Genetic Diversity and Structure of the Portuguese Pear (Pyrus communis L.) Germplasm

 , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and DNA Extraction
2.2. PCR Amplification and Fragments Sizing
2.3. Data Analysis
3. Results
3.1. Overall Genetic Diversity
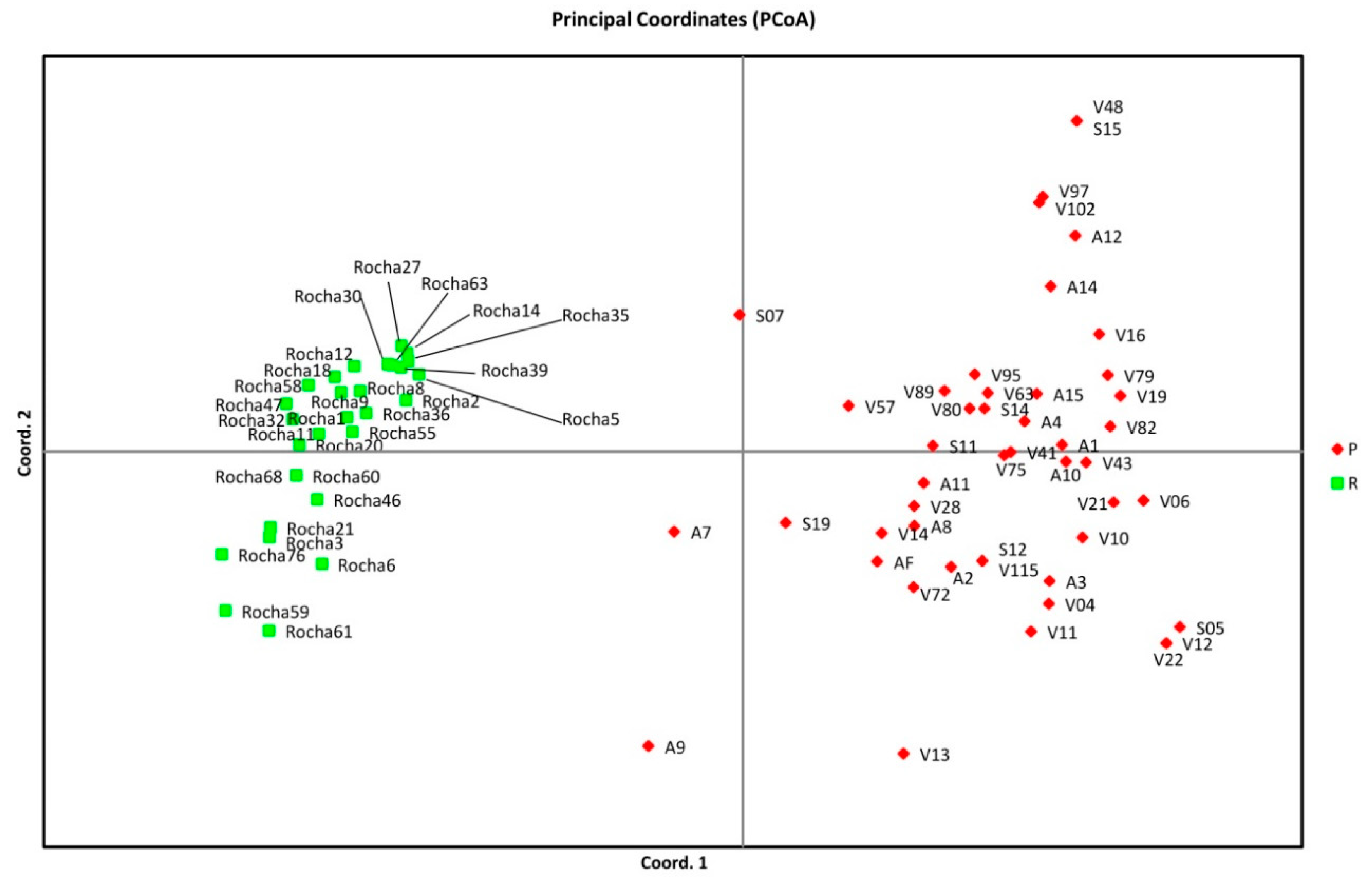
3.2. Genetic Relationships among Pear Genotypes
3.3. Genetic Structure of Pears
3.4. Polyploidy
4. Discussion
4.1. Overall Genetic Diversity
4.2. Genetic Relationships among Pear Genotypes
4.3. Genetic Structure
4.4. Polyploidy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- EUROSTAT. 2017. Available online: https://ec.europa.eu/eurostat/statisticsexplained/index.php/Agricultural_production_-_orchards (accessed on 1 April 2019).
- Bettencourt, E. Status of Malus/Pyrus germplasm collections in Portugal. In Report of a Working Group on Malus/Pyrus, Second Meeting, Dresden-Pillnitz, Germany, 2–4 May 2002; Maggioni, L., Fischer, M., Lateur, M., Lamont, E.J., Lipman, E., Eds.; International Plant Genetic Resources Institute: Rome, Italy, 2004; pp. 46–50. ISBN 92-9043-611-5. [Google Scholar]
- Mata, G.J. A Pereira Rocha. Subsídio para o estudo das variedades de pereiras portuguesas. In Actas do 2º Congresso Nacional de Pomologia; Ministério da Agricultura: Lisboa, Portugal, 1932; Volume I. [Google Scholar]
- Soares, J.; Silva, A.; Alexandre, J. O Livro da Pereira Rocha, 1st ed.; Associação Nacional dos Produtores de Pereira Rocha: Cadaval, Portugal, 2001; p. 184. [Google Scholar]
- Amaral, J.D. As Pomóideas. Manual Enciclopédico do AgricultorPortuguês, 1st ed.; Gazeta das Aldeias: Porto, Portugal, 1959. [Google Scholar]
- Associação Nacional dos Produtores de Pera Rocha. Available online: http://perarocha.pt/ (accessed on 1 April 2019).
- Laurentin, H. Data analysis for molecular characterization of plant genetic resources. Genet. Resour. Crop Evol. 2009, 56, 277–292. [Google Scholar] [CrossRef]
- van Treuren, R.; Kemp, H.; Ernsting, G.; Jongejans, B.; Houtman, H.; Visser, L. Microsatellite genotyping of apple (Malus×domestica Borkh.) genetic resources in the Netherlands: Application in collection management and variety identification. Genet. Resour. Crop Evol. 2010, 57, 853–865. [Google Scholar] [CrossRef]
- Urrestarazu, J.; Denancé, C.; Ravon, E.; Guyader, A.; Guisnel, R.; Feugey, L.; Poncet, C.; Lateur, M.; Houben, P.; Ordidge, M.; et al. Analysis of the genetic diversity and structure across a wide range of germplasm reveals prominent gene flow in apple at the European level. BMC Plant Biol. 2016, 16, 130. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.; Tanabe, K. Reconsideration on the origin of cultivated pears native to East Asia. Acta Hortic. 2004, 634, 175–182. [Google Scholar] [CrossRef]
- Zheng, X.; Cai, D.; Potter, D.; Postman, J.; Liu, J.; Teng, Y. Phylogeny and evolutionary histories of Pyrus, L. revealed by phylogenetic trees and networks based on data from multiple DNA sequences. Mol. Phylogenet. Evol. 2014, 80, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Urrestarazu, J.; Royo, J.B.; Santesteban, L.G.; Miranda, C. Evaluating the influence of the microsatellite marker set on the genetic structure inferred in Pyrus communis L. PLoS ONE 2015, 10, e0138417. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.M.; Mota, M.; Monte-Corvo, L.; Goulão, L.; Silva, D.M. Molecular typing of Pyrus based on RAPD markers. Sci. Hortic. 1999, 79, 163–174. [Google Scholar] [CrossRef]
- Monte-Corvo, L.; Cabrita, L.; Oliveira, C.; Leitão, J. Assessment of genetic relationships among Pyrus species and cultivars using AFLP and RAPD markers. Genet. Resour. Crop Evol. 2000, 47, 257–265. [Google Scholar] [CrossRef]
- Queiroz, A.; Assunção, A.; Ramadas, I.; Viegas, W.; Veloso, M.M. Molecular characterization of Portuguese pear landraces (Pyrus communis L.) using SSR markers. Sci. Hortic. 2015, 183, 72–76. [Google Scholar] [CrossRef]
- Almandanim, M.C.; Baleiras-Couto, M.M.; Pereira, H.S.; Carneiro, L.C.; Fevereiro, P.; Eiras-Dias, J.E.; Morais-Cecílio, L.; Viegas, W.; Veloso, M.M. Genetic diversity of the grapevine (Vitis vinifera L.) cultivars most utilized for wine production in Portugal. Vitis 2007, 46, 116–119. [Google Scholar]
- Kalia, R.K.; Rai, M.K.; Kalia, S.; Singh, R.; Dhawan, A.K. Microsatellite markers: An overview of the recent progress in plants. Euphytica 2011, 177, 309–334. [Google Scholar] [CrossRef]
- Santos, A.R.F.; Ramos-Cabrer, A.M.; Díaz-Hernandez, M.B.; Pereira-Lorenzo, S. Genetic variability and diversification process in local pear cultivars from northwestern Spain using microsatellites. Tree Genet. Genomes 2011, 7, 1041–1056. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhong, C.; Liu, Y.; Zhang, Q.; Sun, X.; Li, D. Agronomic trait variations and ploidy differentiation of kiwiberries in Northwest China: Implication for breeding. Front. Plant Sci. 2017, 8, 711. [Google Scholar] [CrossRef] [PubMed]
- Evans, K.M.; Fernández-Fernández, F.; Govan, C. Harmonising fingerprinting protocols to allow comparisons between germplasm collections–Pyrus. In Acta Horticulturae, Proceedings of the XIIthEucarpia Symposium on Fruit Breeding and Genetics, Zaragoza, Spain, 16–20 September 2007; International Society for Horticultural Science: Leuven, Belgium, 2009; Volume 814, pp. 103–106. [Google Scholar] [CrossRef]
- Liebhard, R.; Gianfranceschi, L.; Koller, B.; Ryder, C.D.; Tarchini, R.; van de Weg, E.; Gessler, C. Development and characterization of 140 microsatellites in apple (Malus×domestica Borkh.). Mol. Breed. 2002, 10, 217–241. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Nei, M.; Li, W.H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef] [PubMed]
- Perrier, X.; Jacquemoud-Collet, J.P.D. DARwin Software. 2006. Available online: http://darwin.cirad.fr/ (accessed on 20 June 2016).
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Sehic, J.; Garkava-Gustavsson, L.; Fernández-Fernández, F.; Nyboma, H. Genetic diversity in a collection of European pear (Pyrus communis) cultivars determined with SSR markers chosen by ECPGR. Sci. Hortic. 2012, 145, 39–45. [Google Scholar] [CrossRef]
- Bassil, N.; Hummer, K.E.; Postman, J.D.; Fazio, G.; Baldo, A.; Armas, I.; Williams, R. Nomenclature and genetic relationships of apples and pears from Terceira Island. Genet. Resour. Crop Evol. 2009, 56, 339–352. [Google Scholar] [CrossRef]
- Miranda, C.; Urrestarazu, J.; Santesteban, L.G.; Royo, J.B.; Urbina, V. Genetic diversity and structure in a collection of ancient Spanish pear cultivars assessed by microsatellite markers. J. Am. Soc. Hortic. Sci. 2010, 135, 428–437. [Google Scholar] [CrossRef]
- Bennici, S.; Las Casas, G.; Distefano, G.; Di Guardo, M.; Continella, A.; Ferlito, F.; Gentile, A.; La Malfa, S. Elucidating the contribution of wild related species on autochthonous pear germplasm: A case study from Mount Etna. PLoS ONE 2018, 13, e0198512. [Google Scholar] [CrossRef] [PubMed]
- Claessen, H.; Keulemans, W.; Poel, B.V.; Storme, M. Finding a compatible partner: Self-incompatibility in European pear (Pyrus communis); molecular control, genetic determination, and impact on fertilization and fruit set. Front. Plant Sci. 2019, 10, 407. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.K.; Hunter, D.M.; Lukens, L.; Strommer, J.N. European and Asian Pears: Simple Sequence Repeat-Polyacrylamide Gel Electrophoresis-base analysis of commercially important North-American cultivars. Hortscience 2006, 41, 304–309. [Google Scholar] [CrossRef]
- Hokanson, S.C.; Lamboy, W.F.; Szewc-McFadden, A.K.; McFerson, J.R. Microsatellite (SSR) variation in a collection of Malus (apple) species and hybrids. Euphytica 2001, 118, 281–294. [Google Scholar] [CrossRef]
- Godinho, C.; Lampreia, F. Caracterização de Variedades Regionais Portuguesas, 1st ed.; Direcção-Geral de Protecção das Culturas: Lisboa, Portugal, 2006; pp. IV1–IV89.
- Pelsy, F. Molecular and cellular mechanisms of diversity within grapevine varieties. Heredity 2010, 104, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Meneghetti, S.; Bavaresco, L.; Calò, A.; Costacurta, A. Inter- and intra-varietal genetic variability in Vitis vinifera L. In The Mediterranean Genetic Code–Grapevine and Olive; IntechOpen Publishers: Rijeka, Croatia, 2013; pp. 73–95. [Google Scholar] [CrossRef]
- Meneghetti, S.; Poljuha, D.; Frare, E.; Costacurta, A.; Morreale, G.; Bavaresco, L.; Calò, A. Inter- and Intra-varietal genetic variability in Malvasia cultivars. Mol. Biotechnol. 2012, 50, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, E.; Carrasquinho, I.; St. Aubyn, A.; Martins, A. Broad-sense heritability in mixed models for grapevine initial selection trials. Euphytica 2013, 189, 379–391. [Google Scholar] [CrossRef]
- Breton, C.; Pinatel, C.; Médail, F.; Bonhomme, F.; Bervillé, A. Comparison between classical and Bayesian methods to investigate the history of olive cultivars using SSR-polymorphisms. Plant Sci. 2008, 175, 524–532. [Google Scholar] [CrossRef]
- Belaj, A.; Rallo, L.; Trujillo, I.; Baldoni, L. Using RAPD and AFLP markers to distinguish individuals obtained by clonal selection of “Arbequina” and “Manzanilla de Sevilla” olive. Hortscience 2004, 39, 1566–1570. [Google Scholar] [CrossRef]
- Sarri, V.; Baldoni, L.; Porceddu, A.; Cultrera, N.G.M.; Contento, A.; Frediani, M.; Belaj, A.; Trujillo, I.; Cionini, P.G. Microsatellite markers are powerful tools for discriminating among olive cultivars and assigning them to geographically defined populations. Genome 2006, 49, 1606–1615. [Google Scholar] [CrossRef] [PubMed]
- Ferradini, N.; Lancioni, H.; Torricelli, R.; Russi, L.; Dalla Ragione, I.; Cardinali, I.; Marconi, G.; Gramaccia, M.; Concezzi, L.; Achill, I.A.; et al. Characterization and phylogenetic analysis of ancient Italian landraces of pear. Front. Plant Sci. 2017, 8, 751. [Google Scholar] [CrossRef] [PubMed]
- Dickson, E.E.; Arumuganathan, K.; Krosovich, S.; Doyle, J.J. Nuclear DNA content variation within the Rosaceae. Am. J. Bot. 1992, 79, 1081–1086. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Reference | Local Designation | Field Collection |
|---|---|---|
| A1 | ÁguaouAguinha | INIAV-Alcobaça |
| A2 | Amendoa | INIAV-Alcobaça |
| V21 | Amendoa | Quinta da Sobreira, Vidago |
| V04 | Amendoa II | Quinta da Sobreira, Vidago |
| V12 | Amendoa I | Quinta da Sobreira, Vidago |
| S15 | Amorim | Quinta de Sergude, Felgueiras |
| V48 | Amorim Branco | Quinta da Sobreira, Vidago |
| V39 | Baguim | Quinta da Sobreira, Vidago |
| S05 | Bela de Junho | Quinta de Sergude, Felgueiras |
| S11 | Bela Feia 1 | Quinta de Sergude, Felgueiras |
| V57 | Bela Feia 2 | Quinta da Sobreira, Vidago |
| S07 | Bojarda | Quinta de Sergude, Felgueiras |
| V79 | CabacinhaPrecoce | Quinta da Sobreira, Vidago |
| A3 | CabeçaPequena | INIAV-Alcobaça |
| V115 | Carapinheira | Quinta da Sobreira, Vidago |
| S12 | CarapinheiraBranca | Quinta de Sergude, Felgueiras |
| A4 | Carvalhal | INIAV-Alcobaça |
| V07 | CoradaParda | Quinta da Sobreira, Vidago |
| V41 | Coradinha | Quinta da Sobreira, Vidago |
| S14 | Coxa de Freira | Quinta de Sergude, Felgueiras |
| A7 | Cristo | INIAV-Alcobaça |
| V19 | D. Joaquina | Quinta da Sobreira, Vidago |
| V80 | Fim de Século | Quinta da Sobreira, Vidago |
| V28 | Formiga | Quinta da Sobreira, Vidago |
| V97 | LambeosDedos | Quinta da Sobreira, Vidago |
| A8 | Marcelina | INIAV-Alcobaça |
| A9 | Marmela | INIAV-Alcobaça |
| V13 | Marmela 1 | Quinta da Sobreira, Vidago |
| V89 | Marmela 2 | Quinta da Sobreira, Vidago |
| S18 | Marquesinha | Quinta de Sergude, Felgueiras |
| V14 | Nacional | Quinta da Sobreira, Vidago |
| A10 | Pera Bonita | INIAV-Alcobaça |
| V26 | PeraCabaça | Quinta da Sobreira, Vidago |
| V06 | Pera de Inverno | Quinta da Sobreira, Vidago |
| A11 | PeraMelão | INIAV-Alcobaça |
| A12 | Perola | INIAV-Alcobaça |
| V16 | PerolaAmarela | Quinta da Sobreira, Vidago |
| V66 | PerolaAmarela 1 | Quinta da Sobreira, Vidago |
| V75 | PerolaAmarela 2 | Quinta da Sobreira, Vidago |
| V63 | PerolaBranca | Quinta da Sobreira, Vidago |
| V10 | Pigarça | Quinta da Sobreira, Vidago |
| V95 | Rabiça | Quinta da Sobreira, Vidago |
| V22 | RaboTorto | Quinta da Sobreira, Vidago |
| V102 | Rosa de Soure | Quinta da Sobreira, Vidago |
| V72 | Rugosa | Quinta da Sobreira, Vidago |
| A14 | S. Bartolomeu | INIAV-Alcobaça |
| V43 | S. Bento de Chaves | Quinta da Sobreira, Vidago |
| A15 | S. Crispim | INIAV-Alcobaça |
| V82 | S. João de Silgueiros | Quinta da Sobreira, Vidago |
| V11 | Tipo Cabaça | Quinta da Sobreira, Vidago |
| AF | Abbé Fétel | Quinta da Sobreira, Vidago |
| S19 | Conference | Quinta de Sergude, Felgueiras |
| Rocha1—Rocha8 | Pera Rocha | INIAV-Alcobaça |
| Rocha9—Rocha79 | Pera Rocha | Quinta de S. João, Caldas da Rainha |
| Locus | Na | Ne | Ho | He | PIC |
|---|---|---|---|---|---|
| CH01d08 | 8 | 4.114 | 0.766 | 0.757 | 0.657 |
| CH01f07a | 13 | 6.786 | 0.894 | 0.853 | 0.654 |
| CH05c06 | 11 | 3.034 | 0.787 | 0.670 | 0.301 |
| CH03g07 | 13 | 8.801 | 0.894 | 0.886 | 0.714 |
| EMPc11 | 11 | 7.425 | 0.872 | 0.865 | 0.553 |
| EMPc117 | 14 | 4.436 | 0.702 | 0.775 | 0.582 |
| CH01d09 | 16 | 8.166 | 0.979 | 0.878 | 0.728 |
| CH02b10 | 11 | 7.058 | 0.915 | 0.858 | 0.744 |
| GD96 | 8 | 4.566 | 1.000 | 0.781 | 0.642 |
| CH03d12 | 13 | 7.477 | 0.848 | 0.866 | 0.537 |
| CH04e03 | 11 | 1.846 | 0.457 | 0.458 | 0.337 |
| Average | 11.72 | 5.79 | 0.83 | 0.79 | 0.59 |
| Synonymies (Name and Ref.) |
|---|
| “Amendoa I” (V12) = “RaboTorto” (V22) |
| “Amorim” (S15) = “Amorim Branco” (V48) |
| “Carapinheira” (V115) = “CarapinheiraBranca” (S12) |
| Homonymies (Name and Ref.) |
|---|
| “Amendoa” (A2); “Amendoa” (V21); “Amendoa II” (V04) |
| “Bela Feia 1” (S11); “Bela Feia 2” (V57) |
| “Marmela” (A9); “Marmela 1” (V13); “Marmela 2” (V89) |
| “PerolaAmarela” (V16); “PerolaAmarela1”(V66); “PerolaAmarela 2” (V75) |
| Axes | 1 | 2 | 3 |
|---|---|---|---|
| % | 33.51 | 7.34 | 5.09 |
| Cum % | 33.51 | 40.85 | 45.94 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Queiroz, Á.; Bagoin Guimarães, J.; Sánchez, C.; Simões, F.; Maia de Sousa, R.; Viegas, W.; Veloso, M.M. Genetic Diversity and Structure of the Portuguese Pear (Pyrus communis L.) Germplasm. Sustainability 2019, 11, 5340. https://doi.org/10.3390/su11195340
Queiroz Á, Bagoin Guimarães J, Sánchez C, Simões F, Maia de Sousa R, Viegas W, Veloso MM. Genetic Diversity and Structure of the Portuguese Pear (Pyrus communis L.) Germplasm. Sustainability. 2019; 11(19):5340. https://doi.org/10.3390/su11195340
Chicago/Turabian StyleQueiroz, Álvaro, Joana Bagoin Guimarães, Claudia Sánchez, Fernanda Simões, Rui Maia de Sousa, Wanda Viegas, and Maria Manuela Veloso. 2019. "Genetic Diversity and Structure of the Portuguese Pear (Pyrus communis L.) Germplasm" Sustainability 11, no. 19: 5340. https://doi.org/10.3390/su11195340
APA StyleQueiroz, Á., Bagoin Guimarães, J., Sánchez, C., Simões, F., Maia de Sousa, R., Viegas, W., & Veloso, M. M. (2019). Genetic Diversity and Structure of the Portuguese Pear (Pyrus communis L.) Germplasm. Sustainability, 11(19), 5340. https://doi.org/10.3390/su11195340