproducing aphid to colonise diverse plant species. Genome Biol. 2017, 18, 27. [Google Scholar] [CrossRef] [PubMed]
- De la Celorio-Mancera, M.P.; Wheat, C.W.; Huss, M.; Vezzi, F.; Neethiraj, R.; Reimegård, J.; Nylin, S.; Janz, N. Evolutionary history of host use, rather than plant phylogeny, determines gene expression in a generalist butterfly. BMC Evol. Biol. 2016, 16, 59. [Google Scholar] [CrossRef] [PubMed]
- Pym, A.; Singh, K.S.; Nordgren, Å.; Davies, T.G.E.; Zimmer, C.T.; Elias, J.; Slater, R.; Bass, C. Host plant adaptation in the polyphagous whitefly, Trialeurodes vaporariorum, is associated with transcriptional plasticity and altered sensitivity to insecticides. BMC Genom. 2019, 20, 996. [Google Scholar] [CrossRef]
- Chen, C.; Bock, C.H.; Brannen, P.M. Novel Primers and Sampling for PCR Detection of Xylella fastidiosa in Peach. Phytopathology 2019, 109, 307–317. [Google Scholar] [CrossRef]
- Shelake, R.M.; Pramanik, D.; Kim, J.-Y. Exploration of Plant-Microbe Interactions for Sustainable Agriculture in CRISPR Era. Microorganisms 2019, 7, 269. [Google Scholar] [CrossRef]
- Da Silva Neto, J.F.; Koide, T.; Gomes, S.L.; Marques, M.V. Site-directed gene disruption in Xylella fastidiosa. FEMS Microbiol. Lett. 2002, 210, 105–110. [Google Scholar] [CrossRef][Green Version]
- Kandel, P.P.; Chen, H.; De La Fuente, L. A Short Protocol for Gene Knockout and Complementation in Xylella fastidiosa Shows that One of the Type IV Pilin Paralogs (PD1926) Is Needed for Twitching while Another (PD1924) Affects Pilus Number and Location. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, P.B.; Teixeira, D.C.; Palma, R.R.; Garnier, M.; Bové, J.-M.; Renaudin, J. Stable Transformation of the Xylella fastidiosa Citrus Variegated Chlorosis Strain withoriC Plasmids. Appl. Environ. Microbiol. 2001, 67, 2263–2269. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Fong, S.S. Challenges and Advances for Genetic Engineering of Non-model Bacteria and Uses in Consolidated Bioprocessing. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Cruz, L.F.; Parker, J.K.; Cobine, P.A.; Fuente, L.D.L. Calcium-Enhanced Twitching Motility in Xylella fastidiosa Is Linked to a Single PilY1 Homolog. Appl. Environ. Microbiol. 2014, 80, 7176–7185. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, A.; Young, G.M.; Igo, M.M. Chromosome-Based Genetic Complementation System for Xylella fastidiosa. Appl. Environ. Microbiol. 2009, 75, 1679–1687. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Li, Y.; Galvani, C.D.; Hao, G.; Turner, J.N.; Burr, T.J.; Hoch, H.C. Upstream Migration of Xylella fastidiosa via Pilus-Driven Twitching Motility. J. Bacteriol. 2005, 187, 5560–5567. [Google Scholar] [CrossRef] [PubMed]
- Navarrete, F.; De La Fuente, L. Zinc Detoxification Is Required for Full Virulence and Modification of the Host Leaf Ionome by Xylella fastidiosa. Mol. Plant-Microbe Interact. 2015, 28, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Newman, K.L.; Almeida, R.P.P.; Purcell, A.H.; Lindow, S.E. Cell-cell signaling controls Xylella fastidiosa interactions with both insects and plants. Proc. Natl. Acad. Sci. USA 2004, 101, 1737–1742. [Google Scholar] [CrossRef]
- Roper, M.C.; Greve, L.C.; Warren, J.G.; Labavitch, J.M.; Kirkpatrick, B.C. Xylella fastidiosa Requires Polygalacturonase for Colonization and Pathogenicity in Vitis vinifera Grapevines. Mol. Plant-Microbe Interact. 2007, 20, 411–419. [Google Scholar] [CrossRef]
- Hao, L.; Johnson, K.; Cursino, L.; Mowery, P.; Burr, T.J. Characterization of the Xylella fastidiosa PD1311 gene mutant and its suppression of Pierce’s disease on grapevines. Mol. Plant Pathol. 2017, 18, 684–694. [Google Scholar] [CrossRef]
- Killiny, N.; Almeida, R.P.P. Gene regulation mediates host specificity of a bacterial pathogen. Environ. Microbiol. Rep. 2011, 3, 791–797. [Google Scholar] [CrossRef]
- Lee, M.W.; Tan, C.C.; Rogers, E.E.; Stenger, D.C. Toxin-antitoxin systems mqsR/ygiT and dinJ/relE of Xylella fastidiosa. Physiol. Mol. Plant Pathol. 2014, 87, 59–68. [Google Scholar] [CrossRef]
- Merfa, M.V.; Niza, B.; Takita, M.A.; De Souza, A.A. The MqsRA Toxin-Antitoxin System from Xylella fastidiosa Plays a Key Role in Bacterial Fitness, Pathogenicity, and Persister Cell Formation. Front. Microbiol. 2016, 7, 904. [Google Scholar] [CrossRef] [PubMed]
- Reddy, J.D.; Reddy, S.L.; Hopkins, D.L.; Gabriel, D.W. TolC is Required for Pathogenicity of Xylella fastidiosa in Vitis vinifera Grapevines. Mol. Plant-Microbe Interact. 2007, 20, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Das, M.; Bhowmick, T.S.; Ahern, S.J.; Young, R.; Gonzalez, C.F. Control of Pierce’s Disease by Phage. PLoS ONE 2015, 10, e0128902. [Google Scholar] [CrossRef] [PubMed]
- Dandekar, A.M.; Gouran, H.; Ibáñez, A.M.; Uratsu, S.L.; Agüero, C.B.; McFarland, S.; Borhani, Y.; Feldstein, P.A.; Bruening, G.; Nascimento, R.; et al. An engineered innate immune defense protects grapevines from Pierce disease. Proc. Natl. Acad. Sci. USA 2012, 109, 3721. [Google Scholar] [CrossRef]
- Li, Z.T.; Hopkins, D.L.; Gray, D.J. Overexpression of antimicrobial lytic peptides protects grapevine from Pierce’s disease under greenhouse but not field conditions. Transgenic Res. 2015, 24, 821–836. [Google Scholar] [CrossRef]
- Caserta, R.; Souza-Neto, R.R.; Takita, M.A.; Lindow, S.E.; De Souza, A.A. Ectopic Expression of Xylella fastidiosa rpfF Conferring Production of Diffusible Signal Factor in Transgenic Tobacco and Citrus Alters Pathogen Behavior and Reduces Disease Severity. Mol. Plant-Microbe Interact. 2017, 30, 866–875. [Google Scholar] [CrossRef]
- Gilchrist, D.; Lincoln, J.E.; Esser, T. Systemic control of Pierce’s disease by altered expression of anti-apoptotic genes or their RNA-based regulatory elements. In Proceedings of the Pierce’s Disease Research Symposium, San Diego, CA, USA, 15–17 December 2008; pp. 208–213. [Google Scholar]
- Lindow, S.; Newman, K.; Chatterjee, S.; Baccari, C.; Iavarone, A.T.; Ionescu, M. Production of Xylella fastidiosa Diffusible Signal Factor in Transgenic Grape Causes Pathogen Confusion and Reduction in Severity of Pierce’s Disease. Mol. Plant-Microbe Interact. 2014, 27, 244–254. [Google Scholar] [CrossRef]
- Zaidi, S.S.-A.; Mukhtar, M.S.; Mansoor, S. Genome Editing: Targeting Susceptibility Genes for Plant Disease Resistance. Trends Biotechnol. 2018, 36, 898–906. [Google Scholar] [CrossRef]
- Low, L.-Y.; Yang, S.-K.; Kok, D.-X.A.; Ong-Abdullah, J.; Tan, N.-P.; Lai, K.-S. Transgenic Plants: Gene Constructs, Vector and Transformation Method. In New Visions in Plant Science; IntechOpen: London, UK, 2018. [Google Scholar]
- Durvasula, R.V.; Gumbs, A.; Panackal, A.; Kruglov, O.; Aksoy, S.; Merrifield, R.B.; Richards, F.F.; Beard, C.B. Prevention of insect-borne disease: An approach using transgenic symbiotic bacteria. Proc. Natl. Acad. Sci. USA 1997, 94, 3274–3278. [Google Scholar] [CrossRef]
- Aksoy, H.M.; Ozman-Sullivan, S.K.; Ocal, H.; Celik, N.; Sullivan, G.T. The effects of Pseudomonas putida biotype B on Tetranychus urticae (Acari: Tetranychidae). Exp. Appl. Acarol. 2008, 46, 223. [Google Scholar] [CrossRef]
- Hurwitz, I.; Hillesland, H.; Fieck, A.; Das, P.; Durvasula, R. The paratransgenic sand fly: A platform for control of Leishmania transmission. Parasit. Vectors 2011, 4, 82. [Google Scholar] [CrossRef]
- Wang, S.; Ghosh, A.K.; Bongio, N.; Stebbings, K.A.; Lampe, D.J.; Jacobs-Lorena, M. Fighting malaria with engineered symbiotic bacteria from vector mosquitoes. Proc. Natl. Acad. Sci. USA 2012, 109, 12734–12739. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.L.; Perring, T.M.; Miller, T.A. Fate of a genetically modified bacterium in foregut of glassy-winged sharpshooter (Hemiptera: Cicadellidae). J. Econ. Entomol. 2008, 101, 1519–1525. [Google Scholar] [CrossRef] [PubMed]
- Arora, A.K.; Pesko, K.N.; Quintero-Hernández, V.; Possani, L.D.; Miller, T.A.; Durvasula, R.V. A paratransgenic strategy to block transmission of Xylella fastidiosa from the glassy-winged sharpshooter Homalodisca vitripennis. BMC Biotechnol. 2018, 18, 50. [Google Scholar] [CrossRef]
- European Food Safety Authority. Workshop on Xylella Fastidiosa: Knowledge Gaps and Research Priorities for the EU; European Food Safety Authority: Parma, Italy, 2016; p. 1039. [Google Scholar]
- Saporta, R.; Pedro, T.S.; Gisbert, C. Attempts at grapevine (Vitis vinifera L.) breeding through genetic transformation: The main limiting factors. VITIS J. Grapevine Res. 2016, 55, 173–186. [Google Scholar] [CrossRef]
- Sharma, A.; Jones, J.B.; White, F.F. Recent advances in developing disease resistance in plants. F1000Research 2019, 8, 1934. [Google Scholar] [CrossRef] [PubMed]
- Gaj, T.; Gersbach, C.A.; Barbas, C.F. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2013, 31, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Gray, D.J.; Li, Z.T.; Dhekney, S.A. Precision breeding of grapevine (Vitis vinifera L.) for improved traits. Plant Sci. 2014, 228, 3–10. [Google Scholar] [CrossRef]
- Hastings, A.; Petrovskii, S.; Morozov, A. Spatial ecology across scales. Biol. Lett. 2011, 7, 163–165. [Google Scholar] [CrossRef]
- Fletcher, R.; Fortin, M.-J. Introduction to Spatial Ecology and Its Relevance for Conservation. In Spatial Ecology and Conservation Modeling; Springer: Cham, Switzerland, 2018; pp. 1–13. ISBN 978-3-030-01989-1. [Google Scholar]
- Lewis, M.A.; Maini, P.K.; Petrovskii, S.V. Dispersal, Individual Movement and Spatial Ecology; Springer: Berlin/Heidelberg, Germany, 2013; Volume 2071. [Google Scholar]
- Santos, M.; Bastos, R.; Vicente, J.; Berger, U.; Soares Filho, B.S.; Rodrigues, H.; Alonso, J.; Guerra, C.; Martins, J.; Honrado, J. 17. Anticipating Invasions and Managing Impacts: A Review of Recent Spatiotemporal Modelling Approaches. In Biological Invasions in Changing Ecosystems; De Gruyter: Berlin, Germany, 2015; pp. 389–410. ISBN 3-11-043866-6. [Google Scholar]
- Cunniffe, N.J.; Koskella, B.; Metcalf, C.J.E.; Parnell, S.; Gottwald, T.R.; Gilligan, C.A. Thirteen challenges in modelling plant diseases. Epidemics 2015, 10, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Joshi, C.; de Leeuw, J.; van Duren, I.C. Remote sensing and GIS applications for mapping and spatial modelling of invasive species. In Proceedings of the ISPRS 2004: Proceedings of the XXth ISPRS Congress: Geo-Imagery Bridging Continents, Istanbul, Turkey, 12–23 July 2004. [Google Scholar]
- Leitão, P.J.; Santos, M.J. Improving models of species ecological niches: A remote sensing overview. Front. Ecol. Evol. 2019, 7, 9. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Vaz, A.S.; Segura, D.A.; Vicente, J.R.; Honrado, J.P. The many roles of remote sensing in invasion science. Front. Ecol. Evol. 2019, 7, 370. [Google Scholar] [CrossRef]
- Ellsbury, M.; Clay, S.; Fleischer, S.; Chandler, L.; Schneider, S. Use of GIS/GPS Systems in IPM: Progress and Reality; American Phytopathological Society Press: Saint Paul, MN, USA, 2000; pp. 419–438. [Google Scholar]
- Jeger, M.; Bragard, C. The epidemiology of Xylella fastidiosa; A perspective on current knowledge and framework to investigate plant host–vector–pathogen interactions. Phytopathology 2019, 109, 200–209. [Google Scholar] [CrossRef]
- Lloyd-Smith, J.O.; Funk, S.; McLean, A.R.; Riley, S.; Wood, J.L. Nine challenges in modelling the emergence of novel pathogens. Epidemics 2015, 10, 35–39. [Google Scholar] [CrossRef]
- Stone, C.; Mohammed, C. Application of remote sensing technologies for assessing planted forests damaged by insect pests and fungal pathogens: A review. Curr. For. Rep. 2017, 3, 75–92. [Google Scholar] [CrossRef]
- Feil, H.; Purcell, A.H. Temperature-dependent growth and survival of Xylella fastidiosa in vitro and in potted grapevines. Plant Dis. 2001, 85, 1230–1234. [Google Scholar] [CrossRef]
- Hoddle, M.S. The potential adventive geographic range of glassy-winged sharpshooter, Homalodisca coagulata and the grape pathogen Xylella fastidiosa: Implications for California and other grape growing regions of the world. Crop Prot. 2004, 23, 691–699. [Google Scholar] [CrossRef]
- Bosso, L.; Di Febbraro, M.; Cristinzio, G.; Zoina, A.; Russo, D. Shedding light on the effects of climate change on the potential distribution of Xylella fastidiosa in the Mediterranean basin. Biol. Invasions 2016, 18, 1759–1768. [Google Scholar] [CrossRef]
- Godefroid, M.; Cruaud, A.; Streito, J.-C.; Rasplus, J.-Y.; Rossi, J.-P. Xylella fastidiosa: Climate suitability of European continent. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.; Camino, C.; Beck, P.; Calderon, R.; Hornero, A.; Hernández-Clemente, R.; Kattenborn, T.; Montes-Borrego, M.; Susca, L.; Morelli, M. Previsual symptoms of Xylella fastidiosa infection revealed in spectral plant-trait alterations. Nat. Plants 2018, 4, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.M.; Bachik, N.A.; Muhadi, N.; Yusof, T.N.T.; Gomes, C. Non-Destructive Techniques of Detecting Plant Diseases: A Review. Physiol. Mol. Plant Pathol. 2019, 108, 101426. [Google Scholar] [CrossRef]
- Mahlein, A.-K. Plant disease detection by imaging sensors–parallels and specific demands for precision agriculture and plant phenotyping. Plant Dis. 2016, 100, 241–251. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Workshop on Xylella fastidiosa: Knowledge gaps and research priorities for the EU. EFSA Support. Publ. 2016, 13, 1039E. [Google Scholar]
- Hornero, A.; Hernández-Clemente, R.; North, P.; Beck, P.; Boscia, D.; Navas-Cortes, J.; Zarco-Tejada, P. Monitoring the incidence of Xylella fastidiosa infection in olive orchards using ground-based evaluations, airborne imaging spectroscopy and Sentinel-2 time series through 3-D radiative transfer modelling. Remote Sens. Environ. 2020, 236, 111480. [Google Scholar] [CrossRef]
- Hornero, A.; Hernández-Clemente, R.; Beck, P.S.; Navas-Cortés, J.A.; Zarco-Tejada, P.J. Using Sentinel-2 Imagery to Track Changes Produced by Xylella Fastidiosa in Olive Trees; IEEE: Piscataway Township, NJ, USA, 2018; pp. 9060–9062. [Google Scholar]
- Poblete, T.; Camino, C.; Beck, P.; Hornero, A.; Kattenborn, T.; Saponari, M.; Boscia, D.; Navas-Cortes, J.; Zarco-Tejada, P. Detection of Xylella fastidiosa infection symptoms with airborne multispectral and thermal imagery: Assessing bandset reduction performance from hyperspectral analysis. ISPRS J. Photogramm. Remote Sens. 2020, 162, 27–40. [Google Scholar] [CrossRef]
- Rey, B.; Aleixos, N.; Cubero, S.; Blasco, J. XF-ROVIM. A field robot to detect olive trees infected by Xylella fastidiosa using proximal sensing. Remote Sens. 2019, 11, 221. [Google Scholar] [CrossRef]
- Scortichini, M.; Jianchi, C.; De Caroli, M.; Dalessandro, G.; Pucci, N.; Modesti, V.; L’Aurora, A.; Petriccione, M.; Zampella, L.; Mastrobuoni, F. A zinc, copper and citric acid biocomplex shows promise for control of Xylella fastidiosa subsp. pauca in olive trees in Apulia region (southern Italy). Phytopathol. Mediterr. 2018, 57, 48–72. [Google Scholar] [CrossRef]
- Nouri, H.; Beecham, S.; Anderson, S.; Nagler, P. High spatial resolution WorldView-2 imagery for mapping NDVI and its relationship to temporal urban landscape evapotranspiration factors. Remote Sens. 2014, 6, 580–602. [Google Scholar] [CrossRef]
- Rouse, J.; Haas, R.; Schell, J.; Deering, D. Monitoring vegetation systems in the Great Plains with ERTS. NASA Spec. Publ. 1974, 351, 309. [Google Scholar]
- Isip, M.; Alberto, R.; Biagtan, A. Exploring vegetation indices adequate in detecting twister disease of onion using Sentinel-2 imagery. Spat. Inf. Res. 2019, 1–7. [Google Scholar] [CrossRef]
- Lugonja, P.; Brdar, S.; Simović, I.; Mimić, G.; Palamarchuk, Y.; Sofiev, M.; Šikoparija, B. Integration of in situ and satellite data for top-down mapping of Ambrosia infection level. Remote Sens. Environ. 2019, 235, 111455. [Google Scholar] [CrossRef]
- Ballester, C.; Zarco-Tejada, P.; Nicolás, E.; Alarcón, J.; Fereres, E.; Intrigliolo, D.S.; Gonzalez-Dugo, V. Evaluating the performance of xanthophyll, chlorophyll and structure-sensitive spectral indices to detect water stress in five fruit tree species. Precis. Agric. 2018, 19, 178–193. [Google Scholar] [CrossRef]
- Calderón, R.; Navas-Cortés, J.A.; Lucena, C.; Zarco-Tejada, P.J. High-resolution airborne hyperspectral and thermal imagery for early detection of Verticillium wilt of olive using fluorescence, temperature and narrow-band spectral indices. Remote Sens. Environ. 2013, 139, 231–245. [Google Scholar] [CrossRef]
- Singh, V.; Rana, A.; Bishop, M.; Filippi, A.M.; Bagavathiannan, M.; Nadon, B.; Jackson, S.; Wang, W.; He, A.; Jiang, G. Unmanned aircraft systems for precision weed detection and management: Prospects and challenges. In Advances in Agronomy; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar] [CrossRef]
- Mohammed, G.H.; Colombo, R.; Middleton, E.M.; Rascher, U.; van der Tol, C.; Nedbal, L.; Goulas, Y.; Pérez-Priego, O.; Damm, A.; Meroni, M. Remote sensing of solar-induced chlorophyll fluorescence (SIF) in vegetation: 50 years of progress. Remote Sens. Environ. 2019, 231, 111177. [Google Scholar] [CrossRef]
- European Space Agency. Report for Mission Selection: FLEX; European Space Agency: Paris, France, 2015; p. 197. [Google Scholar]
- Middleton, E.M.; Rascher, U.; Huemmrich, K.F.; Cook, B.D.; Noormets, A.; Schickling, A.; Pinto, F.; Alonso, L.; Damm, A.; Guanter, L. The 2013 FLEX—US airborne campaign at the parker tract loblolly pine plantation in North Carolina, USA. Remote Sens. 2017, 9, 612. [Google Scholar] [CrossRef]
- Martinetti, D.; Soubeyrand, S. Identifying lookouts for epidemio-surveillance: Application to the emergence of Xylella fastidiosa in France. Phytopathology 2019, 109, 265–276. [Google Scholar] [CrossRef]
- Poliakoff, F.; Legendre, B.; Juteau, V.; Molusson, D.; Dintheer, A.; Sainte-Luce, A.; Dousset, C.; Audusseau, A.; Paillard, S.; Cunty, A.; et al. Current Situation in France Regarding Xylella fastidiosa: Recent Developments and Validation of Detection Method on Plants and Vector Philaenus spumarius. Available online: https://figshare.com/articles/Current_situation_in_France_regarding_Xylella_fastidiosa_recent_developments_and_validation_of_detection_method_on_plants_and_vector_Philaenus_spumarius_/11704422 (accessed on 24 January 2020).
- EFSA. Update of the Xylella spp. host plant database. EFSA J. 2018, 16, e05408. [Google Scholar] [CrossRef]
- Bosso, L.; Russo, D.; Di Febbraro, M.; Cristinzio, G.; Zoina, A. Potential distribution of Xylella fastidiosa in Italy: A maximum entropy model. Phytopathol. Mediterr. 2016, 62–72. [Google Scholar] [CrossRef]
- Cornara, D.; Bosco, D.; Fereres, A. Philaenus spumarius: When an old acquaintance becomes a new threat to European agriculture. J. Pest Sci. 2018, 91, 957–972. [Google Scholar] [CrossRef]
- Dongiovanni, C.; Cavalieri, V.; Bodino, N.; Tauro, D.; Di Carolo, M.; Fumarola, G.; Altamura, G.; Lasorella, C.; Bosco, D. Plant selection and population trend of spittlebug immatures (Hemiptera: Aphrophoridae) in olive groves of the Apulia Region of Italy. J. Econ. Entomol. 2019, 112, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Strona, G.; Carstens, C.J.; Beck, P.S. Network analysis reveals why Xylella fastidiosa will persist in Europe. Sci. Rep. 2017, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Groom, Q.J.; Desmet, P.; Vanderhoeven, S.; Adriaens, T. The importance of open data for invasive alien species research, policy and management. Manag. Biol. Invasions 2015, 6, 119–125. [Google Scholar] [CrossRef]
- Latini, A.; Foxi, C.; Borfecchia, F.; Lentini, A.; De Cecco, L.; Iantosca, D.; Serafini, M.; Laneri, U.; Citterio, M.; Campiotti, A. Tacking the vector of Xylella fastidiosa: Geo-statistical analysis of long-term field observations on host plants influencing the distribution of Phylaenus spumarius nymphs. Environ. Sci. Pollut. Res. 2019, 26, 6503–6516. [Google Scholar] [CrossRef]
- Santoiemma, G.; Tamburini, G.; Sanna, F.; Mori, N.; Marini, L. Landscape composition predicts the distribution of Philaenus spumarius, vector of Xylella fastidiosa, in olive groves. J. Pest Sci. 2019, 92, 1101–1109. [Google Scholar] [CrossRef]
- Di Serio, F.; Bodino, N.; Cavalieri, V.; Demichelis, S.; Di Carolo, M.; Dongiovanni, C.; Fumarola, G.; Gilioli, G.; Guerrieri, E.; Picciotti, U. Collection of Data and Information on Biology and Control of Vectors of Xylella fastidiosa. EFSA Supporting Publ. 2019, 16. [Google Scholar] [CrossRef]
- Morente, M.; Cornara, D.; Plaza, M.; Durán, J.M.; Capiscol, C.; Trillo, R.; Ruiz, M.; Ruz, C.; Sanjuan, S.; Pereira, J.A.; et al. Distribution and relative abundance of insect vectors of Xylella fastidiosa in olive groves of the Iberian peninsula. Insects 2018, 9, 175. [Google Scholar] [CrossRef]
- Maggiore, G.; Semeraro, T.; Aretano, R.; De Bellis, L.; Luvisi, A. GIS analysis of land-use change in threatened landscapes by Xylella fastidiosa. Sustainability 2019, 11, 253. [Google Scholar] [CrossRef]
- Semeraro, T.; Gatto, E.; Buccolieri, R.; Vergine, M.; Gao, Z.; De Bellis, L.; Luvisi, A. Changes in Olive Urban Forests Infected by Xylella fastidiosa: Impact on Microclimate and Social Health. Int. J. Environ. Res. Public Health 2019, 16, 2642. [Google Scholar] [CrossRef]
- De Angelis, D.L.; Yurek, S. Spatially explicit modeling in ecology: A review. Ecosystems 2017, 20, 284–300. [Google Scholar] [CrossRef]
- Austin, M. Species distribution models and ecological theory: A critical assessment and some possible new approaches. Ecol. Model. 2007, 200, 1–19. [Google Scholar] [CrossRef]
- Hernández, O.G.; García, L.V. La dimensión geográfica de las invasiones biológicas en el Antropoceno: El caso de Xylella fastidiosa. Bol. Asoc. Geógrafos Esp. 2019. [Google Scholar] [CrossRef]
- Hernández, O.G.; García, L.V. Incidencia de Xylella fastidiosa en las Islas Baleares y distribución potencial en la península ibérica. Investig. Geográficas Esp 2018, 55–72. [Google Scholar] [CrossRef]
- Godefroid, M.; Cruaud, A.; Streito, J.-C.; Rasplus, J.-Y.; Rossi, J.-P. Climate change and the potential distribution of Xylella fastidiosa in Europe. BioRxiv 2018, 289876. [Google Scholar] [CrossRef]
- Hao, T.; Elith, J.; Guillera-Arroita, G.; Lahoz-Monfort, J.J. A review of evidence about use and performance of species distribution modelling ensembles like BIOMOD. Divers. Distrib. 2019, 25, 839–852. [Google Scholar] [CrossRef]
- Lieth, J.; Meyer, M.; Yeo, K.-H.; Kirkpatrick, B. Modeling cold curing of Pierce’s disease in Vitis vinifera ‘Pinot Noir’and ‘Cabernet sauvignon’grapevines in California. Phytopathology 2011, 101, 1492–1500. [Google Scholar] [CrossRef]
- Broadmeadow, S.; Watts, K.; Quine, C.; Mitchell, R. Risk Mapping of the Likelihood and Impact of a Xylella Fastidiosa Outbreak in Scotland; Plant Health Centre: Dundee, UK, 2019. [Google Scholar]
- Chapman, D.S.; White, S.M.; Hooftman, D.A.; Bullock, J.M. Inventory and Review of Quantitative Models for Spread of Plant Pests for Use in Pest Risk Assessment for the EU Territory; Wiley Online Library: Hoboken, NJ, USA, 2015. [Google Scholar]
- White, S.M.; Bullock, J.M.; Hooftman, D.A.; Chapman, D.S. Modelling the spread and control of Xylella fastidiosa in the early stages of invasion in Apulia, Italy. Biol. Invasions 2017, 19, 1825–1837. [Google Scholar] [CrossRef]
- Abboud, C.; Bonnefon, O.; Parent, E.; Soubeyrand, S. Dating and localizing an invasion from post-introduction data and a coupled reaction–diffusion–absorption model. J. Math. Biol. 2019, 79, 765–789. [Google Scholar] [CrossRef]
- Soubeyrand, S.; de Jerphanion, P.; Martin, O.; Saussac, M.; Manceau, C.; Hendrikx, P.; Lannou, C. Inferring pathogen dynamics from temporal count data: The emergence of Xylella fastidiosa in France is probably not recent. New Phytol. 2018, 219, 824–836. [Google Scholar] [CrossRef]
- White, S.; Bullock, J.; Cavers, S.; Chapman, D. Using Modelling to Investigate the Effectiveness of National Surveillance Monitoring Aimed at Detecting a Xylella Fastidiosa Outbreak in Scotland; Plant Health Centre: Dundee, UK, 2019. [Google Scholar]
- Almeida, R.; De La Fuente, L.; Koebnik, R.; Lopes, J.R.S.; Parnell, S.; Scherm, H. Addressing the new global threat of Xylella fastidiosa. Phytopathology 2019, 109, 172–174. [Google Scholar] [CrossRef] [PubMed]
- Martelli, G.P. The current status of the quick decline syndrome of olive in southern Italy. Phytoparasitica 2016, 44, 1–10. [Google Scholar] [CrossRef]
- Sicard, A.; Zeilinger, A.R.; Vanhove, M.; Schartel, T.E.; Beal, D.J.; Daugherty, M.P.; Almeida, R.P. Xylella fastidiosa: Insights into an emerging plant pathogen. Annu. Rev. Phytopathol. 2018, 56, 181–202. [Google Scholar] [CrossRef]
- Elad, Y.; Pertot, I. Climate change impacts on plant pathogens and plant diseases. J. Crop Improv. 2014, 28, 99–139. [Google Scholar] [CrossRef]
- Trebicki, P.; Finlay, K. Pests and diseases under climate change; its threat to food security. Food Secur. Clim. Chang. 2019, 229–249. [Google Scholar] [CrossRef]
- Trębicki, P.; Vandegeer, R.K.; Bosque-Pérez, N.A.; Powell, K.S.; Dader, B.; Freeman, A.J.; Yen, A.L.; Fitzgerald, G.J.; Luck, J.E. Virus infection mediates the effects of elevated CO 2 on plants and vectors. Sci. Rep. 2016, 6, 22785. [Google Scholar] [CrossRef] [PubMed]
- Kudela, V. Potential impact of climate change on geographic distribution of plant pathogenic bacteria in Central Europe. Plant Prot. Sci. 2009, 45. [Google Scholar] [CrossRef]
- Hegerl, G.C.; Zwiers, F.W.; Stott, P.A.; Kharin, V.V. Detectability of anthropogenic changes in annual temperature and precipitation extremes. J. Clim. 2004, 17, 3683–3700. [Google Scholar] [CrossRef]
- Kosmas, C.; Danalatos, N. Climate change, desertification and the Mediterranean region. In Soil Responses to Climate Change; Springer: Berlin/Heidelberg, Germany, 1994; pp. 25–38. [Google Scholar]
- Robinson, E.A.; Ryan, G.D.; Newman, J.A. A meta-analytical review of the effects of elevated CO2 on plant–arthropod interactions highlights the importance of interacting environmental and biological variables. New Phytol. 2012, 194, 321–336. [Google Scholar] [CrossRef]
- Hughes, L.; Bazzaz, F.A. Effects of elevated CO2 on five plant-aphid interactions. Entomol. Exp. Appl. 2001, 99, 87–96. [Google Scholar] [CrossRef]
- Xie, H.; Zhao, L.; Wang, W.; Wang, Z.; Ni, X.; Cai, W.; He, K. Changes in life history parameters of Rhopalosiphum maidis (Homoptera: Aphididae) under four different elevated temperature and CO2 combinations. J. Econ. Entomol. 2014, 107, 1411–1418. [Google Scholar] [CrossRef] [PubMed]
- Peñalver-Cruz, A.; Garzo, E.; Prieto-Ruiz, I.; Díaz-Carro, M.; Winters, A.; Moreno, A.; Fereres, A. Feeding behavior, life history, and virus transmission ability of Bemisia tabaci Mediterranean species (Hemiptera: Aleyrodidae) under elevated CO2. Insect Sci. 2019, 27, 558–570. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, T.; Xiao, N.; Li, J.; Chen, F. Effects of elevated CO2 on the interspecific competition between two sympatric species of Aphis gossypii and Bemisia tabaci fed on transgenic Bt cotton. Insect Sci. 2011, 18, 426–434. [Google Scholar] [CrossRef]
- Klaiber, J.; Najar-Rodriguez, A.J.; Dialer, E.; Dorn, S. Elevated carbon dioxide impairs the performance of a specialized parasitoid of an aphid host feeding on Brassica plants. Biol. Control 2013, 66, 49–55. [Google Scholar] [CrossRef]
- Overgaard, J.; Kearney, M.R.; Hoffmann, A.A. Sensitivity to thermal extremes in A ustralian Drosophila implies similar impacts of climate change on the distribution of widespread and tropical species. Glob. Chang. Biol. 2014, 20, 1738–1750. [Google Scholar] [CrossRef] [PubMed]
- Wiens, J.J.; Graham, C.H. Niche conservatism: Integrating evolution, ecology, and conservation biology. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 519–539. [Google Scholar] [CrossRef]
- Gerson, U.; Weintraub, P.G. Mites (Acari) as a factor in greenhouse management. Annu. Rev. Entomol. 2012, 57, 229–247. [Google Scholar] [CrossRef]
- Cao, L.-J.; Gao, Y.-F.; Gong, Y.-J.; Chen, J.-C.; Chen, M.; Hoffmann, A.; Wei, S.-J. Population analysis reveals genetic structure of an invasive agricultural thrips pest related to invasion of greenhouses and suitable climatic space. Evol. Appl. 2019, 12, 1868–1880. [Google Scholar] [CrossRef]
- Broennimann, O.; Treier, U.A.; Müller-Schärer, H.; Thuiller, W.; Peterson, A.; Guisan, A. Evidence of climatic niche shift during biological invasion. Ecol. Lett. 2007, 10, 701–709. [Google Scholar] [CrossRef]
- Gholam, Z.; Sadeghi, A. Management strategies for western flower thrips in vegetable greenhouses in Iran: A review. Plant Prot. Sci. 2016, 52, 87–98. [Google Scholar] [CrossRef]
- Ricciardi, A.; Blackburn, T.M.; Carlton, J.T.; Dick, J.T.A.; Hulme, P.E.; Iacarella, J.C.; Jeschke, J.M.; Liebhold, A.M.; Lockwood, J.L.; MacIsaac, H.J.; et al. Invasion Science: A Horizon Scan of Emerging Challenges and Opportunities. Trends Ecol. Evol. 2017, 32, 464–474. [Google Scholar] [CrossRef] [PubMed]
- Bullock, J.M.; Bonte, D.; Pufal, G.; da Silva Carvalho, C.; Chapman, D.S.; García, C.; García, D.; Matthysen, E.; Delgado, M.M. Human-mediated dispersal and the rewiring of spatial networks. Trends Ecol. Evol. 2018, 33, 958–970. [Google Scholar] [CrossRef] [PubMed]
- Trębicki, P.; Dáder, B.; Vassiliadis, S.; Fereres, A. Insect–plant–pathogen interactions as shaped by future climate: Effects on biology, distribution, and implications for agriculture. Insect Sci. 2017, 24, 975–989. [Google Scholar] [CrossRef] [PubMed]
- Toft, C.; Andersson, S.G. Evolutionary microbial genomics: Insights into bacterial host adaptation. Nat. Rev. Genet. 2010, 11, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Esteves, M.B.; Kleina, H.T.; Sales, T.d.M.; Oliveira, T.P.; de Lara, I.A.R.; Almeida, R.P.P.; Coletta-Filho, H.D.; Lopes, J.R.S. Transmission Efficiency of Xylella fastidiosa subsp. pauca Sequence Types by Sharpshooter Vectors after In Vitro Acquisition. Phytopathology 2018, 109, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Salzberg, S.L. Next-generation genome annotation: We still struggle to get it right. Genome Biol. 2019, 20, 92. [Google Scholar] [CrossRef]
- Kuśmirek, W.; Nowak, R. De novo assembly of bacterial genomes with repetitive DNA regions by dnaasm application. BMC Bioinform. 2018, 19, 273. [Google Scholar] [CrossRef]
- Ouyang, S.; Park, G.; Atamian, H.S.; Han, C.S.; Stajich, J.E.; Kaloshian, I.; Borkovich, K.A. MicroRNAs Suppress NB Domain Genes in Tomato That Confer Resistance to Fusarium oxysporum. PLoS Pathog. 2014, 10. [Google Scholar] [CrossRef]
- Benes, V.; Castoldi, M. Expression profiling of microRNA using real-time quantitative PCR, how to use it and what is available. Methods 2010, 50, 244–249. [Google Scholar] [CrossRef]
- Depledge, D.P.; Srinivas, K.P.; Sadaoka, T.; Bready, D.; Mori, Y.; Placantonakis, D.G.; Mohr, I.; Wilson, A.C. Direct RNA sequencing on nanopore arrays redefines the transcriptional complexity of a viral pathogen. Nat. Commun. 2019, 10, 754. [Google Scholar] [CrossRef]
- Quijada, N.M.; Rodríguez-Lázaro, D.; Eiros, J.M.; Hernández, M. TORMES: An automated pipeline for whole bacterial genome analysis. Bioinformatics 2019, 35, 4207–4212. [Google Scholar] [CrossRef]
- Genovesi, P.; Shine, C. European Strategy on Invasive Alien Species: Convention on the Conservation of European Wildlife and Habitats (Bern Convention); Council of Europe: Strasbourg, France, 2004; ISBN 92-871-5488-0. [Google Scholar]
- Gallien, L.; Münkemüller, T.; Albert, C.H.; Boulangeat, I.; Thuiller, W. Predicting potential distributions of invasive species: Where to go from here? Divers. Distrib. 2010, 16, 331–342. [Google Scholar] [CrossRef]
- Huang, C.; Asner, G.P. Applications of remote sensing to alien invasive plant studies. Sensors 2009, 9, 4869–4889. [Google Scholar] [CrossRef] [PubMed]
- Gualano, S.; Tarantino, E.; Santoro, F.; Valentini, F.; Dongiovanni, N.; D’Onghia, A.M. Analisi Assistita da Immagini Aeree ad Elevata Risoluzione Geometrica per il Riconoscimento del Complesso del Disseccamento Rapido dell’Olivo Associato al Batterio Xylella fastidiosa in Puglia. Available online: http://atti.asita.it/ASITA2014/Pdf/097.pdf (accessed on 28 May 2020).
- D’onghia, A.M. CIHEAM/IAMB innovative tools for early surveillance and detection of Xylella fastidiosa. In Xylella Fastidiosa & the Olive Quick Decline Syndrome (OQDS) A Serious Worldwide Challenge for the Safeguard of Olive Trees; D’Onghia, A.M., Brunel, S., Valentini, F., Eds.; CIHEAM: Paris, France, 2017; Volume 172. [Google Scholar]
- D’onghia, A.M.; Brunel, S.; Valentini, F. Xylella Fastidiosa & the Olive Quick Decline Syndrome (OQDS) A Serious Worldwide Challenge for the Safeguard of Olive Trees; CIHEAM: Paris, France, 2017; ISBN 978-2-85352-570-1. [Google Scholar]
- Gippet, J.M.; Liebhold, A.M.; Fenn-Moltu, G.; Bertelsmeier, C. Human-mediated dispersal in insects. Curr. Opin. Insect Sci. 2019, 35, 96–102. [Google Scholar] [CrossRef]
- Almeida, R.P.P. Emerging plant disease epidemics: Biological research is key but not enough. PLoS Biol. 2018, 16, e2007020. [Google Scholar] [CrossRef]





{kind=link}
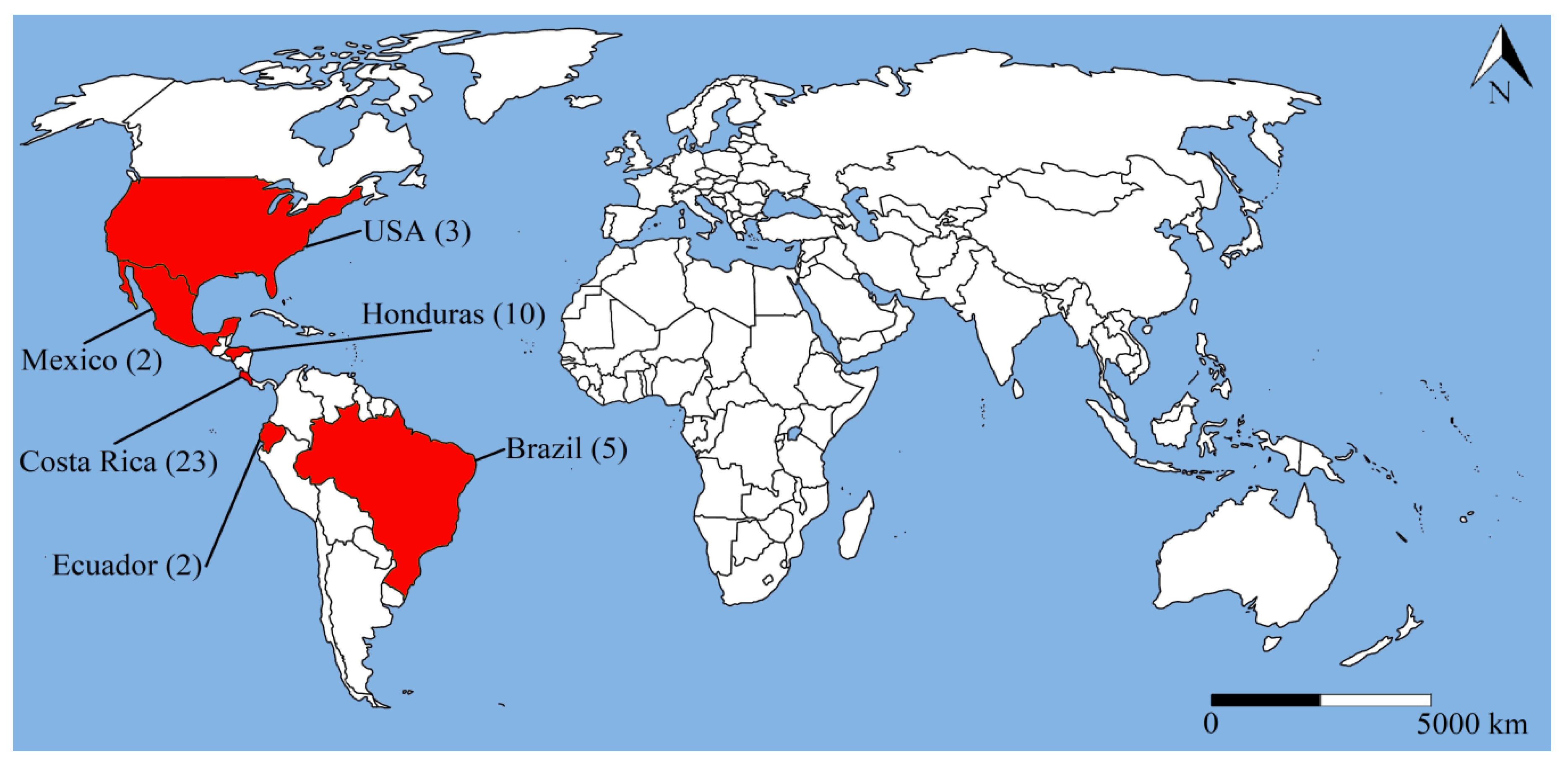{kind=link}
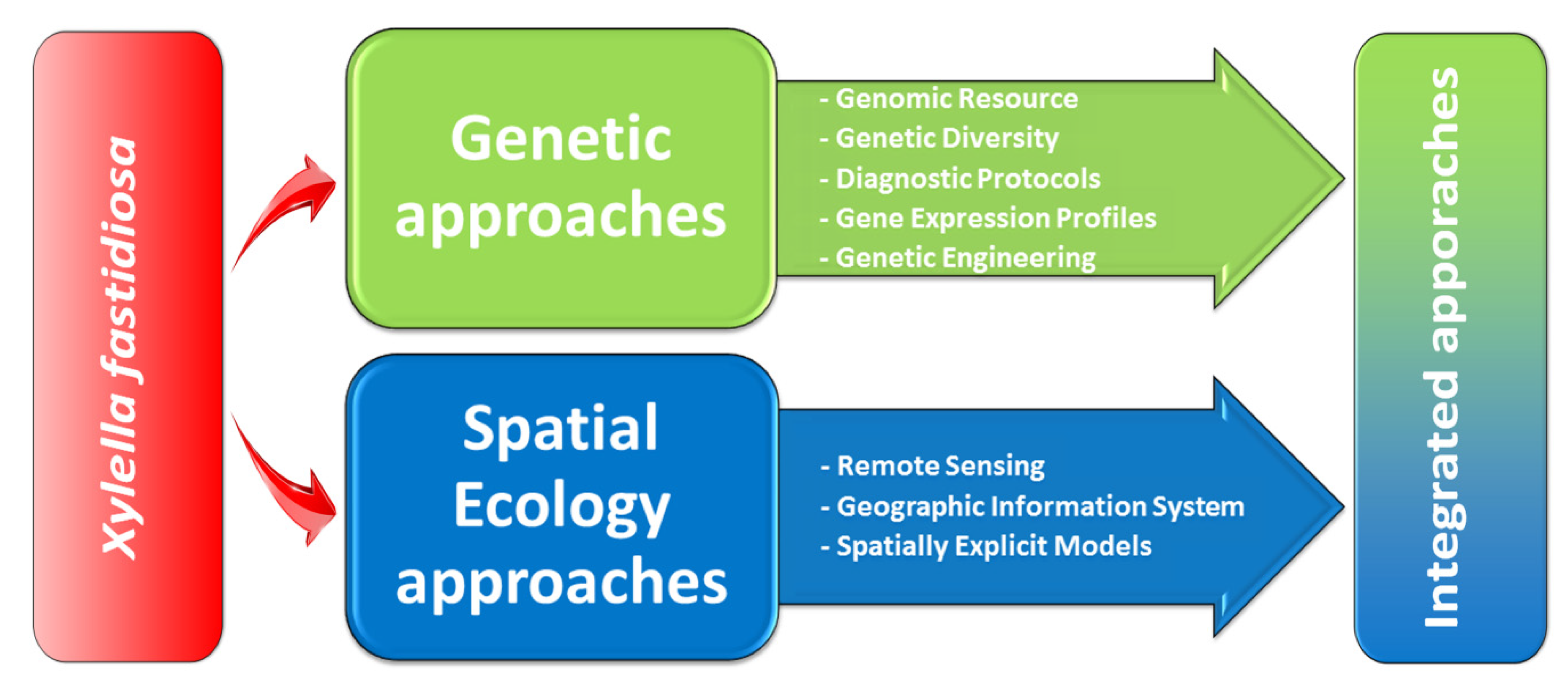{kind=link}
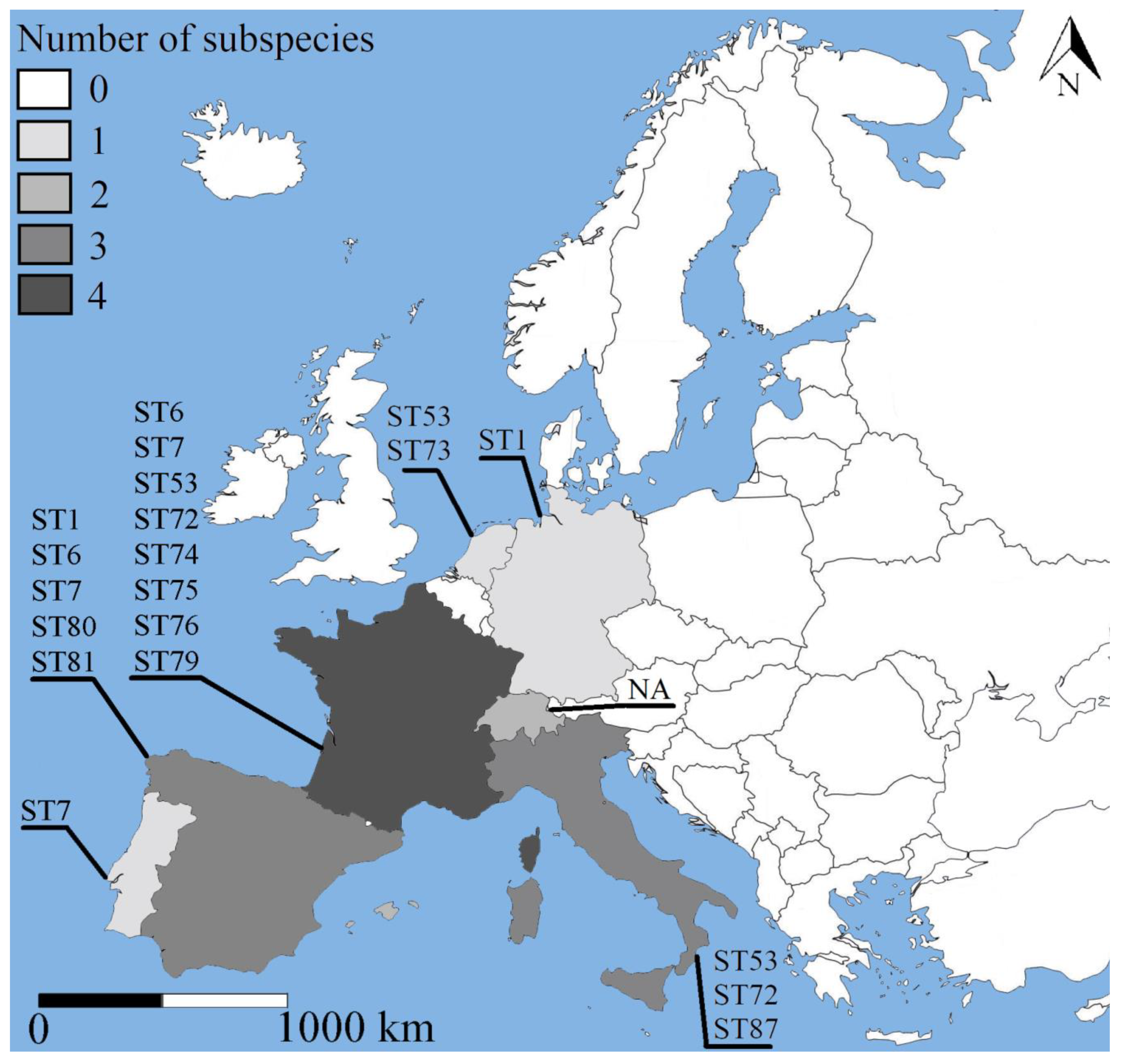{kind=link}
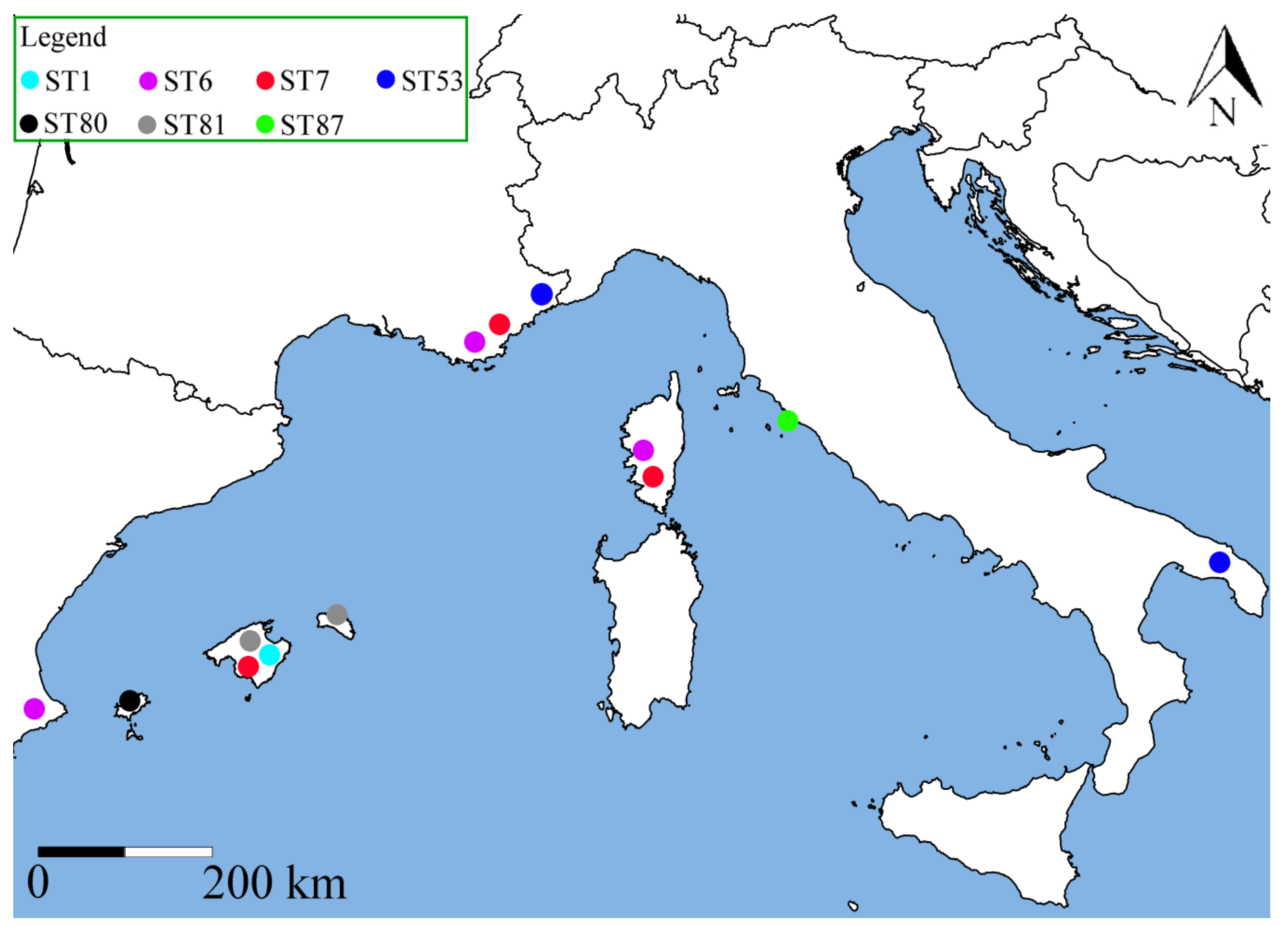{kind=link}
| Region | Source | Link |
|---|---|---|
| Brazil/USA | Xf Comparative Genome Database | https://www.xylella.lncc.br |
| Europe | XF-ACTORS | https://www.xfactorsproject.eu/project/genome-sequences-of-eu-strains-of-xylella-fastidiosa/ |
| UK | BRIGIT | https://www.jic.ac.uk/brigit |
| UK | SapFeedersHub | http://sapfeedershub.jic.ac.uk/ |
| Worldwide | Xf MLST Database | https://pubmlst.org/xfastidiosa/ |
| Worldwide | CBOL | https://www.boldsystems.org |
| Worldwide | NCBI | https://www.ncbi.nlm.nih.gov/genome/genomes/173? |
| Region | Source | Available data | Link |
|---|---|---|---|
| Europe | EFSA | GPS coordinates, taxonomic classification | https://www.efsa.europa.eu/en/microstrategy/xylella |
| Europe | EUROPHYT | Exporting country, commodity, plant species, harmful organism, number of intercepts | https://ec.europa.eu/food/plant/plant_health_biosecurity/europhyt_en |
| Europe | EPPO | Country, state, invasion status | https://gd.eppo.int/taxon/XYLEFA/distribution |
| France | ANSES | Country, region, municipality, taxonomic classification | https://shiny-public.anses.fr/Xylella_fastidiosa/ |
| Italy | SIT.Puglia | GPS coordinates, taxonomic classification, genetic test results of screened plants and insects | https://sit.puglia.it/portal/portale_gestione_agricoltura/Cartografie |
| Spain | MAPA | GPS coordinates, plant host, taxonomic classification | https://www.mapa.gob.es/es/agricultura/temas/sanidad-vegetal/xylella-fastidiosa/ |
| Spain | GOIB | GPS coordinates, plant host, taxonomic classification | https://www.caib.es/sites/sanitatvegetal/es/inicio-1542/?campa=yes |
| Worldwide | GBIF | GPS coordinates, taxonomic classification | https://www.gbif.org/species/3222355 |
| Predictor | Source | Link |
|---|---|---|
| Climate | WorldClim | https://www.worldclim.org/data/worldclim21.html |
| Climate | CHELSA | https://chelsa-climate.org/ |
| Elevation | EEA | https://www.eea.europa.eu/data-and-maps/data/digital-elevation-model-of-europe |
| Elevation | WorldClim | https://www.worldclim.org/data/worldclim21.html |
| Land cover | Copernicus | https://land.copernicus.eu/pan-european/corine-land-cover |
| Land cover | ESA | http://maps.elie.ucl.ac.be/CCI/viewer |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raffini, F.; Bertorelle, G.; Biello, R.; D’Urso, G.; Russo, D.; Bosso, L. From Nucleotides to Satellite Imagery: Approaches to Identify and Manage the Invasive Pathogen Xylella fastidiosa and Its Insect Vectors in Europe. Sustainability 2020, 12, 4508. https://doi.org/10.3390/su12114508
Raffini F, Bertorelle G, Biello R, D’Urso G, Russo D, Bosso L. From Nucleotides to Satellite Imagery: Approaches to Identify and Manage the Invasive Pathogen Xylella fastidiosa and Its Insect Vectors in Europe. Sustainability. 2020; 12(11):4508. https://doi.org/10.3390/su12114508
Chicago/Turabian StyleRaffini, Francesca, Giorgio Bertorelle, Roberto Biello, Guido D’Urso, Danilo Russo, and Luciano Bosso. 2020. "From Nucleotides to Satellite Imagery: Approaches to Identify and Manage the Invasive Pathogen Xylella fastidiosa and Its Insect Vectors in Europe" Sustainability 12, no. 11: 4508. https://doi.org/10.3390/su12114508
APA StyleRaffini, F., Bertorelle, G., Biello, R., D’Urso, G., Russo, D., & Bosso, L. (2020). From Nucleotides to Satellite Imagery: Approaches to Identify and Manage the Invasive Pathogen Xylella fastidiosa and Its Insect Vectors in Europe. Sustainability, 12(11), 4508. https://doi.org/10.3390/su12114508