Biodiversity Impact of Green Roofs and Constructed Wetlands as Progressive Eco-Technologies in Urban Areas


Abstract

1. Introduction
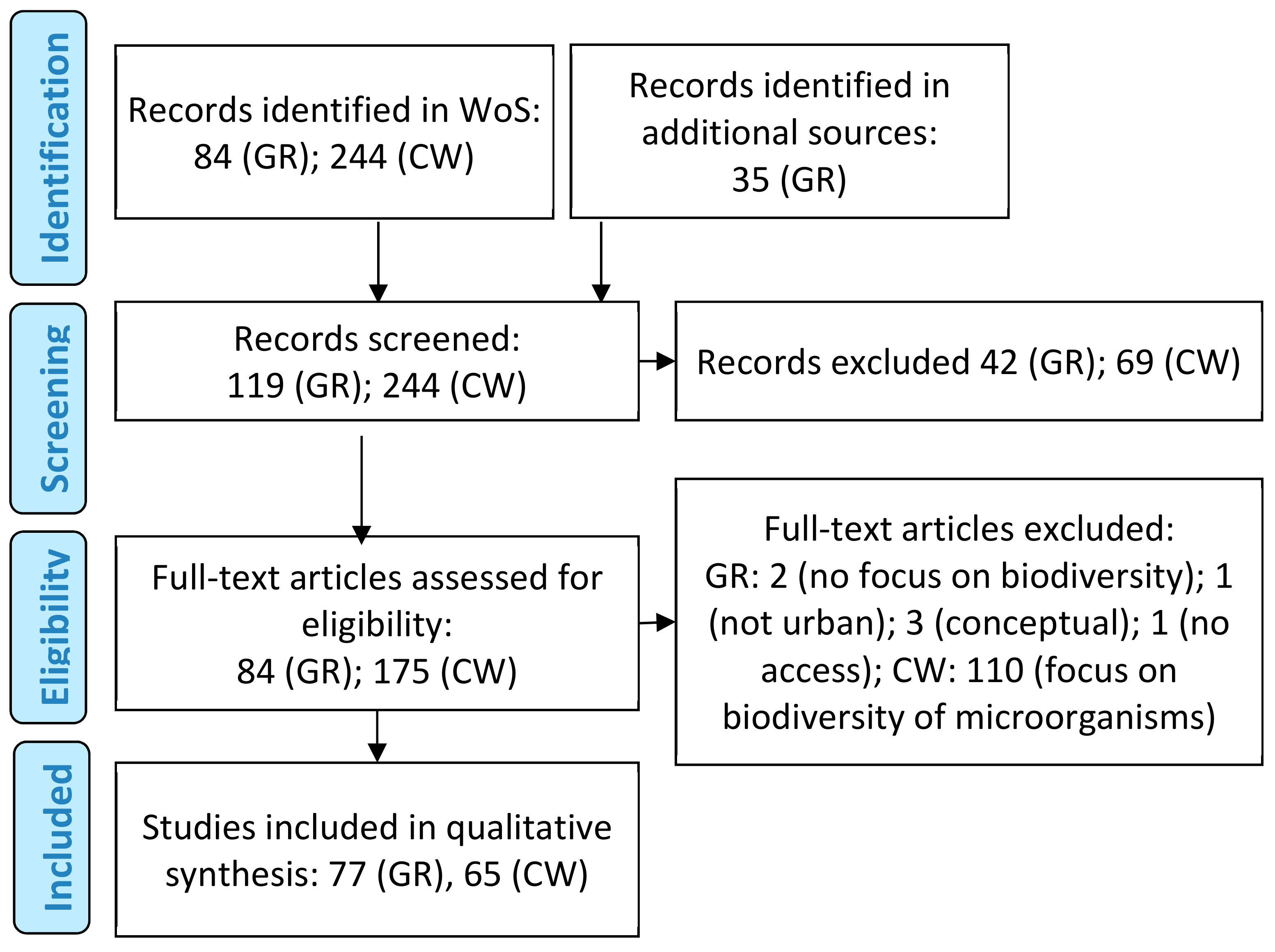
2. Materials and Methods
- Whether a study had been conducted in an urban area
- The country and city in which a study had taken place
- The type of roof (green/brown/conventional roof, intensive/extensive, experimental) or of constructed wetland (laboratory scale vs. full scale)
- The number of roofs studied
- The length of the study period
- The focal species group(s)
- Whether cultivated and/or spontaneously occurring species were in focus
- Species richness and abundance reported in a study
- Trends in species richness and abundance
- Other reported diversity measures and their trends, such as functional diversity indices, numbers of rare and endangered or native and exotic species
3. Results and Discussion
3.1. Green Roofs
3.1.1. Patterns in Green Roof Research
3.1.2. Trends in Green Roof Species Richness
3.1.3. Trends in Green Roof Species Abundance
3.1.4. Trends in Green Roof Functional Diversity and Patterns in Functional Traits
3.1.5. Patterns of Rare and Endangered Species on Green Roofs
3.1.6. Patterns of Native vs. Exotic Species on Green Roofs
3.2. Constructed Wetlands
3.2.1. Patterns in Constructed Wetland Research
- Effects of root characteristics of wetland plants on nutrient decomposition [108]
- Viruses in wetland ecosystems, with the result that little is known about this topic [109]
- Native herbaceous plant species with potential use in phytoremediation of heavy metals with a focus on wetlands [110]
- Borrow pits (ponds created during highway construction) - not wetlands for water treatment purposes [111]
- Wastewater reuse for fiber crop cultivation as a strategy to mitigate desertification [115]
- Assessment of pathogenic bacteria in treated greywater and irrigated soils [116]
- Impact of trout aquaculture on water quality and farm effluent treatment options [117]
3.2.2. Impacts of Constructed Wetlands on Biodiversity
4. Conclusions
4.1. Opportunities to Increase the Biodiversity of Green Roofs
4.2. Opportunities to Increase Biodiversity in Constructed Wetlands
- Flat shores
- Barrier-free design of shores
- Vegetation consisting of shore plants and submerged plants
- Varied layout of the wetland’s surroundings, including hiding places
- Temporal drying-out of some parts of a wetland
- Minimal disturbance by human activities
4.3. Combining Green Roofs and Constructed Wetlands in Urban Settings
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- United Nations Department of Economic and Social Affairs Population Division. World Urbanization Prospects: The 2018 Revision, Online ed.; United Nations Department of Economic and Social Affairs Population Division: New York, NY, USA, 2018. [Google Scholar]
- Barton, H. Land use planning and health and well-being. Land Use Policy 2009, 26, S115–S123. [Google Scholar] [CrossRef]
- Kowarik, I. Novel urban ecosystems, biodiversity, and conservation. Environ. Pollut. 2011, 159, 1974–1983. [Google Scholar] [CrossRef] [PubMed]
- Natural Capital Germany—TEEB DE. Ecosystem Services in the City—Protecting Health and Enhancing Quality of Life. Summary for Deci-Sion-Makers; Technical University of Berlin, Helmholtz Centre for Environmental Research—UFZ: Berlin, Germany; Leipzig, Germany, 2017. [Google Scholar]
- European Union. The EU Biodiversity Strategy to 2020; Publications Office of the European Union: Luxembourg, 2011. [Google Scholar]
- European Commission Green Infrastructure (GI)—Enhancing Europe’s Natural Capital. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:52013DC0249 (accessed on 20 May 2019).
- TEEB. The Economics of Ecosystems and Biodiversity: Mainstreaming the Economics of Nature: A Synthesis of the Approach, Conclusions and Recommendations of TEEB. 2010. Available online: http://www.teebweb.org/publication/mainstreaming-the-economics-of-nature-a-synthesis-of-the-approach-conclusions-and-recommendations-of-teeb/ (accessed on 18 October 2019).
- Die Bundesregierung. Perspektiven für Deutschland. Unsere Strategie für Eine Nachhaltige Entwicklung; Presse-Und Informationsamt der Bundesregierung: Berlin, Germany, 2002. [Google Scholar]
- Böhm, J.; Böhme, C.; Bunzel, A.; Landua, D.; Kühnau, C.; Reinke, M. Urbanes Grün in der Doppelten Innenentwicklung; Bundesamt für Naturschutz: Bonn, Germany, 2016; Volume 444. [Google Scholar]
- Fischer, L.K.; Honold, J.; Cvejić, R.; Delshammar, T.; Hilbert, S.; Lafortezza, R.; Nastran, M.; Nielsen, A.B.; Pintar, M.; van der Jagt, A.P.N.; et al. Beyond green: Broad support for biodiversity in multicultural European cities. Glob. Environ. Chang. 2018, 49, 35–45. [Google Scholar] [CrossRef]
- BMUB—Federal Ministry for the Environment, Nature Conservation, B. and N.S.; BfN—Federal Agency for Nature Conservation. 2015 Nature Awareness Study. Population Survey on Nature and Biological Diversity; BMUB and BfN: Berlin, Germany, 2016. [Google Scholar]
- Haaland, C.; van den Bosch, C.K. Challenges and strategies for urban green-space planning in cities undergoing densification: A review. Urban For. Urban Green. 2015, 14, 760–771. [Google Scholar] [CrossRef]
- Climate Service Center. Machbarkeitsstudie Starkregenrisiko 2050: Abschlussbericht Kooperationsprojekt des Gesamt-Verbandes der Deutschen Versicherungswirtschaft e. V. (GDV) und des Climate Service Centers (CSC); Climate Service Center: Hamburg, Germany, 2012. [Google Scholar]
- Kotowski, A.; Kazmierczak, B.; Nowakowska, M. Analysis of Rainwater Sewerage Systems Overloads on Rakowiec Estate in Wroclaw Caused by Cli-mate Changes. Rocznik Ochrona Srodowiska 2014, 16, 608–626. [Google Scholar]
- Teklehaimanot, G.Z.; Kamika, I.; Coetzee, M.A.A.; Momba, M.N.B. Population Growth and Its Impact on the Design Capacity and Performance of the Wastewater Treatment Plants in Sedibeng and Soshanguve, South Africa. Environ. Manag. 2015, 56, 984–997. [Google Scholar] [CrossRef]
- Zehnsdorf, A.; Blumberg, M.; Müller, R.A. Helophyte mats (wetland roofs) with high evapotranspiration rates as a tool for decentralised rain-water management-process stability improved by simultaneous greywater treatment. Water Sci. Technol. Water Supply 2019, in press. [Google Scholar] [CrossRef]
- Nivala, J.; Zehnsdorf, A.; van Afferden, M.; Müller, R.A. Green infrastructure for increased resource efficiency in urban water management. In Urban transformations—Sustainable Urban Development through Resource Efficiency, Quality of Life and Resilience; Kabisch, S., Koch, F., Gawel, E., Haase, A., Knapp, S., Krellenberg, K., Nivala, J., Zehnsdorf, A., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 133–143. [Google Scholar]
- Sailor, D.; Elley, T.B.; Gibson, M. Exploring the building energy impacts of green roof design decisions—A modeling study of buildings in four distinct climates. J. Build. Phys. 2011, 35, 372–391. [Google Scholar] [CrossRef]
- Foustalierakia, M.; Assimakopoulosa, M.N.; Santamourisa, M.; Pangalouca, H. Energy performance of a medium scale green roof system installed on a commercial building using numerical and experimental data recorded during the cold period of the year. Energy Build. 2017, 135, 33–38. [Google Scholar] [CrossRef]
- Baik, J.-J.; Kwak, K.-H.; Park, S.-B.; Ryu, Y.-H. Effects of building roof greening on air quality in street canyons. Atmos. Environ. 2012, 61, 48–55. [Google Scholar] [CrossRef]
- Carter, T.; Jackson, C.R. Vegetated roofs for stormwater management at multiple spatial scales. Landsc. Urban Plan. 2007, 80, 84–94. [Google Scholar] [CrossRef]
- Berndtsson, J.C.; Bengtsson, L.; Jinno, K. Runoff water quality from intensive and extensive vegetated roofs. Ecol. Eng. 2009, 35, 369–380. [Google Scholar] [CrossRef]
- Williams, N.S.G.; Lundholm, J.; Scott Macivor, J. Do green roofs help urban biodiversity conservation? J. Appl. Ecol. 2014, 51, 1643–1649. [Google Scholar] [CrossRef]
- Rosenzweig, M.L. Green roofs: New ecosystems to defend species diversity. Isr. J. Ecol. Evol. 2016, 62, 7–14. [Google Scholar] [CrossRef]
- Vymazal, J. Constructed wetlands for wastewater treatment: Five decades of experience. Environ. Sci. Technol. 2011, 45, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, J.B.; de Oliveira, P.J.A.; Árpád Boncz, M.; Loureiro Paulo, P. A modified constructed wetland system for greywater treatmen. Desalin. Water Treat. 2017, 91, 31–39. [Google Scholar] [CrossRef]
- Abtew, W. Evapotranspiration measurements and modeling for three wetland systems in South Florida. J. Am. Water Resour. Assoc. 1996, 32, 465–473. [Google Scholar] [CrossRef]
- Otterpohl, R.; Braun, U.; Oldenburg, M. Innovative technologies for decentralised water-, wastewater and biowaste management in urban and peri-urban areas. Water Sci. Technol. 2004, 48, 23–32. [Google Scholar] [CrossRef]
- Steele, M.K.; Heffernan, J.B.; Bettez, N.; Cavender-Bares, J.; Groffman, P.M.; Grove, J.M.; Hall, S.; Hobbie, S.E.; Larson, K.; Morse, J.L.; et al. Convergent Surface Water Distributions in U.S. Cities. Ecosystems 2014, 17, 685–697. [Google Scholar] [CrossRef]
- Mao, D.; Wang, Z.; Wu, J.; Wu, B.; Zeng, Y.; Song, K.; Yi, K.; Luo, L. China’s wetlands loss to urban expansion. L. Degrad. Dev. 2018, 29, 2644–2657. [Google Scholar] [CrossRef]
- Knapp, S.; Kühn, I.; Stolle, J.; Klotz, S. Changes in the functional composition of a Central European urban flora over three centuries. Perspect. Plant Ecol. Evol. Syst. 2010, 12, 235–244. [Google Scholar] [CrossRef]
- Pickering, C.; Byrne, J. The benefits of publishing systematic quantitative literature reviews for PhD candidates and other early-career researchers. High. Educ. Res. Dev. 2014, 33, 534–548. [Google Scholar] [CrossRef]
- Schweizerischer Ingenieur-und Architektenverein. Begrünung von Dächern; Schweizerischer Ingenieur-und Architektenverein: Zürich, Switzerland, 2013. [Google Scholar]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. PRISMA 2009 Flow Diagram. The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar]
- Aronson, M.F.J.; La Sorte, F.A.; Nilon, C.H.; Katti, M.; Goddard, M.A.; Lepczyk, C.A.; Warren, P.S.; Williams, N.S.G.; Cilliers, S.; Clarkson, B.; et al. A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers. Proc. R. Soc. B Biol. Sci. USA 2014, 281, 20133330. [Google Scholar] [CrossRef] [PubMed]
- Christoffel, R.A.; Lepczyk, C.A. Representation of herpetofauna in wildlife research journals. J. Wildl. Manag. 2012, 76, 661–669. [Google Scholar] [CrossRef]
- Titley, M.A.; Snaddon, J.L.; Turner, E.C. Scientific research on animal biodiversity is systematically biased towards vertebrates and temperate regions. PLoS ONE 2017, 12, e0189577. [Google Scholar] [CrossRef]
- Donaldson, M.R.; Burnett, N.J.; Braun, D.C.; Suski, C.D.; Hinch, S.G.; Cooke, S.J.; Kerr, J.T. Taxonomic bias and international biodiversity conservation research. Facets 2016, 1, 105–113. [Google Scholar] [CrossRef]
- Rumble, H.; Finch, P.; Gange, A.C. Green roof soil organisms: Anthropogenic assemblages or natural communities? Appl. Soil Ecol. 2018, 126, 11–20. [Google Scholar] [CrossRef]
- Vasl, A.; Shalom, H.; Kadas, G.J.; Blaustein, L. Sedum—Annual plant interactions on green roofs: Facilitation, competition and exclusion. Ecol. Eng. 2017, 108, 318–329. [Google Scholar] [CrossRef]
- Molineux, C.J.; Gange, A.C.; Connop, S.P.; Newport, D.J. Using recycled aggregates in green roof substrates for plant diversity. Ecol. Eng. 2015, 82, 596–604. [Google Scholar] [CrossRef]
- McKinney, M.L.; Sisco, N.D. Systematic Variation in Roof Spontaneous Vegetation: Residential “Low Rise” Versus Commercial “High Rise” Buildings. Urban Nat. 2018, SI, 73–88. [Google Scholar]
- Ksiazek-Mikenas, K.; Herrmann, J.; Menke, S.B.; Köhler, M. If You Build It, Will They Come? Plant and Arthropod Diversity on Urban Green Roofs Over Time. Urban Nat. 2018, 1, 52–72. [Google Scholar]
- Catalano, C.; Marcenò, C.; Laudicina, V.A.; Guarino, R. Thirty years unmanaged green roofs: Ecological research and design implications. Landsc. Urban Plan. 2016, 149, 11–19. [Google Scholar] [CrossRef]
- Klein, P.M.; Coffman, R. Establishment and performance of an experimental green roof under extreme climatic conditions. Sci. Total Environ. 2015, 512, 82–93. [Google Scholar] [CrossRef]
- Schrader, S.; Böning, M. Soil formation on green roofs and its contribution to urban biodiversity with emphasis on Collembolans. Pedobiologia (Jena) 2006, 50, 347–356. [Google Scholar] [CrossRef]
- McGuire, K.L.; Payne, S.G.; Palmer, M.I.; Gillikin, C.M.; Keefe, D.; Kim, S.J.; Gedallovich, S.M.; Discenza, J.; Rangamannar, R.; Koshner, J.A.; et al. Digging the New York City Skyline: Soil Fungal Communities in Green Roofs and City Parks. PLoS ONE 2013, 8, e58020. [Google Scholar] [CrossRef]
- Eakin, C.J.; Campa, H.; Linden, D.W.; Roloff, G.J.; Bradley Rowe, D.; Westphal, J. Avian response to green roofs in urban landscapes in the Midwestern USA. Wildl. Soc. Bull. 2015, 39, 574–582. [Google Scholar] [CrossRef]
- Tonietto, R.; Fant, J.; Ascher, J.; Ellis, K.; Larkin, D. A comparison of bee communities of Chicago green roofs, parks and prairies. Landsc. Urban Plan. 2011, 103, 102–108. [Google Scholar] [CrossRef]
- Madre, F.; Vergnes, A.; Machon, N.; Clergeau, P. A comparison of 3 types of green roof as habitats for arthropods. Ecol. Eng. 2013, 57, 109–117. [Google Scholar] [CrossRef]
- Blank, L.; Vasl, A.; Schindler, B.Y.; Kadas, G.J.; Blaustein, L. Horizontal and vertical island biogeography of arthropods on green roofs: A review. Urban Ecosyst. 2017, 20, 911–917. [Google Scholar] [CrossRef]
- Thuring, C.; Grant, G. The biodiversity of temperate extensive green roofs—A review of research and practice. Isr. J. Ecol. Evol. 2016, 62, 44–57. [Google Scholar] [CrossRef]
- Joimel, S.; Grard, B.; Auclerc, A.; Hedde, M.; Le Doaré, N.; Salmon, S.; Chenu, C. Are Collembola “flying” onto green roofs? Ecol. Eng. 2018, 111, 117–124. [Google Scholar] [CrossRef]
- Nagase, A.; Yamada, Y.; Aoki, T.; Nomura, M. Developing Biodiverse Green Roofs for Japan: Arthropod and Colonizer Plant Diversity on Harappa and Biotope Roofs. Urban Nat. 2018, 1, 16–38. [Google Scholar]
- Deng, H.; Jim, C.Y. Spontaneous plant colonization and bird visits of tropical extensive green roof. Urban Ecosyst. 2017, 20, 337–352. [Google Scholar] [CrossRef]
- Kandas, G. Rare Invertebrates Colonizing Green Roofs in London. Urban Habitat 2006, 4, 66–86. [Google Scholar]
- Partridge, D.R.; Clark, J.A. Urban green roofs provide habitat for migrating and breeding birds and their arthropod prey. PLoS ONE 2018, 13, e0202298. [Google Scholar] [CrossRef] [PubMed]
- Parkins, K.L.; Clark, J.A. Green roofs provide habitat for urban bats. Glob. Ecol. Conserv. 2015, 4, 349–357. [Google Scholar] [CrossRef]
- Washburn, B.E.; Swearingin, R.M.; Pullins, C.K.; Rice, M.E. Composition and Diversity of Avian Communities Using a New Urban Habitat: Green Roofs. Environ. Manag. 2016, 57, 1230–1239. [Google Scholar] [CrossRef]
- Pearce, H.; Walters, C.L. Do Green Roofs Provide Habitat for Bats in Urban Areas? Acta Chiropterol. 2012, 14, 469–478. [Google Scholar] [CrossRef]
- Heim, A.; Lundholm, J. The effects of substrate depth heterogeneity on plant species coexistence on an extensive green roof. Ecol. Eng. 2014, 68, 184–188. [Google Scholar] [CrossRef]
- Van Mechelen, C.; Van Meerbeek, K.; Dutoit, T.; Hermy, M. Functional diversity as a framework for novel ecosystem design: The example of extensive green roofs. Landsc. Urban Plan. 2015, 136, 165–173. [Google Scholar] [CrossRef]
- Walker, E.A.; Lundholm, J.T. Designed habitat heterogeneity on green roofs increases seedling survival but not plant species diversity. J. Appl. Ecol. 2018, 55, 694–704. [Google Scholar] [CrossRef]
- Dunnett, N.; Nagase, A.; Hallam, A. The dynamics of planted and colonising species on a green roof over six growing seasons 2001–2006: Influence of substrate depth. Urban Ecosyst. 2008, 11, 373–384. [Google Scholar] [CrossRef]
- Brown, C.; Lundholm, J. Microclimate and substrate depth influence green roof plant community dynamics. Landsc. Urban Plan. 2015, 143, 134–142. [Google Scholar] [CrossRef]
- Braaker, S.; Obrist, M.K.; Ghazoul, J.; Moretti, M. Habitat connectivity and local conditions shape taxonomic and functional diversity of arthropods on green roofs. J. Anim. Ecol. 2017, 86, 521–531. [Google Scholar] [CrossRef]
- Kyrö, K.; Brenneisen, S.; Kotze, D.J.; Szallies, A.; Gerner, M.; Lehvävirta, S. Local habitat characteristics have a stronger effect than the surrounding urban landscape on beetle communities on green roofs. Urban For. Urban Green. 2018, 29, 122–130. [Google Scholar] [CrossRef]
- Molineux, C.J.; Gange, A.C.; Newport, D.J. Using soil microbial inoculations to enhance substrate performance on extensive green roofs. Sci. Total Environ. 2017, 580, 846–856. [Google Scholar] [CrossRef]
- Nagase, A.; Dunnett, N.; Choi, M.S. Investigation of weed phenology in an establishing semi-extensive green roof. Ecol. Eng. 2013, 58, 156–164. [Google Scholar] [CrossRef]
- MacIvor, J.S.; Margolis, L.; Puncher, C.L.; Carver Matthews, B.J. Decoupling factors affecting plant diversity and cover on extensive green roofs. J. Environ. Manag. 2013, 130, 297–305. [Google Scholar] [CrossRef]
- Aloisio, J.M.; Palmer, M.I.; Giampieri, M.A.; Tuininga, A.R.; Lewis, J.D. Spatially dependent biotic and abiotic factors drive survivorship and physical structure of green roof vegetation. Ecol. Appl. 2017, 27, 297–308. [Google Scholar] [CrossRef]
- Nash, C.; Clough, J.; Gedge, D.; Lindsay, R.; Newport, D.; Ciupala, M.A.; Connop, S. Initial insights on the biodiversity potential of biosolar roofs: A London Olympic Park green roof case study. Isr. J. Ecol. Evol. 2016, 62, 74–87. [Google Scholar] [CrossRef]
- Schindler, B.Y.; Blank, L.; Levy, S.; Kadas, G.; Pearlmutter, D.; Blaustein, L. Integration of photovoltaic panels and green roofs: Review and predictions of effects on electricity production and plant communities. Isr. J. Ecol. Evol. 2016, 62, 68–73. [Google Scholar] [CrossRef]
- Grant, G. Extensive green roofs in London. Urban Habitats 2006, 4, 51–65. [Google Scholar]
- Benvenuti, S. Wildflower green roofs for urban landscaping, ecological sustainability and biodiversity. Landsc. Urban Plan. 2014, 124, 151–161. [Google Scholar] [CrossRef]
- Rumble, H.; Gange, A.C. Soil microarthropod community dynamics in extensive green roofs. Ecol. Eng. 2013, 57, 197–204. [Google Scholar] [CrossRef]
- Braaker, S.; Ghazoul, J.; Obrist, M.K.; Moretti, M. Habitat connectivity shapes urban arthropod communities: The key role of green roofs. Ecology 2014, 95, 1010–1021. [Google Scholar] [CrossRef]
- Molineux, C.J.; Gange, A.C.; Connop, S.P.; Newport, D.J. Are microbial communities in green roof substrates comparable to those in post-industrial sites?—A preliminary study. Urban Ecosyst. 2015, 18, 1245–1260. [Google Scholar] [CrossRef]
- MacIvor, J.S.; Lundholm, J. Insect species composition and diversity on intensive green roofs and adjacent level-ground habitats. Urban Ecosyst. 2011, 14, 225–241. [Google Scholar] [CrossRef]
- Ksiazek, K.; Fant, J.; Skogen, K. An assessment of pollen limitation on Chicago green roofs. Landsc. Urban Plan. 2012, 107, 401–408. [Google Scholar] [CrossRef]
- Schmauck, S. Dach-und Fassadenbegrünung - neue Lebensräume im Siedlungsbereich. Fakten, Argumente und Empfehlungen. BfN-Skripten 2019, 538, 1–64. [Google Scholar]
- Baumann, N. Ground-Nesting Birds on Green Roofs in Switzerland: Preliminary Observations. Urban Habitats 2006, 4, 37–50. [Google Scholar]
- Baumann, N. Ecological compensation on the roof: Ground-nesting birds and vegetation. In World Green Roof Congress; World Green Roof Congress Proceedings: London, UK, 2010. [Google Scholar]
- Cook-Patton, S.C.; Bauerle, T.L. Potential benefits of plant diversity on vegetated roofs: A literature review. J. Environ. Manag. 2012, 106, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Lundholm, J.; Heim, A.; Tran, S.; Smith, T. Leaf and Life History Traits Predict Plant Growth in a Green Roof Ecosystem. PLoS ONE 2014, 9, e101395.s. [Google Scholar] [CrossRef] [PubMed]
- Getter, K.L.; Rowe, D.B. Media depth influences Sedum green roof establishment. Urban Ecosyst. 2008, 11, 361–372. [Google Scholar] [CrossRef]
- Yalcinalp, E.; Ozveren, S.; Meral, A.; Pulatkan, M.; Akbulut, S. Habitat effect on urban roof vegetation. Sustainability 2017, 9, 1985. [Google Scholar] [CrossRef]
- Köhler, M.; Poll, P.H. Long-term performance of selected old Berlin greenroofs in comparison to younger extensive greenroofs in Berlin. Ecol. Eng. 2010, 36, 722–729. [Google Scholar] [CrossRef]
- Van Mechelen, C.; Dutoit, T.; Hermy, M. Vegetation development on different extensive green roof types in a Mediterranean and temperate maritime climate. Ecol. Eng. 2015, 82, 571–582. [Google Scholar] [CrossRef]
- Bates, A.J.; Sadler, J.P.; Greswell, R.B.; Mackay, R. Effects of varying organic matter content on the development of green roof vegetation: A six year experiment. Ecol. Eng. 2015, 82, 301–310. [Google Scholar] [CrossRef]
- Bates, A.J.; Sadler, J.P.; Greswell, R.B.; Mackay, R. Effects of recycled aggregate growth substrate on green roof vegetation development: A six year experiment. Landsc. Urban Plan. 2015, 135, 22–31. [Google Scholar] [CrossRef]
- Bornkamm, R. Vegetation und vegetations-entwicklung auf Kiesdächern. Vegetatio 1961, 10, 1–24. [Google Scholar] [CrossRef]
- Bates, A.J.; Sadler, J.P.; Mackay, R. Vegetation development over four years on two green roofs in the UK. Urban For. Urban Green. 2013, 12, 98–108. [Google Scholar] [CrossRef]
- Lundholm, J.; Tran, S.; Gebert, L. Plant functional traits predict green roof ecosystem services. Environ. Sci. Technol. 2015, 49, 2366–2374. [Google Scholar] [CrossRef] [PubMed]
- Villéger, S.; Mason, N.W.H.; Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef] [PubMed]
- Garnier, E.; Cortez, J.; Billès, G.; Navas, M.L.; Roumet, C.; Debussche, M.; Laurent, G.; Blanchard, A.; Aubry, D.; Bellmann, A.; et al. Plant functional markers capture ecosystem properties during secondary succession. Ecology 2004, 85, 2630–2637. [Google Scholar] [CrossRef]
- Čeplová, N.; Lososová, Z.; Zelený, D.; Chytrý, M.; Danihelka, J.; Fajmon, K.; Láníková, D.; Preislerová, Z.; Řehořek, V.; Tíchý, L. Phylogenetic diversity of central-European urban plant communities: Effects of alien species and habitat types. Preslia 2015, 87, 1–16. [Google Scholar]
- Holt, R.D. Green roofs may cast shadows. Isr. J. Ecol. Evol. 2016, 62, 15–22. [Google Scholar] [CrossRef]
- Ricotta, C.; Moretti, M. CWM and Rao’s quadratic diversity: A unified framework for functional ecology. Oecologia 2011, 167, 181–188. [Google Scholar] [CrossRef]
- Madre, F.; Vergnes, A.; Machon, N.; Clergeau, P. Green roofs as habitats for wild plant species in urban landscapes: First insights from a large-scale sampling. Landsc. Urban Plan. 2014, 122, 100–107. [Google Scholar] [CrossRef]
- Kinlock, N.L.; Schindler, B.Y.; Gurevitch, J. Biological invasions in the context of green roofs. Isr. J. Ecol. Evol. 2016, 62, 32–43. [Google Scholar] [CrossRef]
- Oberndorfer, E.; Lundholm, J.; Bass, B.; Coffman, R.R.; Doshi, H.; Dunnett, N.; Gaffin, S.; Köhler, M.; Liu, K.K.Y.; Rowe, B. Green Roofs as Urban Ecosystems: Ecological Structures, Functions, and Services. Bioscience 2007, 57, 823–833. [Google Scholar] [CrossRef]
- Kratschmer, S.; Kriechbaum, M.; Pachinger, B. Buzzing on top: Linking wild bee diversity, abundance and traits with green roof qualities. Urban Ecosyst. 2018, 21, 429–446. [Google Scholar] [CrossRef]
- MacIvor, J.S.; Ruttan, A.; Salehi, B. Exotics on exotics: Pollen analysis of urban bees visiting Sedum on a green roof. Urban Ecosyst. 2015, 18, 419–430. [Google Scholar] [CrossRef]
- Pétremand, G.; Chittaro, Y.; Braaker, S.; Brenneisen, S.; Gerner, M.; Obrist, M.K.; Rochefort, S.; Szallies, A.; Moretti, M. Ground beetle (Coleoptera: Carabidae) communities on green roofs in Switzerland: Synthesis and perspectives. Urban Ecosyst. 2018, 21, 119–132. [Google Scholar] [CrossRef]
- Starry, O.; Gonsalves, S.; Ksiazek-Mikenas, K.; MacIvor, J.S.; Gardner, M.; Szallies, A.; Brenneisen, S. A Global Comparison of Beetle Community Composition on Green Roofs and the Potential for Homogenization. Urban Nat. 2018, 1, 1–15. [Google Scholar]
- Kohler, M. Long-term vegetation research on two extensive green roofs in Berlin. Urban Habitats 2006, 4, 3–26. [Google Scholar]
- Cheng, X.Y.; Chen, W.Y.; Gu, B.H.; Liu, X.C.; Chen, F.; Chen, Z.H.; Zhou, X.Y.; Li, Y.X.; Huang, H.; Chen, Y.J. Morphology, ecology, and contaminant removal efficiency of eight wetland plants with differing root systems. Hydrobiologia 2009, 623, 77–85. [Google Scholar] [CrossRef]
- Jackson, E.F.; Jackson, C.R. Viruses in wetland ecosystems. Freshw. Biol. 2008, 53, 1214–1227. [Google Scholar] [CrossRef]
- Oyuela Leguizamo, M.A.; Fernández Gómez, W.D.; Sarmiento, M.C.G. Native herbaceous plant species with potential use in phytoremediation of heavy metals, spotlight on wetlands—A review. Chemosphere 2017, 168, 1230–1247. [Google Scholar] [CrossRef]
- Kuczynski, E.C.; Paszkowski, C.A.; Gingras, B.A. Horned grebe habitat use of constructed wetlands in Alberta, Canada. J. Wildl. Manag. 2012, 76, 1694–1702. [Google Scholar] [CrossRef]
- Denton, R.D.; Richter, S.C. Amphibian communities in natural and constructed ridge top wetlands with implications for wetland construction. J. Wildl. Manag. 2013, 77, 886–896. [Google Scholar] [CrossRef]
- Hartwig, T.S.; Kiviat, E. Microhabitat Association of Blanding’s Turtles in Natural and Constructed Wetlands in Southeastern New York. J. Wildl. Manag. 2007, 71, 576–582. [Google Scholar] [CrossRef]
- Balcombe, C.K.; Anderson, J.T.; Fortney, R.H.; Rentch, J.S.; Grafton, W.N.; Kordek, W.S. A comparison of plant communities in mitigation and reference wetlands in the mid-appalachians. Wetlands 2006, 25, 130–142. [Google Scholar] [CrossRef]
- Barbosa, B.; Costa, J.; Fernando, A.L.; Papazoglou, E.G. Wastewater reuse for fiber crops cultivation as a strategy to mitigate desertification. Ind. Crops Prod. 2015, 68, 17–23. [Google Scholar] [CrossRef]
- Benami, M.; Gross, A.; Herzberg, M.; Orlofsky, E.; Vonshak, A.; Gillor, O. Assessment of pathogenic bacteria in treated graywater and irrigated soils. Sci. Total Environ. 2013, 458–460, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Sindilariu, P.-D.; Reiter, R.; Wedekind, H. Impact of trout aquaculture on water quality and farm effluent treatment options. Aquat. Living Resour. 2009, 22, 93–103. [Google Scholar] [CrossRef]
- Yoon, C.G. Wise use of paddy rice fields to partially compensate for the loss of natural wetlands. Paddy Water Environ. 2009, 7, 357–366. [Google Scholar] [CrossRef]
- Lacki, M.J.; Hummer, J.W.; Webster, H.J. Effect of Reclamation Technique on Mammal Communitites Inhabiting Wetlands on Mined Lands in East-Central Ohio. Ohio J. Sci. 1991, 92, 154–158. [Google Scholar]
- De Martis, G.; Mulas, B.; Malavasi, V.; Marignani, M. Can Artificial Ecosystems Enhance Local Biodiversity? The Case of a Constructed Wetland in a Mediterranean Urban Context. Environ. Manag. 2016, 57, 1088–1097. [Google Scholar] [CrossRef]
- Hsu, C.B.; Hsieh, H.L.; Yang, L.; Wu, S.H.; Chang, J.S.; Hsiao, S.C.; Su, H.C.; Yeh, C.H.; Ho, Y.S.; Lin, H.J. Biodiversity of constructed wetlands for wastewater treatment. Ecol. Eng. 2011, 37, 1533–1545. [Google Scholar] [CrossRef]
- Van Loon, E.E.; Bos, D.; van Hellenberg Hubar, C.J.; Ydenberg, R.C. A historical perspective on the effects of trapping and controlling the muskrat (Ondatra zibethicus) in the Netherlands. Pest Manag. Sci. 2017, 73, 305–312. [Google Scholar] [CrossRef]
- Kadlec, R.H.; Pries, J.; Mustard, H. Muskrats (Ondatra zibethicus) in treatment wetlands. Ecol. Eng. 2007, 29, 143–153. [Google Scholar] [CrossRef]
- Greenhorn, J.E.; Sadowski, C.; Holden, J.; Bowman, J. Coastal Wetlands Connected to Lake Ontario Have Reduced Muskrat (Ondatra zibethicus) Abundance. Wetlands 2017, 37, 339–349. [Google Scholar] [CrossRef]
- Orłowski, G. Factors affecting the use of waste-stabilization ponds by birds: A case study of conservation implications of a sewage farm in Europe. Ecol. Eng. 2013, 61, 436–445. [Google Scholar] [CrossRef]
- Scarton, F. Long-term trend of the waterbird community breeding in a heavily man-modified coastal lagoon: The case of the important bird area “Lagoon of Venice” . J. Coast. Conserv. 2017, 21, 35–45. [Google Scholar] [CrossRef]
- Lin, J.L.; Tu, Y.T.; Chiang, P.C.; Chen, S.H.; Kao, C.M. Using aerated gravel-packed contact bed and constructed wetland system for polluted river water purification: A case study in Taiwan. J. Hydrol. 2015, 525, 400–408. [Google Scholar] [CrossRef]
- Strand, J.A.; Weisner, S.E.B. Effects of wetland construction on nitrogen transport and species richness in the agricultural landscape-Experiences from Sweden. Ecol. Eng. 2013, 56, 14–25. [Google Scholar] [CrossRef]
- Scheffers, B.R.; Paszkowski, C.A. Amphibian use of urban stormwater wetlands: The role of natural habitat features. Landsc. Urban Plan. 2013, 113, 139–149. [Google Scholar] [CrossRef]
- Drayer, A.N.; Richter, S.C. Physical wetland characteristics influence amphibian community composition differently in constructed wetlands and natural wetlands. Ecol. Eng. 2016, 93, 166–174. [Google Scholar] [CrossRef]
- Furman, B.L.S.; Scheffers, B.R.; Taylor, M.; Davis, C.; Paszkowski, C.A. Limited genetic structure in a wood frog (Lithobates sylvaticus) population in an urban landscape inhabiting natural and constructed wetlands. Conserv. Genet. 2016, 17, 19–30. [Google Scholar] [CrossRef]
- Mulkeen, C.J.; Gibson-Brabazon, S.; Carlin, C.; Williams, C.D.; Healy, M.G.; Mackey, P.; Gormally, M.J. Habitat suitability assessment of constructed wetlands for the smooth newt (Lissotriton vulgaris [Linnaeus, 1758]): A comparison with natural wetlands. Ecol. Eng. 2017, 106, 532–540. [Google Scholar] [CrossRef]
- Shulse, C.D.; Semlitsch, R.D.; Trauth, K.M.; Williams, A.D. Influences of design and landscape placement parameters on amphibian abundance in constructed wetlands. Wetlands 2010, 30, 915–928. [Google Scholar] [CrossRef]
- Walton, W.E.; Workman, P.D. Effect of marsh design on the abundance of mosquitoes in experimental constructed wetlands in Southern California. J. Am. Mosq. Control Assoc. 1998, 14, 95–107. [Google Scholar] [PubMed]
- Walton, W.E. Design and management of free water surface constructed wetlands to minimize mosquito production. Wetl. Ecol. Manag. 2012, 20, 173–195. [Google Scholar] [CrossRef]
- Molnár, Á.; Csabai, Z.; Tóthmérész, B. Influence of Flooding and Vegetation Patterns on Aquatic Beetle Diversity in a Constructed Wetland Complex. Wetlands 2009, 29, 1214–1223. [Google Scholar] [CrossRef]
- Becerra-Jurado, G.; Foster, G.; Harrington, R.; Kelly-Quinn, M. Integrated constructed wetlands: Hotspots for freshwater coleopteran diversity in the landscape of Ireland. Biol. Environ. 2014, 114B, 271–279. [Google Scholar] [CrossRef]
- Steinly, B.A. Primary consumer and detritivore communities (Diptera: Ephydridae) in newly restored and constructed wetlands. Proc. Entomol. Soc. Washingt. USA 2004, 106, 460–471. [Google Scholar]
- Becerra Jurado, G.; Johnson, J.; Feeley, H.; Harrington, R.; Kelly-Quinn, M. The potential of integrated constructed wetlands (ICWs) to enhance macroinvertebrate diversity in agricultural landscapes. Wetlands 2010, 30, 393–404. [Google Scholar] [CrossRef]
- Becerra-Jurado, G.; Harrington, R.; Kelly-Quinn, M. A review of the potential of surface flow constructed wetlands to enhance macroinvertebrate diversity in agricultural landscapes with particular reference to Integrated Constructed Wetlands (ICWs). Hydrobiologia 2012, 692, 121–130. [Google Scholar] [CrossRef]
- Holtmann, L.; Juchem, M.; Brüggeshemke, J.; Möhlmeyer, A.; Fartmann, T. Stormwater ponds promote dragonfly (Odonata) species richness and density in urban areas. Ecol. Eng. 2018, 118, 1–11. [Google Scholar] [CrossRef]
- Giordano, R.; Weber, E.; Darby, B.J.; Soto-Adames, F.N.; Murray, R.E.; Drizo, A. Invertebrates Associated With a Horizontal-Flow, Subsurface Constructed Wetland in a Northern Climate. Environ. Entomol. 2014, 43, 283–290. [Google Scholar] [CrossRef]
- Ouattara, J.M.P.; Coulibaly, L.; Tiho, S.; Gourène, G. Comparison of macrofauna communities in sediments of the beds of vertical flow constructed wetlands planted with Panicum maximum (Jacq.) treating domestic wastewater. Ecol. Eng. 2009, 35, 1237–1242. [Google Scholar] [CrossRef]
- Ouattara, J.M.P.; Coulibaly, L.; Tiho, S.; Ouattara, A.; Gourène, G. Panicum maximum (Jacq.) density effect upon macrofauna structure in sediments of pilot-scale vertical flow constructed wetlands treating domestic wastewater. Ecol. Eng. 2011, 37, 217–223. [Google Scholar] [CrossRef]
- Pedescoll, A.; Rodríguez, L.; Sarañana, A.A.; Hijosa-Valsero, M.; Bécares, E. Microfaunal community in horizontal constructed wetlands with different design configurations. Ecol. Eng. 2016, 91, 16–23. [Google Scholar] [CrossRef]
- Spieles, D.J.; Horn, J.D. Macroinvertebrate community structure in created wetlands of different successional stage. Aquat. Ecosyst. Heal. Manag. 2009, 12, 320–329. [Google Scholar] [CrossRef]
- Zehnsdorf, A.; Scherber, A.; Schmidt, S.; Ebersbach, A.T.; Bernhard, K.; Jentzsch, D.; Schlenker, U. Waste Water Treatment with Reeds in a Mobile Root Space Treatment Plant. Chem. Ing. Tech. 2018, 90, 333–339. [Google Scholar] [CrossRef]
- Barczak, T.; Bennewicz, J.; Kaminski, P. Parasitoids (Hymenoptera: Braconidae, aphidiinae) of the mealy plum aphid Hyalopterus pruni (geoffr.) on common reed (Phragmites australis) in different types of habitat in poland. Arch. Biol. Sci. 2013, 65, 71–79. [Google Scholar]
- Basky, Z. Predators and Parasites of Hyalopterus-pruni and Hyalopterus-amygdali populations living on Peach, Plum and Reed. Acta Phytopathol. Acad. Sci. Hung. 1982, 17, 311–316. [Google Scholar]
- Mann, G. Rückhalt, Abflussverzögerung, Speicherung. Greenbuild. Mag. 2015, 6/15, 3. [Google Scholar]
- Lisecke, H.-J. Das Retentionsvermögen von Dachbegrünungen. Wasserspeicherfähigkeit, Wasserrückhaltung, Abflussverzögerung und Abflussbeiwerte unter besonderer Berücksichtigung von Extensivbegrünungen. Stadt Grün 1998, 47, 46–53. [Google Scholar]
- Deutschewitz, K.; Lausch, A.; Kühn, I.; Klotz, S. Native and alien plant species richness in relation to spatial heterogeneity on a regional scale in Germany. Glob. Ecol. Biogeogr. 2003, 12, 299–311. [Google Scholar] [CrossRef]
- Vanuytrecht, E.; Van Mechelen, C.; Van Meerbeek, K.; Willems, P.; Hermy, M.; Raes, D. Runoff and vegetation stress of green roofs under different climate change scenarios. Landsc. Urban Plan. 2014, 122, 68–77. [Google Scholar] [CrossRef]
- Brenneisen, S. Space for urban wildlife: Designing green roofs as habitats in Switzerland. Urban Habitats 2006, 4, 27–36. [Google Scholar]
- Catalano, C.; Laudicina, V.A.; Badalucco, L.; Guarino, R. Some European green roof norms and guidelines through the lens of biodiversity: Do ecoregions and plant traits also matter? Ecol. Eng. 2018, 115, 15–26. [Google Scholar] [CrossRef]
- Dunnett, N. Ruderal green roofs. In Green Roof Ecosystems; Sutton, R.K., Ed.; Springer: Cham, Switzerland, 2015; pp. 233–255. [Google Scholar]
- Hirschfelder, A.; Zucchi, H. Zur Besiedelung begrünter Gebäudedächer durch Carabiden—Ein Beitrag zur Stadtökologie. Zeitschrift für Ökologie und Naturschutz 1992, 1, 59–66. [Google Scholar]
- Buch, C.; Jagel, A. Schmetterlingswiese, Bienenschmaus und Hummelmagnet-Insektenrettung aus der Samentüte? Veröffentlichungen Bochumer Botanischen Vereins 2019, 11, 9–24. [Google Scholar]
- Hauck, T.E.; Weisser, W.W. AAD—Animal-aided design; Technische Universität München: München, Germany; Universität Kassel: Kassel, Germany, 2016. [Google Scholar]
- Roy, M.C.; Foote, L.; Ciborowski, J.J.H. Vegetation community composition in wetlands created following oil sand mining in Alberta, Canada. J. Environ. Manag. 2016, 172, 18–28. [Google Scholar] [CrossRef]
- Crowe, A.U.; Plant, A.L.; Kermode, A.R. Effects of an industrial effluent on plant colonization and on the germination and post-germinative growth of seeds of terrestrial and aquatic plant species. Environ. Pollut. 2002, 117, 179–189. [Google Scholar] [CrossRef]
- Liu, J.; Wang, J.; Zhao, C.; Hay, A.G.; Xie, H.; Zhan, J. Triclosan removal in wetlands constructed with different aquatic plants. Appl. Microbiol. Biotechnol. 2016, 100, 1459–1467. [Google Scholar] [CrossRef]
- Zhang, C.B.; Wang, J.; Liu, W.L.; Zhu, S.X.; Ge, H.L.; Chang, S.X.; Chang, J.; Ge, Y. Effects of plant diversity on microbial biomass and community metabolic profiles in a full-scale constructed wetland. Ecol. Eng. 2010, 36, 62–68. [Google Scholar] [CrossRef]
- Zhang, C.B.; Ke, S.S.; Wang, J.; Ge, Y.; Chang, S.X.; Zhu, S.X.; Chang, J. Responses of microbial activity and community metabolic profiles to plant functional group diversity in a full-scale constructed wetland. Geoderma 2011, 160, 503–508. [Google Scholar] [CrossRef]
- Zhang, C.B.; Liu, W.L.; Han, W.J.; Guan, M.; Wang, J.; Liu, S.Y.; Ge, Y.; Chang, J. Responses of Dissimilatory Nitrate Reduction to Ammonium and Denitrification to Plant Presence, Plant Species and Species Richness in Simulated Vertical Flow Constructed Wetlands. Wetlands 2017, 37, 109–122. [Google Scholar] [CrossRef]
- Zhu, S.; Huang, X.; Ho, S.H.; Wang, L.; Yang, J. Effect of plant species compositions on performance of lab-scale constructed wetland through investigating photosynthesis and microbial communities. Bioresour. Technol. 2017, 229, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.B.; Liu, W.L.; Wang, J.; Chen, T.; Yuan, Q.Q.; Huang, C.C.; Ge, Y.; Chang, S.X.; Chang, J. Plant functional group richness-affected microbial community structure and function in a full-scale constructed wetland. Ecol. Eng. 2011, 37, 1360–1368. [Google Scholar] [CrossRef]
- Brister, J.S.; Nelson, A.D. Vegetative ecology of natural and constructed wetlands along the Leon River in Comanche County, Texas. Tex. J. Sci. 2012, 64, 103–128. [Google Scholar]
- Mitsch, W.J.; Zhang, L.; Waletzko, E.; Bernal, B. Validation of the ecosystem services of created wetlands: Two decades of plant succession, nutrient retention, and carbon sequestration in experimental riverine marshes. Ecol. Eng. 2014, 72, 11–24. [Google Scholar] [CrossRef]
- White, S.A.; Taylor, M.D.; Damrel, D.Z. Floral Colonization of a Free-Water Surface Constructed Wetland System in Grady County, Georgia. Castanea 2012, 77, 159–171. [Google Scholar] [CrossRef]
- Pier, B.M.; Dresser, B.R.; Lee, J.J.; Boylen, C.W.; Nierzwicki-Bauer, S.A. Ecological Analysis Before and After Planting in a Constructed Wetland in the Adirondacks. Wetlands 2015, 35, 611–624. [Google Scholar] [CrossRef]
- Kearney, M.A.; Fickbohm, S.; Zhu, W. Loss of plant biodiversity over a seven-year period in two constructed wetlands in central New York. Environ. Manag. 2013, 51, 1067–1076. [Google Scholar] [CrossRef]
- Bae, S.-H.; Lee, S.-D. Construction and Management Plan of Constructed Wetland for Promoting Biodiversity. J. People Plants Environ. 2018, 21, 185–202. [Google Scholar] [CrossRef]
- Leck, M.A.; Leck, C.F. Vascular plants of a Delaware River tidal freshwater wetland and adjacent terrestrial areas: Seed bank and vegetation comparisons of reference and constructed marshes and annotated species list 1. J. Torrey Bot. Soc. 2005, 132, 323–354. [Google Scholar] [CrossRef]
- Du, Y.; Pan, K.; Yu, C.; Luo, B.; Gu, W.; Sun, H.; Min, Y.; Liu, D.; Geng, Y.; Han, W.; et al. Plant diversity decreases net global warming potential integrating multiple functions in microcosms of constructed wetlands. J. Clean. Prod. 2018, 184, 718–726. [Google Scholar] [CrossRef]
- Hansson, L.A.; Brönmark, C.; Nilsson, P.A.; Åbjörnsson, K. Conflicting demands on wetland ecosystem services: Nutrient retention, biodiversity or both? Freshw. Biol. 2005, 50, 705–714. [Google Scholar] [CrossRef]
- Tixier, G.; Lafont, M.; Grapentine, L.; Rochfort, Q.; Marsalek, J. Ecological risk assessment of urban stormwater ponds: Literature review and proposal of a new conceptual approach providing ecological quality goals and the associated bioassessment tools. Ecol. Indic. 2011, 11, 1497–1506. [Google Scholar] [CrossRef]
- Bullock, J.M.; Aronson, J.; Newton, A.C.; Pywell, R.F.; Rey-Benayas, J.M. Restoration of ecosystem services and biodiversity: Conflicts and opportunities. Trends Ecol. Evol. 2011, 26, 541–549. [Google Scholar] [CrossRef]
- Clifford, C.C.; Heffernan, J.B. Artificial aquatic ecosystems. Water 2018, 10, 1096. [Google Scholar] [CrossRef]
- Zhang, K.; Chui, T.F.M. Linking hydrological and bioecological benefits of green infrastructures across spatial scales—A literature review. Sci. Total Environ. 2019, 646, 1219–1231. [Google Scholar] [CrossRef]
- Hultberg, M.; Prade, T.; Bodin, H.; Vidakovic, A.; Asp, H. Adding benefit to wetlands-Valorization of harvested common reed through mushroom production. Sci. Total Environ. 2018, 637–638, 1395–1399. [Google Scholar] [CrossRef]
- Zapater-Pereyra, M.; Lavrnić, S.; van Dien, F.; van Bruggen, J.J.A.; Lens, P.N.L. Constructed wetroofs: A novel approach for the treatment and reuse of domestic wastewater. Ecol. Eng. 2016, 94, 545–554. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Roof Characteristics | Species Richness | Species Abundance | Functional Diversity |
|---|---|---|---|
| Age | → (8) | → (13) | N.A. |
| Area | ↑ (1) | ↑ (1) | N.A. |
| Connectivity | ↗ (2) | ↗ (3) | ↗ (1) |
| Conventionala | ↘ (5) | ↓ (2) | N.A. |
| Extensiveb | ↘ (5) | N.A. | ↘ (2) |
| Height | ↗ (3) | ↓ (3) | N.A. |
| Irrigation | ↑ (1) | ↑ (1) | N.A. |
| Mowing | → (1) | → (1) | N.A. |
| Mycorrhiza | ↑ (1) | N.A. | N.A. |
| Plant cover | ↑ (2) | → (3) | N.A. |
| Plant diversity | ↑ (2) | ↑ (1) | → (1) |
| Roof compared to ground | ↘ (7) | ↘ (7) | - (1) |
| Shade | ↑ (3) | ↑ (2) | N.A. |
| Structural/ habitat diversity/ heterogeneity | ↑ (5) | ↑ (2) | N.A. |
| Substrate depth | ↗ (11) | ↗ (7) | - (1) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knapp, S.; Schmauck, S.; Zehnsdorf, A. Biodiversity Impact of Green Roofs and Constructed Wetlands as Progressive Eco-Technologies in Urban Areas. Sustainability 2019, 11, 5846. https://doi.org/10.3390/su11205846
Knapp S, Schmauck S, Zehnsdorf A. Biodiversity Impact of Green Roofs and Constructed Wetlands as Progressive Eco-Technologies in Urban Areas. Sustainability. 2019; 11(20):5846. https://doi.org/10.3390/su11205846
Chicago/Turabian StyleKnapp, Sonja, Sebastian Schmauck, and Andreas Zehnsdorf. 2019. "Biodiversity Impact of Green Roofs and Constructed Wetlands as Progressive Eco-Technologies in Urban Areas" Sustainability 11, no. 20: 5846. https://doi.org/10.3390/su11205846
APA StyleKnapp, S., Schmauck, S., & Zehnsdorf, A. (2019). Biodiversity Impact of Green Roofs and Constructed Wetlands as Progressive Eco-Technologies in Urban Areas. Sustainability, 11(20), 5846. https://doi.org/10.3390/su11205846