Effects of High Impact Grazing on Species Diversity and Plant Functional Groups in Grasslands of Northern Argentina

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Layout
2.3. Grassland Species Composition, Diversity and Plant Functional Groups
2.4. Statistical Analysis
3. Results
3.1. Plant Species Identified on the Plots
3.2. Standing Dead and Green Biomass Ground Cover
3.3. Green Biomass Ground Cover of Plant Functional Groups
3.4. Diversity
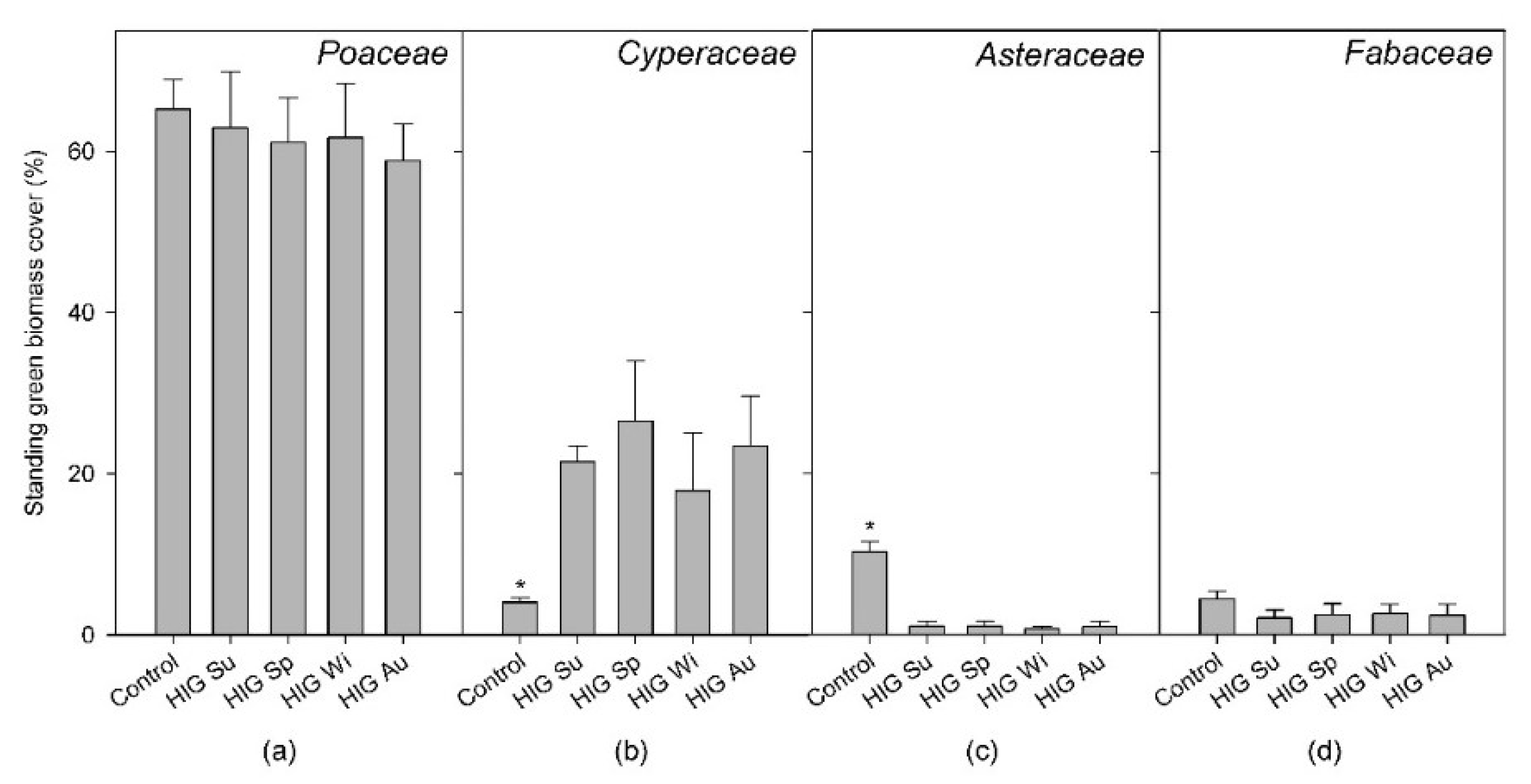
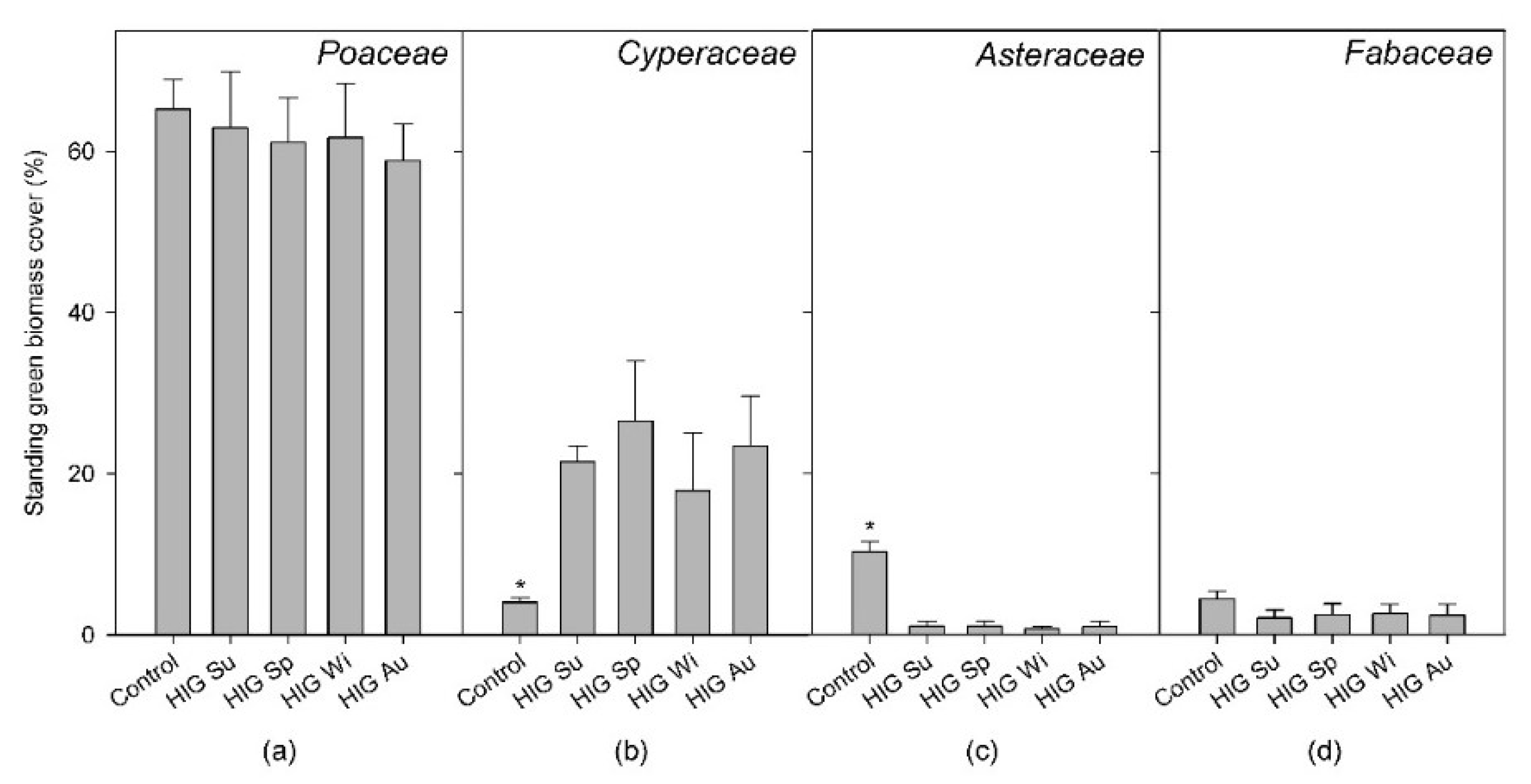
3.5. Relative Green Biomass Share of Selected Plant Families and Species
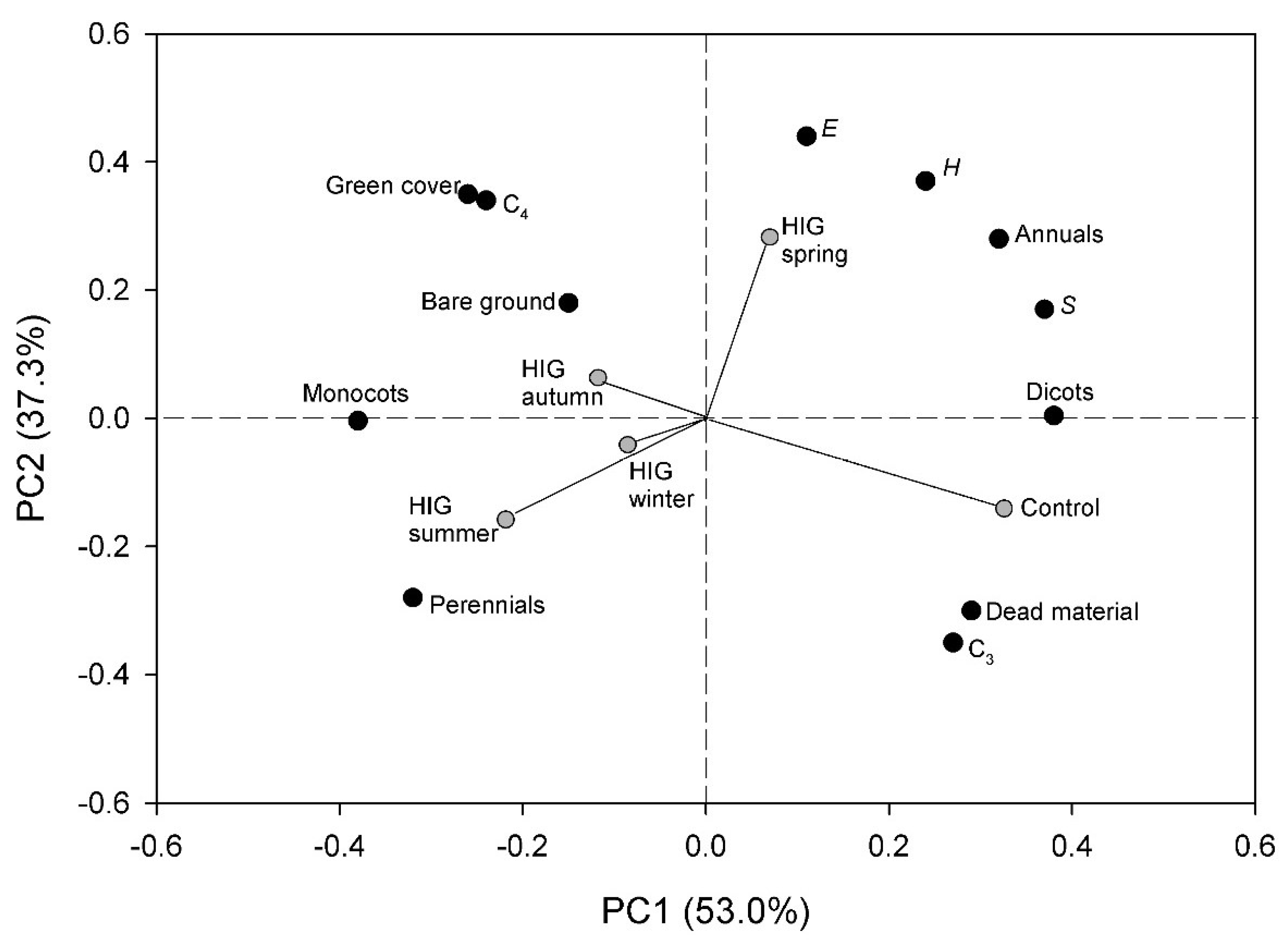
3.6. Principal Component Analysis (PCA)
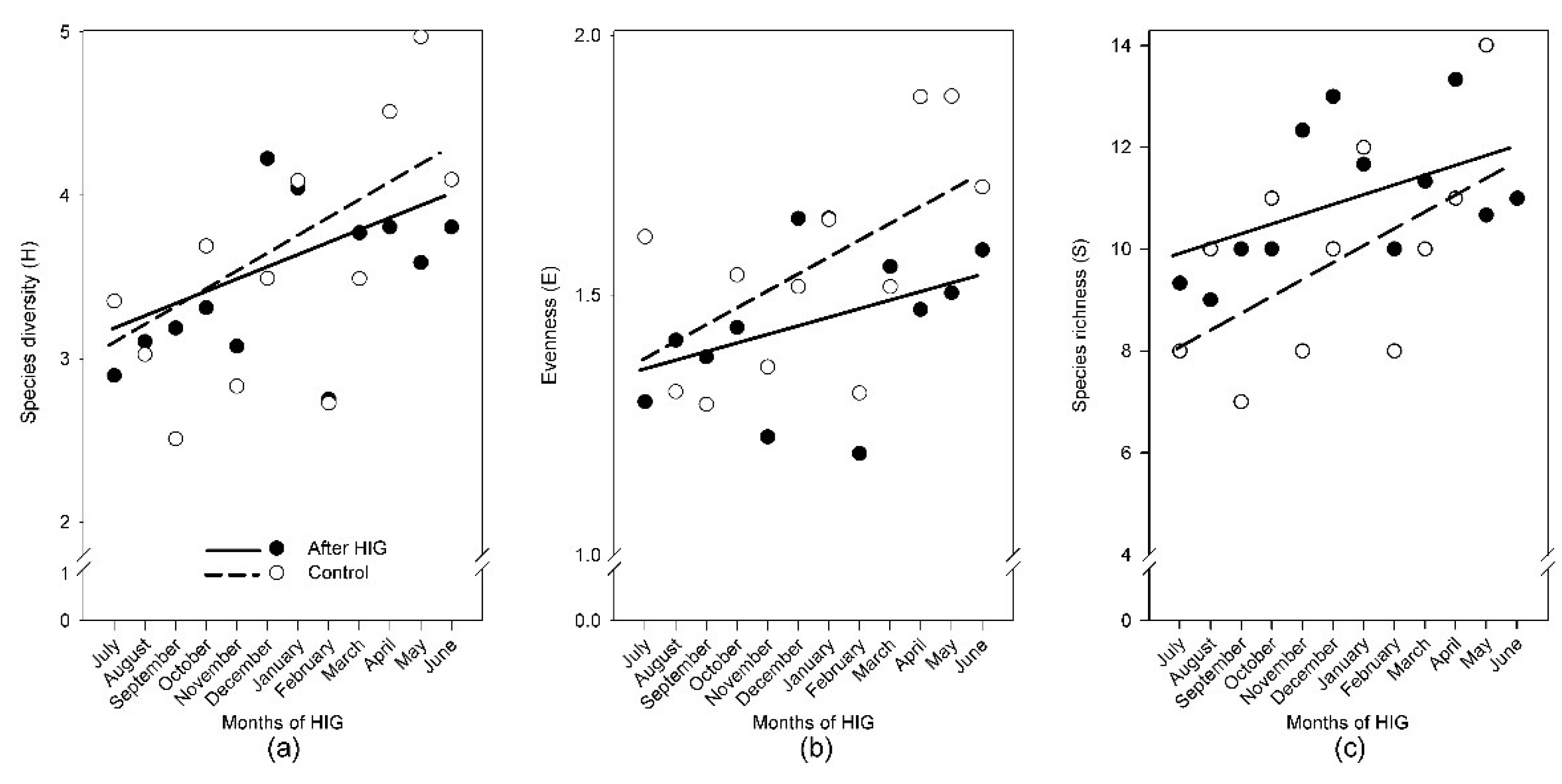
3.7. Monthly Monitoring of Diversity One Year after HIG
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Species | Familiy | 1 Number of Cotyledons | 2 Life Cycle | 3 Photo-Synthesis Pathway | 4 Response to HIG |
|---|---|---|---|---|---|
| Justicia laevilinguis (Nees) Lindau | Acanthaceae | 2 | Pe | C4 | i |
| Ruellia sp. L. | Acanthaceae | 2 | An | C4 | i |
| Alternanthera philoxeroides (Mart.) Griseb. | Amaranthaceae | 2 | Pe | C4 | in |
| Amaranthus sp. L. | Amaranthaceae | 2 | An | C4 | i |
| Gomphrena celocioides Mart. | Amaranthaceae | 2 | An | C4 | d |
| Zepyranthes sp. Herb. | Amaryllidaceae | 1 | Pe | C3 | i |
| Eryngium coronatum Hook. & Arn. | Apiaceae | 2 | Pe | C3 | i |
| Eryngium ebracteatum Lam. | Apiaceae | 2 | Pe | C3 | i |
| Eryngium horridum Urb. ex H. Wolff, Malme | Apiaceae | 2 | Pe | C3 | i |
| Ammis majus L. | Apiaceae | 2 | An | CAM | i |
| Apium leptophyllum (Pers) F. Muell. Ex Benth | Apiaceae | 2 | Pe | CAM | i |
| Aristolochia fimbriata Cham. | Aristolochiaceae | 2 | Pe | C3 | i |
| Asceplias curassavica L. | Asclepiadaceae | 2 | Pe | C3 | i |
| Acmella sp. R.K. Jansen | Asteraceae | 2 | An | C4 | i |
| Baccharis coridifolia DC. | Asteraceae | 2 | Pe | C4 | i |
| Baccharis notorsegila Griseb. | Asteraceae | 2 | Pe | C4 | d |
| Baccharis punctulata DC. | Asteraceae | 2 | Pe | C4 | in |
| Baccharis sp. DC. | Asteraceae | 2 | Pe | C4 | i |
| Bidens pilosa L. | Asteraceae | 2 | An | C4 | in |
| Chaptalia nutans (L.) Polák | Asteraceae | 2 | Pe | C4 | i |
| Conyza bonariensis (L.) Cronquist | Asteraceae | 2 | An | C4 | i |
| Eupatorium candolleanum Hook. & Arn. | Asteraceae | 2 | Pe | C4 | i |
| Eupatorium macrocephalum Less. | Asteraceae | 2 | Pe | C4 | d |
| Eupatorium clematideum Griseb. | Asteraceae | 2 | Pe | C4 | in |
| Eupatorium subhastatum Hock. & Arn. | Asteraceae | 2 | Pe | C4 | i |
| Eupatorium sp. Hock. & Arn. | Asteraceae | 2 | Pe | C4 | in |
| Mikania coridifolia (L.f.) Willd. | Asteraceae | 2 | Pe | C4 | in |
| Orthoppapus angustifolius (Sw.) | Asteraceae | 2 | Pe | C4 | d |
| Pterocaulon lorentzii Malme | Asteraceae | 2 | An | C4 | in |
| Pterocaulon pycnostachyum (Michx.) Elliott | Asteraceae | 2 | Pe | C4 | in |
| Pterocaulon sp. DC. | Asteraceae | 2 | Pe | C4 | d |
| Senecio grisebachii Baker | Asteraceae | 2 | Pe | C4 | i |
| Solidago chilensis Meyen | Asteraceae | 2 | Pe | C4 | i |
| Vernonia incana (Less.) DC. | Asteraceae | 2 | Pe | C4 | i |
| Vernonia chamaedrys Lees. | Asteraceae | 2 | Pe | C4 | i |
| Vernonia rubricaulis Hum. & Bonpl. | Asteraceae | 2 | Pe | C4 | d |
| Chenopodium nigra L. | Chenopodiaceae | 2 | An | C4 | i |
| Commelina erecta L. | Commelinaceae | 1 | Pe | CAM | i |
| Tripogandra radiata (C.B. Clarke) Bacigalupo | Commelinaceae | 1 | Pe | CAM | i |
| Dichondra repens J.R. Forst. & G. Forst. | Convolvulaceae | 2 | Pe | C4 | i |
| Evolvulus sericeus Sw. | Convolvulaceae | 2 | Pe | C4 | d |
| Ascolepsis brasiliensis (Kunth) Benth. Ex Clarke. | Cyperaceae | 1 | Pe | C4 | i |
| Carex sororia Kunth | Cyperaceae | 1 | Pe | C4 | i |
| Cyperus aggregatus (Willd.) Endl. | Cyperaceae | 1 | Pe | C3 | in |
| Cyperus entrerrianus Boeckeler | Cyperaceae | 1 | Pe | C4 | i |
| Cyperus haspan ssp. Juncoides | Cyperaceae | 1 | Pe | C3 | i |
| Cyperus iria L. | Cyperaceae | 1 | Pe | C4 | in |
| Cyperus obtusatus (J. Presl & C. Presl) Mattf. & Kük. | Cyperaceae | 1 | Pe | C4 | i |
| Cyperus rigens C. Presl | Cyperaceae | 1 | Pe | C4 | i |
| Cyperus virens Michx. | Cyperaceae | 1 | Pe | C4 | i |
| Eleocharis nodulosa (Roth) Schult. | Cyperaceae | 1 | Pe | C4 | i |
| Eleocharis viridans Kük ex. Osten | Cyperaceae | 1 | Pe | C4 | i |
| Fimbristylis dichotoma (L.) Vahl | Cyperaceae | 1 | Pe | C4 | i |
| Rhynchospora corymbosa (L.) Britton | Cyperaceae | 1 | Pe | C4 | in |
| Rhynchospora scutellata Griseb. | Cyperaceae | 1 | Pe | C4 | i |
| Rhynchospora tenuis Link | Cyperaceae | 1 | Pe | C4 | i |
| Scleria sellowiana Kunth | Cyperaceae | 1 | Pe | C4 | d |
| Pteridium aquilinum (L.) Kuhn | Dennstaedtiaceae | - | Pe | C3 | d |
| Scoparia dulcis L. | Escrofulariaceae | 2 | An | C3 | i |
| Scoparia muricata L. | Escrofulariaceae | 2 | An | C3 | in |
| Phyllanthus stipulatus (Raf.) G.L. Webster | Euphorbiaceae | 2 | An | C4 | i |
| Euphorbia prostrata Aiton | Euphorbiaceae | 2 | An | C4 | d |
| Tragia geraniifolia Klotzsch ex Müll.Arg. | Euphorbiaceae | 2 | Pe | C4 | i |
| Aeschynomene americana L. | Fabaceae | 2 | Pe | C3 | i |
| Chamaecrista rotundifolia (Pers.) Greene | Fabaceae | 2 | Pe | C3 | d |
| Desmanthus virgatus (L.) Wild. | Fabaceae | 2 | Pe | C3 | i |
| Desmodium pachyrizum Vogel | Fabaceae | 2 | Pe | C3 | i |
| Rhynchosia laterita Burkart | Fabaceae | 2 | Pe | C3 | in |
| Desmanthus depressus Willd. | Fabaceae | 2 | Pe | C3 | in |
| Desmodium incanum DC. | Fabaceae | 2 | Pe | C3 | i |
| Discolobium sp. Benth. | Fabaceae | 2 | Pe | C3 | in |
| Galactia marginalis Benth. | Fabaceae | 2 | An | C3 | i |
| Indigofera asperifolia Benth. | Fabaceae | 2 | Pe | C3 | i |
| Leucaena leucocephala (Lam.) de Wit | Fabaceae | 2 | Pe | C3 | i |
| Macroptilium lathyroides (L.) Urb. | Fabaceae | 2 | Pe | C3 | d |
| Macroptilium postratum Benth. (Urb.) | Fabaceae | 2 | An | C3 | d |
| Phaseolus sp. Benth. | Fabaceae | 2 | Pe | C3 | in |
| Rhynchosia edulis Griseb. | Fabaceae | 2 | Pe | C3 | i |
| Rhynchosia sp. Lour. | Fabaceae | 2 | Pe | C3 | in |
| Stylosanthes hippocampoides Mohlenbr. | Fabaceae | 2 | Pe | C3 | i |
| Stylosanthes montevidensis Vogel | Fabaceae | 2 | Pe | C3 | in |
| Hydrolea spinosa L. | Hydrophyllaceae | 2 | An | C3 | i |
| Sisyrinchium sp. Baker | Iridaceae | 1 | Pe | C3 | d |
| Juncus microcephalus Kunth | Juncaceae | 1 | Pe | C3 | i |
| Hyptis lappacea Benth. | Labiadae | 2 | An | C3 | i |
| Nothoscordum inodorum (Aiton) G. Nicholson | Liliaceae | 1 | Pe | C3 | in |
| Selaginella sp. Spring | Lycopdiopsida | - | Pe | C3 | i |
| Cuphea carthagenensis (Jacq.) J. F. Macbr. | Lythraceae | 2 | Pe | C3 | i |
| Cuphea lysimachioides Cham. & Schltdl. | Lythraceae | 2 | Pe | C3 | i |
| Cuphea sp. Koehne | Lythraceae | 2 | Pe | C3 | i |
| Heymia salicifolia (Kunth) Link & Otto | Lythraceae | 2 | Pe | C3 | i |
| Krapovickasia sp. Fryxell | Malvaceae | 2 | An | C3 | i |
| Malvastrum coromandelianum (L.) Garcke | Malvaceae | 2 | An | C3 | i |
| Melochia hernannioides A. St. Hil. | Malvaceae | 2 | Pe | C3 | i |
| Sida rhombifolia L. | Malvaceae | 2 | Pe | C3 | i |
| Sida tuberculata R.E.Fr. | Malvaceae | 2 | Pe | C3 | i |
| Marsilea consinea Mirb. | Marsiliaceae | - | Pe | C3 | i |
| Cissampelos sp. Kunth | Menispermaceae | 2 | Pe | C3 | i |
| Ludwigia major (Micheli) Ramamoorthy | Onagraceae | 2 | Pe | C3 | i |
| Orchidia sp. Juss. | Orchidaceae | 1 | Pe | CAM | in |
| Oxalis sp. L. | Oxalidaceae | 2 | Pe | CAM | i |
| Passiflora coerulea L. | Passifloraceae | 2 | Pe | CAM | i |
| Plantago officinalis Crantz | Plantaginaceae | 2 | Pe | CAM | i |
| Andropogon lateralis Ness | Poaceae | 1 | Pe | C4 | i |
| Axonopus affinis Chase | Poaceae | 1 | Pe | C4 | in |
| Axonopus compressus (Sw.) P. Beauv. | Poaceae | 1 | Pe | C4 | i |
| Axonopus fissifolius (Raddi) Kuhlm. | Poaceae | 1 | Pe | C4 | i |
| Bothriochloa laguroides DC. | Poaceae | 1 | Pe | C4 | i |
| Bothriochloa saccharoides Sw. | Poaceae | 1 | Pe | C4 | i |
| Briza uniolae (Nees) Steud. | Poaceae | 1 | An | C4 | i |
| Chloris distichophylla Lag. | Poaceae | 1 | Pe | C4 | i |
| Cynodon dactylon (L.) Pers. | Poaceae | 1 | Pe | C4 | i |
| Digitaria insularis (L.) Mez ex Ekman | Poaceae | 1 | Pe | C4 | in |
| Digitaria phaeotrix (Trin.) Parodi | Poaceae | 1 | Pe | C4 | i |
| Digitaria sp. Haller | Poaceae | 1 | An | C4 | i |
| Eleusine indica (L.) Gaertn. | Poaceae | 1 | Pe | C4 | i |
| Eleusine tristachya Lam | Poaceae | 1 | Pe | C4 | i |
| Elyonurus muticus (Spreng.) Kuntze | Poaceae | 1 | An | C4 | i |
| Eragrostis airoides Nees | Poaceae | 1 | Pe | C4 | i |
| Eragrostis bahiensis Roem. & Schult. | Poaceae | 1 | Pe | C4 | i |
| Hemarthria altissima (Poir) Stapf & C.E. Hubb. | Poaceae | 1 | Pe | C4 | i |
| Leersia hexandra Sw. | Poaceae | 1 | Pe | C3 | i |
| Panicum miliaceum L. | Poaceae | 1 | Pe | C4 | i |
| Panicum milioides Ness. Ex Trin. | Poaceae | 1 | Pe | C4 | i |
| Panicum prionitis Nees | Poaceae | 1 | Pe | C4 | - |
| Panicum sp. L. | Poaceae | 1 | Pe | C4 | in |
| Paspalum acuminatum Raddi | Poaceae | 1 | Pe | C4 | i |
| Paspalum almun Chase | Poaceae | 1 | Pe | C4 | i |
| Paspalum notatum Flügé | Poaceae | 1 | Pe | C4 | i |
| Paspalum plicatulum Michx. | Poaceae | 1 | Pe | C4 | i |
| Paspalum simplex Morong | Poaceae | 1 | Pe | C4 | i |
| Paspalum urvillei Steud. | Poaceae | 1 | Pe | C4 | i |
| Phalaris sp. Jansen & Wacht. | Poaceae | 1 | An | C4 | i |
| Piptochaetium montevidense (Spreng.) Parodi | Poaceae | 1 | Pe | C4 | in |
| Rotboellia selloana L.F. | Poaceae | 1 | Pe | C4 | i |
| Schizachyrium microstachyum (Desv.) Roseng. | Poaceae | 1 | Pe | C4 | i |
| Setaria geniculata P.Beauv. | Poaceae | 1 | Pe | C4 | i |
| Sorghastrum pellitum (Hack.) Parodi | Poaceae | 1 | Pe | C4 | in |
| Sorghastrum setosum (Griseb.) Hitchc. | Poaceae | 1 | Pe | C4 | i |
| Sporobolus indicus (L.) R. Br. | Poaceae | 1 | Pe | C4 | i |
| Sporobolus monandrus Roseng., B.R. Arrill. & Izag. | Poaceae | 1 | Pe | C4 | in |
| Sporobolus poiretii (Roem. & Schult.) Hitchs. | Poaceae | 1 | Pe | C4 | i |
| Sporobolus sp. R. Br. | Poaceae | 1 | Pe | C4 | i |
| Steinchisma hians (Elliott) Nash | Poaceae | 1 | Pe | C4 | i |
| Steinchisma laxa Sw. | Poaceae | 1 | Pe | C4 | i |
| Tridens brasiliensis (Steud.) Parodi | Poaceae | 1 | Pe | C3 | i |
| Polygala molluginifolia A. St. Hil. | Polygalaceae | 2 | An | C3 | i |
| Polygala obovata A. St. Hil. & Moq. | Polygalaceae | 2 | An | C3 | i |
| Polygala sp. DC. | Polygalaceae | 2 | An | C3 | i |
| Muehlenbeckia sagittifolia (Ortega) Meisn. | Polygonaceae | 2 | Pe | C4 | i |
| Poligonum punctatum Elliot | Polygonaceae | 2 | Pe | C3 | i |
| Clematis bonariensis Juss. Ex DC. | Ranunculaceae | 2 | Pe | C3 | i |
| Relbunium richardianum (Gillies ex Hook. & Arn.) Hicken | Rubiaceae | 2 | An | C3 | d |
| Rubiacea Juss. | Rubiaceae | 2 | An | C3 | d |
| Bouchetia anomale (Miers) Britton & Ruby | Solanaceae | 2 | Pe | C3 | i |
| Petunia sp. Juss. | Solanaceae | 2 | An | C3 | i |
| Phisalis viscosa L. | Solanaceae | 2 | Pe | C3 | i |
| Solanum granulosum-leprosum Dunal | Solanaceae | 2 | Pe | C3 | in |
| Turnera sidoides DC. | Turneraceae | 2 | An | C3 | d |
| Lantana sp. Moldenke | Verbenaceae | 2 | Pe | C3 | d |
| Lippia sp. Moldenke | Verbenaceae | 2 | Pe | C3 | d |
| Glandularia peruviana (L.) Small | Verbenaceae | 2 | Pe | C3 | i |
| Glandularia rigida Sprengel | Verbenaceae | 2 | Pe | C3 | i |
| Verbena litoralis Kunth | Verbenaceae | 2 | Pe | C3 | i |
| Verbena rigida Spreng. | Verbenaceae | 2 | Pe | C3 | i |
References
- Di Gregorio, A.; Jansen, L.J.M. Land Cover Classification System. Classification Concepts and User Manual; FAO: Rome, Italy, 2005; p. 193. [Google Scholar]
- Dixon, A.P.; Faber-Langendoen, D.; Josse, C.; Morrison, J.; Loucks, C.J. Distribution Mapping of World Grassland Types. J. Biogeogr. 2014, 41, 2003–2019. [Google Scholar] [CrossRef]
- Lund, H.G. Accounting for the world’s rangelands. Rangelands 2007, 29, 3–10. [Google Scholar] [CrossRef]
- Frame, J. Improved Grassland Management, new ed.; Crowood: Ramsbury, UK, 2011; p. 352. [Google Scholar]
- Grau, H.R.; Torres, R.; Gasparri, N.I.; Blendinger, P.G.; Marinaro, S.; Macchi, L. Natural Grasslands in The Chaco. A Neglected Ecosystem Under Threat by Agriculture Expansion and Forest-Oriented Conservation Policies. J. Arid. Environ. 2015, 123, 40–46. [Google Scholar] [CrossRef]
- Carnevali, R. Fitogeografía de la Provincia de Corrientes; NTA—Gobierno de la provincia de Corrientes: Corrientes, Argentina, 1994; p. 324. [Google Scholar]
- Rosengurtt, B. Tablas de Comportamiento de las Especies de Plantas de Campos Naturales del Uruguay. Montevideo; Universidad de la República, Facultad de Agronomía, Dirección General de Extensión Universitaria: Montevideo, Uruguay, 1979; p. 15. [Google Scholar]
- Cingolani, A.M.; Vaieretti, M.V.; Giorgis, M.A.; Poca, M.; Tecco, P.A.; Gurvich, D.E. Can Livestock Grazing Maintain Landscape Diversity and Stability in An Ecosystem That Evolved with Wild Herbivores? Perspect. Plant Ecol. Evol. Syst. 2014, 16, 143–153. [Google Scholar] [CrossRef]
- Hendricks, H.H.; Bond, W.J.; Midgley, J.J.; Novellie, P.A. Plant Species Richness and Composition Along Livestock Grazing Intensity Gradients in a Namaqualand (South Africa) Protected Area. Plant Ecol. 2005, 176, 19–33. [Google Scholar] [CrossRef]
- Milchunas, D.G.; Lauenroth, W.K. Quantitative Effects of Grazing on Vegetation and Soils over a Global Range of Environments. Ecol. Monogr. 1993, 63, 327–366. [Google Scholar] [CrossRef]
- Rockström, J.; Steffen, W.; Noone, K.; Persson, A.; Chapin, F.S.; Lambin, E.F.; Lenton, T.M.; Scheffer, M.; Folke, C.; Schellnhuber, H.J.; et al. A Safe Operating Space for Humanity. Nature 2009, 461, 472–475. [Google Scholar] [CrossRef] [PubMed]
- West, N.E. Biodiversity of Rangelands. J. Range Manag. 1993, 46, 2–13. [Google Scholar] [CrossRef]
- Alkemade, R.; Reid, R.S.; van den Berg, M.; de Leeuw, J.; Jeuken, M. Assessing the Impacts of Livestock Production on Biodiversity in Rangeland Ecosystems. Proc. Natl. Acad. Sci. USA 2013, 110, 20900–20905. [Google Scholar] [CrossRef] [PubMed]
- Cingolani, A.M.; Noy-Meir, I.; Díaz, S. Grazing Effects or Rangeland Diversity: A Synthesis of Contemporary Models. Ecol. Appl. 2005, 15, 757–773. [Google Scholar] [CrossRef]
- Bock, C.E.; Jones, Z.F.; Bock, J.H. Relationships between Species Richness, Evenness, And Abundance in a Southwestern Savanna. Ecology 2007, 88, 1322–1327. [Google Scholar] [CrossRef] [PubMed]
- Pizzio, R.; Herrero-Jáuregui, C.; Pizzio, M.; Oesterheld, M. Impact of Stocking Rate on Species Diversity and Composition of a Subtropical Grassland in Argentina. Appl. Veg. Sci. 2016, 19, 454–461. [Google Scholar] [CrossRef]
- Watkinson, A.R.; Ormerod, S.J. Grasslands, Grazing and Biodiversity: Editors’ Introduction. J. Appl. Ecol. 2001, 38, 233–237. [Google Scholar]
- Milchunas, D.G.; Sala, O.E.; Lauenroth, W.K. A Generalized Model of The Effects of Grazing by Large Herbivores on Grassland Community Structure. Am. Nat. 1988, 132, 87–106. [Google Scholar] [CrossRef]
- Borer, E.T.; Seabloom, E.W.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Lind, E.M.; Adler, P.B.; Alberti, J.; Anderson, T.M.; Bakker, J.D.; et al. Herbivores and Nutrients Control Grassland Plant Diversity Via Light Limitation. Nature 2014, 508, 517–520. [Google Scholar] [CrossRef] [PubMed]
- Briske, D.D.; Derner, J.D.; Brown, J.R.; Fuhlendorf, S.D.; Teague, W.R.; Havstad, K.M.; Gillen, R.L.; Ash, A.J.; Willms, W.D. Rotational Grazing on Rangelands: Reconciliation of Perception and Experimental Evidence. Rangel. Ecol. Manag. 2008, 61, 3–17. [Google Scholar] [CrossRef]
- Kurtz, D.B.; Schellberg, J.; Braun, M. Ground and Satellite-Based Assessment of Rangeland Management in Sub-Tropical Argentina. Appl. Geogr. 2010, 30, 210–220. [Google Scholar] [CrossRef]
- Dowling, P.M.; Kemp, D.R.; Ball, P.D.; Langford, C.M.; Michalk, D.L.; Millar, G.D.; Simpson, P.C.; Thompson, R.P. Effect of Continuous and Time-Control Grazing on Grassland Components in South-Eastern Australia. Aust. J. Exp. Agric. 2005, 45, 369–382. [Google Scholar] [CrossRef]
- Savory, A. The Savory Grazing Method or Holistic Resource Management. Rangelands 1983, 5, 155–159. [Google Scholar]
- Mann, C.; Sherren, K. Holistic Management and Adaptive Grazing: A Trainers’ View. Sustainability 2018, 10, 1848. [Google Scholar] [CrossRef]
- Jacobo, E.J.; Rodríguez, A.M.; Bartoloni, N.; Deregibus, V.A. Rotational Grazing Effects on Rangeland Vegetation at a Farm Scale. Rangel. Ecol. Manag. 2006, 59, 249–257. [Google Scholar] [CrossRef]
- Kurtz, D.B.; Asch, F.; Giese, M.; Hüsebusch, C.; Goldfarb, M.C.; Casco, J.F. High Impact Grazing as a Management Tool to Optimize Biomass Growth in Northern Argentinean Grassland. Ecol. Indic. 2016, 63, 100–109. [Google Scholar] [CrossRef]
- Balph, D.F.; Malecheck, J.C. Cattle Trampling of Crested Wheatgrass Under Short Duration Grazing. J. Range Manag. 1985, 38, 226–227. [Google Scholar] [CrossRef]
- Moisey, D.M.; Willms, W.D.; Bork, E.W. Effect of Standing Litter on Rough Fescue Utilization by Cattle. Rangel. Ecol. Manag. 2006, 59, 197–203. [Google Scholar] [CrossRef]
- Cromsigt, J.P.G.M.; Olff, H. Dynamics of Grazing Lawn Formation: An Experimental Test of The Role of Scale-Dependent Processes. Oikos 2008, 117, 1444–1452. [Google Scholar] [CrossRef]
- Hempson, G.P.; Archibald, S.; Bond, W.J.; Ellis, R.P.; Grant, C.C.; Kruger, F.J.; Kruger, L.M.; Moxley, C.; Owen-Smith, N.; Peel, M.J.S.; et al. Ecology of Grazing Lawns in Africa. Biol. Rev. 2015, 90, 979–994. [Google Scholar] [CrossRef] [PubMed]
- McNaughton, S.J. Grazing Lawns: Animals in Herds, Plant Form, And Coevolution. Am. Nat. 1984, 124, 863–886. [Google Scholar] [CrossRef]
- Altesor, A.; Oesterheld, M.; Leoni, E.; Lezama, F.; Rodríguez, C. Effect of Grazing on Community Structure and Productivity of a Uruguayan Grassland. Plant Ecol. 2005, 179, 83–91. [Google Scholar] [CrossRef]
- Cabrera, A.L. Fitogeografía de la República Argentina. Bol. Soc. Argent. Bot. 1971, 14, 1–50. [Google Scholar]
- Escobar, E.H.; Ligier, H.D.; Melgar, R.; Matteio, H.; Vallejos, O. Mapa de Suelos de la Provincia de Corrientes 1:500.000, 1st ed.; INTA: Corrientes, Argentina, 1996; Volume 1, p. 329. [Google Scholar]
- Schinini, A.; Ciotti, E.M.; Tomei, C.E.; Castelán, M.E.; Hack, C.M. Especies Nativas de Campos Bajos Con Potencial Valor Forrajero. Agrotecnia 2004, 12, 18–22. [Google Scholar]
- Sampedro, D.; Vogel, O.; Celser, R. Suplementación de Vacunos en Pastizales Naturales; INTA: Mercedes, Argentina, 2004; pp. 1–11. [Google Scholar]
- Calvi, M. Evolución de la Ganadería Correntina; INTA: Corrientes, Argentina, 2010; pp. 1–28. [Google Scholar]
- Laurila-Pant, M.; Lehikoinen, A.; Uusitalo, L.; Venesjärvi, R. How to Value Biodiversity in Environmental Management? Ecol. Indicators 2015, 55, 1–11. [Google Scholar] [CrossRef]
- Spellerberg, I.F.; Fedor, P.J. A Tribute to Claude Shannon (1916–2001) and a Plea for More Rigorous Use of Species Richness, Species Diversity and the ‘Shannon-Wiener’ Index. Glob. Ecol. Biogeogr. 2003, 12, 177–179. [Google Scholar] [CrossRef]
- Piepho, H.P.; Büchse, A.; Richter, C. A Mixed Modeling Approach for Randomized Experiments with Repeated Measure. J. Agron. Crop Sci. 2004, 190, 230–247. [Google Scholar] [CrossRef]
- Carter, J.; Jones, A.; O’Brien, M.; Ratner, J.; Wuerthner, G. Holistic Management: Misinformation on the Science of Grazed Ecosystems. Int. J. Biodivers. 2014, 2014, 1–10. [Google Scholar] [CrossRef]
- Briske, D.D.; Bestelmeyer, B.T.; Brown, J.R.; Fuhlendorf, S.D.; Polley, H.W. The Savory Method Cannot Green Deserts or Reverse Climate Change. Rangelands 2013, 35, 72–74. [Google Scholar] [CrossRef]
- Teague, W.R.; Dowhower, S.L.; Baker, S.A.; Haile, N.; DeLaune, P.B.; Conover, D.M. Grazing Management Impacts on Vegetation, Soil Biota and Soil Chemical, Physical and Hydrological Properties in Tall Grass Prairie. Agriculture. Ecosyst. Environ. 2011, 141, 310–322. [Google Scholar] [CrossRef]
- Grime, J.P. Evidence for the Existence of Three Primary Strategies in Plants and Its Relevance to Ecological and Evolutionary Theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Fernández, J.G.; Benítez, C.A.; Pallarés, O.R. Principales Forrajeras Nativas del Medio-Este de la Provincia de Corrientes; INTA: Mercedes, Argentina, 1983; pp. 1–80. [Google Scholar]
- Caspe, S.G.; Bendersky, D.; Barbera, P. Plantas Tóxicas de la Provincia de Corrientes; INTA: Mercedes, Argentina, 2008; pp. 1–34. [Google Scholar]
- Heckathorn, S.A.; McNaughton, S.J.; Coleman, J.S. C4 Plants and Herbivory. In C4 Plant Biology; Sage, R.F., Monson, R.K., Eds.; Academic Press: San Diego, CA, USA, 1999; pp. 285–312. [Google Scholar]
- McMillan, B.R.; Pfeiffer, K.A.; Kaufman, D.W. Vegetation Responses to An Animal-Generated Disturbance (Bison Wallows) In Tallgrass Prairie. Am. Midl. Nat. 2011, 165, 60–73. [Google Scholar] [CrossRef]
- Bayer, W.; Waters-Bayer, A. Forage Husbandry, 2. publ. reprinted ed.; Macmillan: London, UK, 2013; pp. IX & 198. [Google Scholar]
- Feldman, S.R.; Bisaro, V.; Biani, N.B.; Prado, D.E. Soil Salinity Determines the Relative Abundance of C3/C4 Species in Argentinean Grasslands. Glob. Ecol. Biogeogr. 2008, 17, 708–714. [Google Scholar] [CrossRef]
- Fidelis, A.; Overbeck, G.; Pillar, V.D.; Pfadenhauer, J. Effects of Disturbance on Population Biology of The Rosette Species Eryngium Horridum Malme In Grasslands in Southern Brazil. Plant Ecol. 2008, 195, 55–67. [Google Scholar] [CrossRef]
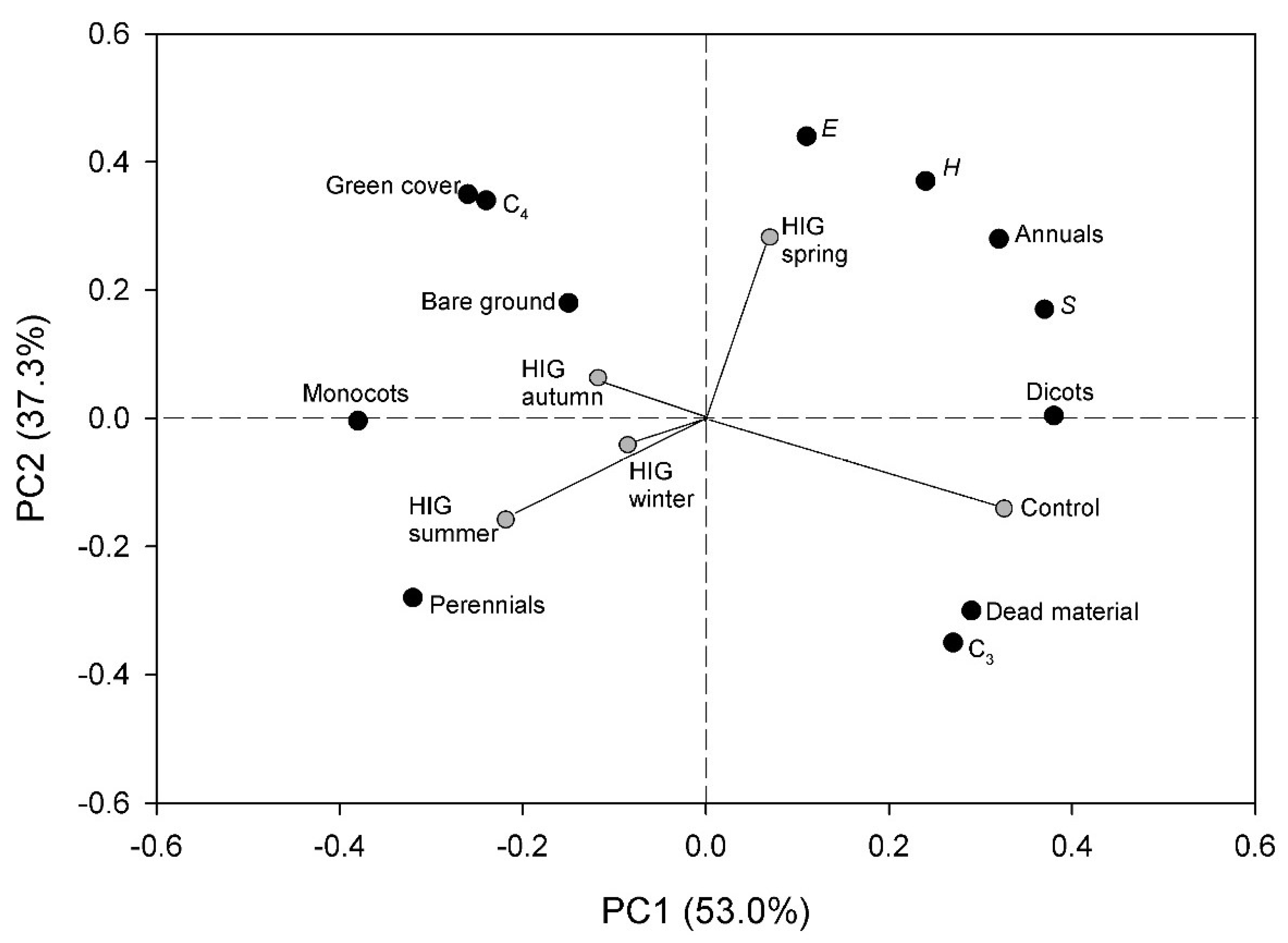
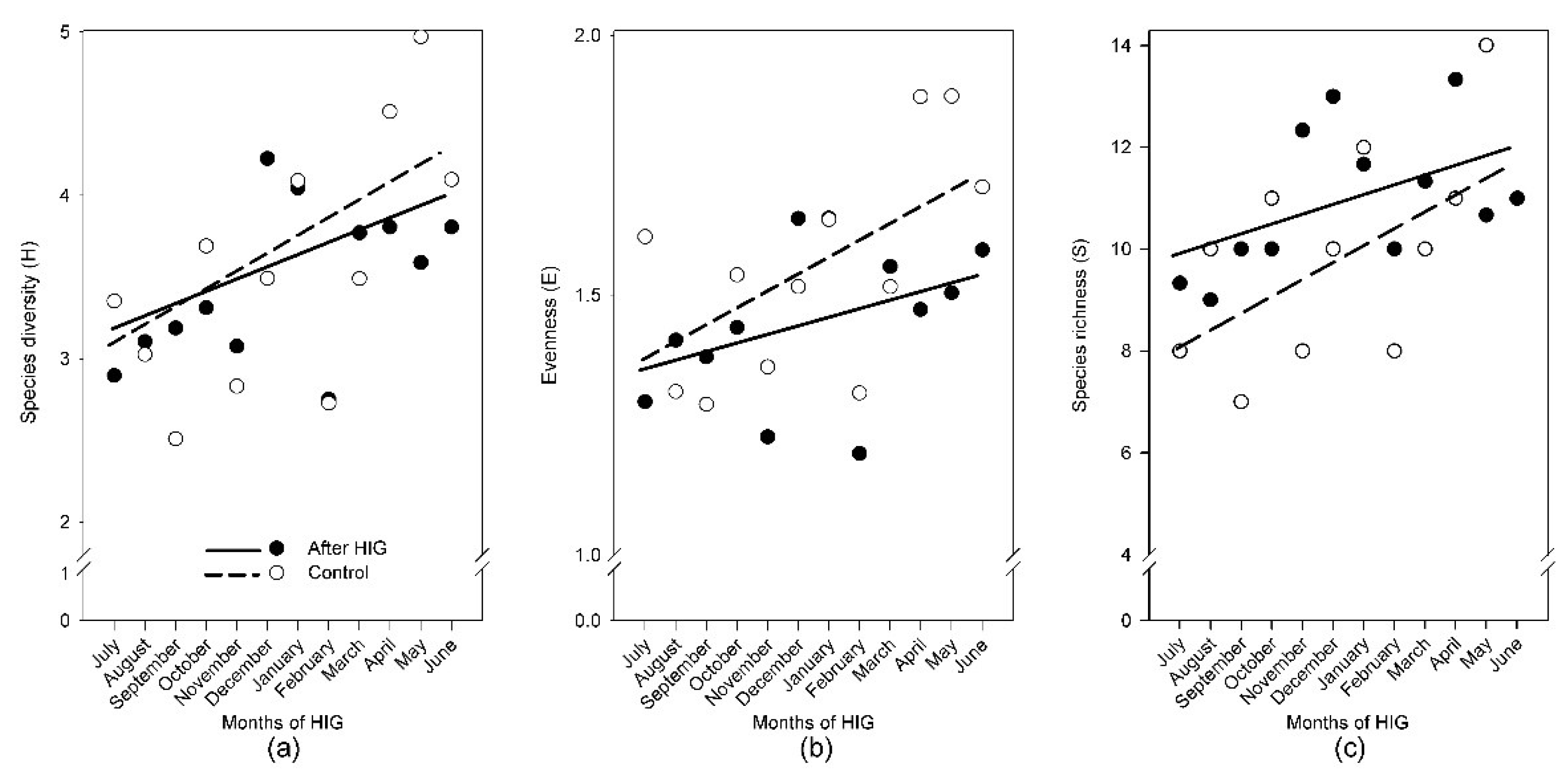
| Impact Season | Impact Month | Days after HIG | SGB (%) | SDB (%) | C3 (%) | C4 (%) | Monocots (%) | Dicots (%) | Annual Species (%) | Perennial Species (%) | S | H | E | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | - | 25.0 ± 8.3 | d | 66.7 ± 13.0 | a | 12.3 ± 4.6 | ab | 12.7 ± 4.6 | a | 17.4 ± 1.7 | a | 7.6 ± 1.7 | ns | 1.1 ± 1.0 | ns | 23.9 ± 1.0 | a | 48.0 ± 6.9 | ns | 2.6 ± 0.4 | ns | 0.66 ± 0.06 | ns | |
| Summer | February | 19 | 18.8 ± 5.9 | d | 45.1 ± 4.6 | b | 8.0 ± 5.4 | a | 10.7 ± 5.5 | a | 17.9 ± 0.6 | a | 0.9 ± 0.6 | ns | 0.0005 ± 0.001 | ns | 18.8 ± 0.0009 | a | 42.3 ± 5.4 | ns | 1.8 ± 0.3 | ns | 0.58 ± 0.07 | ns |
| January | 52 | 67.4 ± 7.8 | ac | 16.2 ± 6.1 | ce | 23.3 ± 5.9 | bcd | 43.6 ± 6.6 | bcd | 60.7 ± 4.6 | bcd | 6.7 ± 4.6 | ns | 0.7 ± 0.2 | ns | 66.7 ± 0.2 | d | 42.0 ± 5.4 | ns | 2.5 ± 0.1 | ns | 0.67 ± 0.02 | ns | |
| December | 54 | 62.8 ± 9.7 | bc | 18.5 ± 8.0 | cd | 32.1 ± 18.5 | d | 30.6 ± 18.6 | b | 45.8 ± 18.7 | b | 17.0 ± 18.7 | ns | 2.1 ± 1.5 | ns | 60.7 ± 1.5 | bc | 43.0 ± 6.0 | ns | 2.4 ± 0.1 | ns | 0.63 ± 0.05 | ns | |
| Spring | November | 87 | 67.8 ± 6.5 | ac | 10.3 ± 5.2 | cf | 16.8 ± 8.6 | abc | 50.9 ± 8.8 | cd | 57.1 ± 9.1 | bcd | 10.7 ± 9.1 | ns | 2.3 ± 3.3 | ns | 65.5 ± 3.3 | cd | 43.0 ± 6.1 | ns | 2.6 ± 0.2 | ns | 0.70 ± 0.04 | ns |
| October | 116 | 59.8 ± 4.0 | c | 4.4 ± 0.8 | f | 11.4 ± 7.0 | ab | 48.4 ± 7.0 | cd | 53.6 ± 2.3 | bc | 6.2 ± 2.3 | ns | 0.8 ± 0.8 | ns | 59.0 ± 0.8 | b | 36.3 ± 5.3 | ns | 2.5 ± 0.2 | ns | 0.69 ± 0.03 | ns | |
| September | 146 | 71.6 ± 16.1 | ac | 8.4 ± 3.4 | def | 21.6 ± 3.9 | abcd | 49.8 ± 3.9 | cd | 64.6 ± 2.3 | cd | 7.0 ± 2.3 | ns | 1.0 ± 1.2 | ns | 70.6 ± 1.2 | de | 36.3 ± 5.3 | ns | 2.4 ± 0.1 | ns | 0.66 ± 0.08 | ns | |
| Winter | August | 176 | 78.2 ± 9.6 | a | 11.2 ± 5.9 | def | 31.7 ± 15.0 | cd | 46.1 ± 14.4 | cd | 57.9 ± 20.3 | bcd | 20.3 ± 20.3 | ns | 1.7 ± 2.1 | ns | 76.5 ± 2.1 | fg | 40.7 ± 7.2 | ns | 2.5 ± 0.2 | ns | 0.66 ± 0.06 | ns |
| July | 206 | 67.8 ± 4.0 | ac | 20.1 ± 7.3 | c | 27.5 ± 3.4 | cd | 40.2 ± 3.4 | bc | 56.9 ± 10.4 | cd | 10.9 ± 10.4 | ns | 2.3 ± 2.7 | ns | 65.5 ± 2.7 | cd | 44.3 ± 7.2 | ns | 2.5 ± 0.3 | ns | 0.67 ± 0.06 | ns | |
| June | 253 | 78.3 ± 1.5 | a | 5.1 ± 1.5 | ef | 20.5 ± 7.5 | abcd | 57.8 ± 7.4 | d | 64.6 ± 4.4 | cd | 13.7 ± 4.4 | ns | 5.9 ± 7.2 | ns | 72.4 ± 7.2 | ef | 51.7 ± 7.7 | ns | 2.9 ± 0.1 | ns | 0.73 ± 0.01 | ns | |
| Autumn | May | 285 | 77.6 ± 2.9 | ab | 7.0 ± 1.2 | def | 22.9 ± 6.5 | abcd | 54.6 ± 6.5 | cd | 62.1 ± 12.9 | bcd | 15.5 ± 12.9 | ns | 2.5 ± 3.4 | ns | 75.1 ± 3.4 | efg | 45.7 ± 7.8 | ns | 2.9 ± 0.3 | ns | 0.75 ± 0.03 | ns |
| April | 317 | 77.9 ± 4.1 | a | 7.8 ± 3.3 | def | 26.7 ± 10.6 | cd | 51.0 ± 10.8 | cd | 62.5 ± 13.9 | bcd | 15.4 ± 13.9 | ns | 4.0 ± 7.0 | ns | 73.9 ± 7.0 | efg | 44.0 ± 7.7 | ns | 2.7 ± 0.3 | ns | 0.71 ± 0.03 | ns | |
| March | 357 | 80.4 ± 3.2 | a | 7.2 ± 0.4 | def | 25.7 ± 10.3 | cd | 54.5 ± 10.3 | cd | 73.4 ± 4.7 | d | 7.0 ± 4.7 | ns | 1.5 ± 1.6 | ns | 78.9 ± 1.6 | g | 50.0 ± 6.5 | ns | 2.7 ± 0.2 | ns | 0.69 ± 0.05 | ns |
| Indifferent | Decrease | Increase | ||||
|---|---|---|---|---|---|---|
| Plant Functional Groups | Number of Species | % | Number of Species | % | Number of Species | % |
| Dicotyledonous | 16 | 17 | 16 | 17 | 64 | 67 |
| Monocotyledonous | 11 | 17 | 2 | 3 | 52 | 80 |
| Perennials | 24 | 18 | 13 | 10 | 95 | 72 |
| Annuals | 3 | 10 | 6 | 19 | 22 | 71 |
| C3 | 10 | 15 | 10 | 15 | 48 | 71 |
| C4 | 16 | 18 | 9 | 10 | 64 | 72 |
| CAM | 1 | 13 | - | 7 | 88 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurtz, D.B.; Giese, M.; Asch, F.; Windisch, S.H.; Goldfarb, M.C. Effects of High Impact Grazing on Species Diversity and Plant Functional Groups in Grasslands of Northern Argentina. Sustainability 2018, 10, 3153. https://doi.org/10.3390/su10093153
Kurtz DB, Giese M, Asch F, Windisch SH, Goldfarb MC. Effects of High Impact Grazing on Species Diversity and Plant Functional Groups in Grasslands of Northern Argentina. Sustainability. 2018; 10(9):3153. https://doi.org/10.3390/su10093153
Chicago/Turabian StyleKurtz, Ditmar Bernardo, Marcus Giese, Folkard Asch, Saskia Helen Windisch, and María Cristina Goldfarb. 2018. "Effects of High Impact Grazing on Species Diversity and Plant Functional Groups in Grasslands of Northern Argentina" Sustainability 10, no. 9: 3153. https://doi.org/10.3390/su10093153
APA StyleKurtz, D. B., Giese, M., Asch, F., Windisch, S. H., & Goldfarb, M. C. (2018). Effects of High Impact Grazing on Species Diversity and Plant Functional Groups in Grasslands of Northern Argentina. Sustainability, 10(9), 3153. https://doi.org/10.3390/su10093153