Effect of Sugarcane Straw and Goat Manure on Soil Nutrient Transformation and Bacterial Communities

 , ,
, ,  ,
,
Abstract
1. Introduction
2. Material and Methods
2.1. Experimental Design and Soil Sample Preparation
2.2. Physiochemical Properties of the Soil
2.3. Soil Enzymes Activity
2.4. DNA Extraction and Purification
2.5. Metagenomics Analysis of the Soil Bacteria
2.6. Statistical and Bioinformatics Analysis
3. Results
3.1. Soil Physicochemical Characteristics
3.2. Soil Enzyme Activity (SEA)
3.3. Meta-Genomic Analysis Based on 16S rDNA of All Soils
3.4. Overlying and Separate Bacterial Groups across All Soil Sample with Different Time
3.5. Correlation among Bacterial Groups and Soil Variables
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Li, F.; Chen, L.; Zhang, J.; Yin, J.; Huang, S. Bacterial community structure after long-term organic and inorganic fertilization reveals important associations between soil nutrients and specific taxa involved in nutrient transformations. Front. Microbiol. 2017, 8, 187. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Liu, X.; Liang, Y.; Niu, J.; Xiao, Y.; Gu, Y.; Ma, L.; Meng, D.; Zhang, Y.; Huang, W.; et al. Maize growth responses to soil microbes and soil properties after fertilization with different green manures. Appl. Microbiol. Biotechnol. 2017, 101, 1289–1299. [Google Scholar] [CrossRef] [PubMed]
- Ju, X.-T.; Xing, G.-X.; Chen, X.-P.; Zhang, S.-L.; Zhang, L.-J.; Liu, X.-J.; Cui, Z.-L.; Yin, B.; Christie, P.; Zhu, Z.-L. Reducing environmental risk by improving N management in intensive Chinese agricultural systems. Proc. Natl. Acad. Sci. USA 2009, 106, 3041–3046. [Google Scholar] [CrossRef] [PubMed]
- Tayyab, M.; Islam, W.; Khalil, F.; Ziqin, P.; Caifang, Z.; Arafat, Y.; Hui, L.; Rizwan, M.; Ahmad, K.; Waheed, S.; et al. Biochar: An efficient way to manage low water availability in plants. Appl. Ecol. Environ. Res. 2018, 16, 2565–2583. [Google Scholar]
- Kravchenko, A.N.; Snapp, S.S.; Robertson, G.P. Field-scale experiments reveal persistent yield gaps in low-input and organic cropping systems. Proc. Natl. Acad. Sci. USA 2017, 114, 926–931. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Chen, Y.; Yang, X.; Cui, J.; Sui, P. The effect of different organic materials amendment on soil bacteria communities in barren sandy loam soil. Environ. Sci. Pollut. Res. 2017, 24, 24019–24028. [Google Scholar] [CrossRef] [PubMed]
- Sarfraz, R.; Shakoor, A.; Abdullah, M.; Arooj, A.; Hussain, A.; Xing, S. Impact of integrated application of biochar and nitrogen fertilizers on maize growth and nitrogen recovery in alkaline calcareous soil. Soil Sci. Plant Nutr. 2017, 63, 488–498. [Google Scholar] [CrossRef]
- Das, S.; Jeong, S.T.; Das, S.; Kim, P.J. Composted cattle manure increases microbial activity and soil fertility more than composted swine manure in a submerged rice paddy. Front. Microbiol. 2017, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Cesarano, G.; De Filippis, F.; La Storia, A.; Scala, F.; Bonanomi, G. Organic amendment type and application frequency affect crop yields, soil fertility and microbiome composition. Appl. Soil Ecol. 2017, 120, 254–264. [Google Scholar] [CrossRef]
- Li, X.; Fan, P.; Zang, M.; Xing, J. Rapid Determination of Oligopeptides and Amino Acids in Soybean Protein Hydrolysates using High-Resolution Mass Spectrometry. Phytochem. Anal. 2015, 26, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Suthar, S.; Gairola, S. Nutrient recovery from urban forest leaf litter waste solids using Eisenia fetida. Ecol. Eng. 2014, 71, 660–666. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, S.; He, T.; Liu, L.; Wu, J. Response of organic carbon mineralization and microbial community to leaf litter and nutrient additions in subtropical forest soils. Soil Biol. Biochem. 2014, 71, 13–20. [Google Scholar] [CrossRef]
- Fanin, N.; Bertrand, I. Aboveground litter quality is a better predictor than belowground microbial communities when estimating carbon mineralization along a land-use gradient. Soil Biol. Biochem. 2016, 94, 48–60. [Google Scholar] [CrossRef]
- Hristovski, S.; Berg, B.; Melovski, L. Limitless decomposition in leaf litter of Common beech: Patterns, nutrients’ and heavy metal’s dynamics. Pedobiologia 2014, 57, 131–138. [Google Scholar] [CrossRef]
- Sun, H.; Wang, Q.X.; Liu, N.; Li, L.; Zhang, C.G.; Liu, Z.B.; Zhang, Y.Y. Effects of different leaf litters on the physicochemical properties and bacterial communities in Panax ginseng-growing soil. Appl. Soil Ecol. 2017, 111, 17–24. [Google Scholar] [CrossRef]
- Prosser, J.I.; Bohannan, B.J.M.; Curtis, T.P.; Ellis, R.J.; Firestone, M.K.; Freckleton, R.P.; Green, J.L.; Green, L.E.; Killham, K.; Lennon, J.J. The role of ecological theory in microbial ecology. Nat. Rev. Microbiol. 2007, 5, 384–392. [Google Scholar] [CrossRef] [PubMed]
- He, J.-Z.; Zheng, Y.; Chen, C.-R.; He, Y.-Q.; Zhang, L.-M. Microbial composition and diversity of an upland red soil under long-term fertilization treatments as revealed by culture-dependent and culture-independent approaches. J. Soils Sediments 2008, 8, 349–358. [Google Scholar] [CrossRef]
- Shen, J.-P.; Zhang, L.-M.; Guo, J.-F.; Ray, J.L.; He, J.-Z. Impact of long-term fertilization practices on the abundance and composition of soil bacterial communities in Northeast China. Appl. Soil Ecol. 2010, 46, 119–124. [Google Scholar] [CrossRef]
- Giacometti, C.; Demyan, M.S.; Cavani, L.; Marzadori, C.; Ciavatta, C.; Kandeler, E. Chemical and microbiological soil quality indicators and their potential to differentiate fertilization regimes in temperate agroecosystems. Appl. Soil Ecol. 2013, 64, 32–48. [Google Scholar] [CrossRef]
- Peruzzi, E.; Franke-Whittle, I.H.; Kelderer, M.; Ciavatta, C.; Insam, H. Microbial indication of soil health in apple orchards affected by replant disease. Appl. Soil Ecol. 2017, 119, 115–127. [Google Scholar] [CrossRef]
- Shendure, J.; Ji, H. Next-generation DNA sequencing. Nat. Biotechnol. 2008, 26, 1135–1145. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xin, L.; Liu, J.; Yuan, M.; Liu, S.; Jiang, W.; Chen, J.; Chen, Y.; Xin, L.; Liu, J.; et al. Changes in bacterial community of soil induced by long-term straw returning. Sci. Agric. 2017, 74, 349–356. [Google Scholar] [CrossRef]
- Scotti, R.; Bonanomi, G.; Scelza, R.; Zoina, A.; Rao, M.A. Organic amendments as sustainable tool to recovery fertility in intensive agricultural systems. J. Soil Sci. Plant Nutr. 2015, 15, 333–352. [Google Scholar] [CrossRef]
- Wang, J.; Song, Y.; Ma, T.; Raza, W.; Li, J.; Howland, J.G.; Huang, Q.; Shen, Q. Impacts of inorganic and organic fertilization treatments on bacterial and fungal communities in a paddy soil. Appl. Soil Ecol. 2017, 112, 42–50. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, Q.; Zhou, J.; Wei, Q. Pyrosequencing technology reveals the impact of different manure doses on the bacterial community in apple rhizosphere soil. Appl. Soil Ecol. 2014, 78, 28–36. [Google Scholar] [CrossRef]
- Pansu, M.; Gautheyrou, J. Handbook of Soil Analysis: Mineralogical, Organic and Inorganic Methods; Springer Science & Business Media: Berlin, Germany, 2007; ISBN 3540312110. [Google Scholar]
- Li, Z.G.; Luo, Y.M.; Teng, Y. Research Methods of Soil and Environmental Microbiology; Science Press: Beijing, China, 2008. [Google Scholar]
- Eivazi, F.; Tabatabai, M.A. Glucosidases and galactosidases in soils. Soil Biol. Biochem. 1988, 20, 601–606. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pẽa, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Saqib, H.S.A.; You, M.; Gurr, G.M. Multivariate ordination identifies vegetation types associated with spider conservation in brassica crops. PeerJ 2017, 5, e3795. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Gu, T.; Wang, W.; Zhang, B.; Lin, X.; Huang, Q.; Shen, W. The effects of mineral fertilizer and organic manure on soil microbial community and diversity. Plant Soil 2010, 326, 511–522. [Google Scholar] [CrossRef]
- Deng, S.P.; Parham, J.A.; Hattey, J.A.; Babu, D. Animal manure and anhydrous ammonia amendment alter microbial carbon use efficiency, microbial biomass, and activities of dehydrogenase and amidohydrolases in semiarid agroecosystems. Appl. Soil Ecol. 2006, 33, 258–268. [Google Scholar] [CrossRef]
- Han, S.H.; An, J.Y.; Hwang, J.; Kim, S.B.; Park, B.B. The effects of organic manure and chemical fertilizer on the growth and nutrient concentrations of yellow poplar (Liriodendron tulipifera Lin.) in a nursery system. Forest Sci. Technol. 2016, 12, 137–143. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Nannipieri, P.; Giagnoni, L.; Renella, G.; Puglisi, E.; Ceccanti, B.; Masciandaro, G.; Fornasier, F.; Moscatelli, M.C.; Marinari, S. Soil enzymology: Classical and molecular approaches. Biol. Fertil. Soils 2012, 48, 743–762. [Google Scholar] [CrossRef]
- Burns, R.G.; DeForest, J.L.; Marxsen, J.; Sinsabaugh, R.L.; Stromberger, M.E.; Wallenstein, M.D.; Weintraub, M.N.; Zoppini, A. Soil enzymes in a changing environment: Current knowledge and future directions. Soil Biol. Biochem. 2013, 58, 216–234. [Google Scholar] [CrossRef]
- Mikanová, O.; Šimon, T.; Kopecký, J.; Ságová-Marečková, M. Soil biological characteristics and microbial community structure in a field experiment. Open Life Sci. 2015, 10, 249–259. [Google Scholar] [CrossRef]
- Francioli, D.; Schulz, E.; Lentendu, G.; Wubet, T.; Buscot, F.; Reitz, T. Mineral vs. organic amendments: Microbial community structure, activity and abundance of agriculturally relevant microbes are driven by long-term fertilization strategies. Front. Microbiol. 2016, 7, 1446. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, M.; Frey, B.; Mayer, J.; Ma, P. Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 2015, 9, 1177–1194. [Google Scholar] [CrossRef] [PubMed]
- Newton, R.J.; Mcmahon, K.D. Seasonal differences in bacterial community composition following nutrient additions in a eutrophic lake. Environ. Microbiol. 2011, 13, 887–899. [Google Scholar] [CrossRef] [PubMed]
- Shanks, O.C.; Kelty, C.A.; Archibeque, S.; Jenkins, M.; Newton, R.J.; McLellan, S.L.; Huse, S.M.; Sogin, M.L. Community structures of fecal bacteria in cattle from different animal feeding operations. Appl. Environ. Microbiol. 2011, 77, 2992–3001. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Martínez, V.; Dowd, S.; Sun, Y.; Allen, V. Tag-encoded pyrosequencing analysis of bacterial diversity in a single soil type as affected by management and land use. Soil Biol. Biochem. 2008, 40, 2762–2770. [Google Scholar] [CrossRef]
- Sauvadet, M.; Chauvat, M.; Cluzeau, D.; Maron, P.A.; Villenave, C.; Bertrand, I. The dynamics of soil micro-food web structure and functions vary according to litter quality. Soil Biol. Biochem. 2016, 95, 262–274. [Google Scholar] [CrossRef]
- Herzog, S.; Wemheuer, F.; Wemheuer, B.; Daniel, R. Effects of fertilization and sampling time on composition and diversity of entire and active bacterial communities in German grassland soils. PLoS ONE 2015, 10, e0145575. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Zhang, X.X.; Guo, X.; Wang, D.; Chu, H. Bacterial diversity in soils subjected to long-term chemical fertilization can be more stably maintained with the addition of livestock manure than wheat straw. Soil Biol. Biochem. 2015, 88, 9–18. [Google Scholar] [CrossRef]
- Bonanomi, G.; De Filippis, F.; Cesarano, G.; La Storia, A.; Ercolini, D.; Scala, F. Organic farming induces changes in soil microbiota that affect agro-ecosystem functions. Soil Biol. Biochem. 2016, 103, 327–336. [Google Scholar] [CrossRef]
- Sykes, G.; Skinner, F.A. Actinomycetales: Characteristics and Practical Importance; Academic Press: Cambridge, MA, USA, 1973; ISBN 0126799504. [Google Scholar]
- Pershina, E.; Valkonen, J.; Kurki, P.; Ivanova, E.; Chirak, E.; Korvigo, I.; Provorov, N.; Andronov, E. Comparative analysis of prokaryotic communities associated with organic and conventional farming systems. PLoS ONE 2015, 10, e0145072. [Google Scholar] [CrossRef] [PubMed]
- Charbonneau, D.M.; Meddeb-Mouelhi, F.; Boissinot, M.; Sirois, M.; Beauregard, M. Identification of Thermophilic Bacterial Strains Producing Thermotolerant Hydrolytic Enzymes from Manure Compost. Indian J. Microbiol. 2012, 52, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Nagao, N.; Yamamoto, S.; Toda, T.; Kurosawa, N. Thermobacillus composti sp. nov., a moderately thermophilic bacterium isolated from a composting reactor. Int. J. Syst. Evol. Microbiol. 2007, 57, 1473–1477. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.R.; He, J. Characterization of anaerobic consortia coupled lignin depolymerization with biomethane generation. Bioresour. Technol. 2013, 139, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, T.; Nakajima, Y.; Matsuya, K.; Ikenaga, M.; Asakawa, S.; Kimura, M. Bacterial community in plant residues in a Japanese paddy field estimated by RFLP and DGGE analyses. Soil Biol. Biochem. 2007, 39, 463–472. [Google Scholar] [CrossRef]
- Tayyab, M.; Noman, A.; Islam, W.; Waheed, S.; Arafat, Y.; Ali, F.; Zaynab, M.; Lin, S.; Zhang, H.; Lin, W. Bioethanol production from lignocellulosic biomass by environment-friendly pretreatment methods: A review. Appl. Ecol. Environ. Res. 2018, 16, 225–249. [Google Scholar] [CrossRef]
- Zhang, H.B.; Yang, M.X.; Tu, R. Unexpectedly high bacterial diversity in decaying wood of a conifer as revealed by a molecular method. Int. Biodeterior. Biodegrad. 2008, 62, 471–474. [Google Scholar] [CrossRef]
- Mori, K.; Kamagata, Y. The challenges of studying the anaerobic microbial world. Microbes Environ. 2014, 29, 335–337. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, R.; Gao, J.; Wang, X.; Fan, F.; Ma, X.; Yin, H.; Zhang, C.; Feng, K.; Deng, Y. Thirty-one years of rice-rice-green manure rotations shape the rhizosphere microbial community and enrich beneficial bacteria. Soil Biol. Biochem. 2017, 104, 208–217. [Google Scholar] [CrossRef]
- Arafat, Y.; Wei, X.; Jiang, Y.; Chen, T.; Saqib, H.S.A.; Lin, S.; Lin, W. Spatial distribution patterns of root-associated bacterial communities mediated by root exudates in different aged ratooning tea monoculture systems. Int. J. Mol. Sci. 2017, 18, 1–16. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
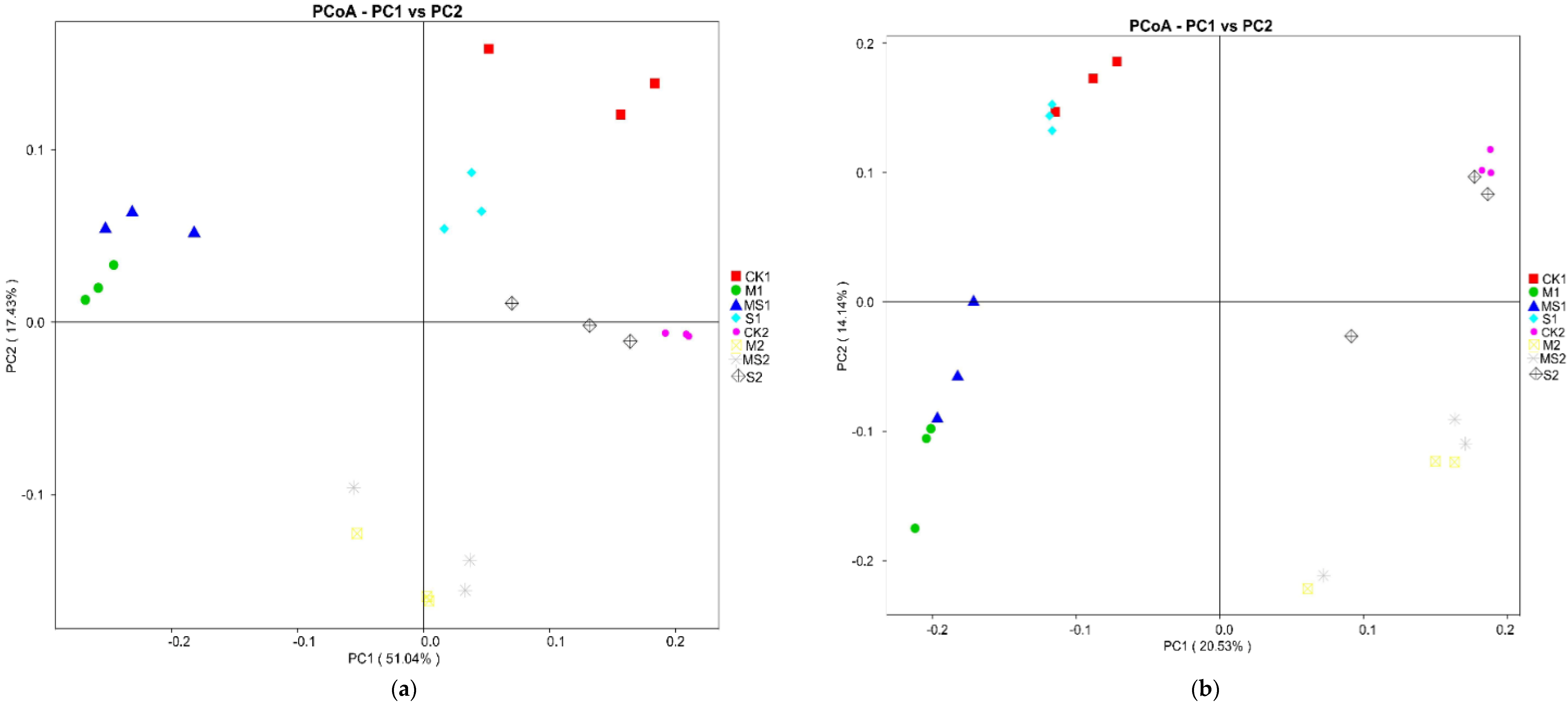{kind=link}
{kind=link}
{kind=link}
| Treatments | pH | AP | TP | AK | TN | AN | NH4+ | NO3− | TC | C/N |
|---|---|---|---|---|---|---|---|---|---|---|
| g/kg | g/kg | mg/kg | g/kg | mg/kg | mg/kg | mg/kg | g/kg | |||
| CK1 | 5.64 ± 0.01 f | 0.09 ± 0.00 e | 0.70 ± 0.07 bc | 71.85 ± 3.140 e | 1.12 ± 0.07 f | 113.05 ± 1.01 cd | 2.79 ± 0.13 d | 21.85 ± 1.10 b | 9.96 ± 0.10 g | 8.95 ± 0.660 e |
| M1 | 5.72 ± 0.03 e | 0.20 ± 0.00 c | 0.88 ± 0.08 b | 865.92 ± 3.02 c | 2.50 ± 0.02 c | 307.30 ± 2.02 a | 9.49 ± 0.52 a | 29.46 ± 0.37 a | 32.71 ± 0.36 c | 13.09 ± 0.22 a |
| MS1 | 7.51 ± 0.01 b | 0.25 ± 0.01 b | 1.27 ± 0.04 a | 1008.41 ± 6.18 b | 2.29 ± 0.02 d | 207.55 ± 9.09 b | 9.79 ± 0.26 a | 20.16 ± 0.12 bc | 27.32 ± 0.07 d | 11.91 ± 0.06 b |
| S1 | 6.10 ± 0.02 d | 0.10 ± 0.00 d | 0.71 ± 0.00 bc | 151.46 ± 18.09 d | 1.11 ± 0.01 f | 113.05 ± 5.05 cd | 5.90 ± 0.09 c | 19.53 ± 0.49 bc | 12.69 ± 0.03 f | 11.47 ± 0.09 bc |
| CK2 | 5.73 ± 0.02 e | 0.09 ± 0.00 de | 0.61 ± 0.01 c | 65.73 ± 6.28 e | 0.93 ± 0.01 g | 79.80 ± 4.040 d | 2.10 ± 0.05 d | 18.89 ± 0.36 bc | 10.16 ± 0.06 g | 10.89 ± 0.08 cd |
| M2 | 6.80 ± 0.02 c | 0.39 ± 0.00 a | 1.34 ± 0.07 a | 1138.21 ± 3.09 a | 4.59 ± 0.02 a | 327.25 ± 5.46 a | 6.90 ± 0.33 bc | 32.41 ± 0.12 a | 47.50 ± 1.00 a | 10.35 ± 0.19 d |
| MS2 | 7.61 ± 0.02 a | 0.25 ± 0.00 b | 1.42 ± 0.11 a | 980.61 ± 3.03 b | 3.17 ± 0.11 b | 222.95 ± 15.16 b | 7.71 ± 0.12 b | 21.00 ± 0.85 bc | 36.70 ± 1.26 b | 11.59 ± 0.02 bc |
| S2 | 6.15 ± 0.02 d | 0.09 ± 0.00 de | 0.75 ± 0.04 bc | 129.03 ± 6.13 d | 1.31 ± 0.01 e | 136.50 ± 5.66 c | 6.18 ± 1.00 c | 18.05 ± 0.12 c | 16.67 ± 0.09 e | 12.72 ± 0.10 a |
| Treatment | Urease (NH3-N, μg g−1, 24 h, 37 °C) | Neutral Phosphatase (Phenol, nmol g−1, 24 h, 37 °C) | β-Glucosidase (p-Nitrophenol, μmol g−1, 24 h, 37 °C) | Cellulase (Glucose, mg g−1, 24 h, 37 °C) |
|---|---|---|---|---|
| CK1 | 14.09 ± 1.09 c | 2.71 ± 0.13 b | 18.34 ± 0.35 bc | 11.65 ± 1.24 bc |
| M1 | 31.79 ± 0.08 a | 1.58 ± 0.03 d | 28.12 ± 1.95 a | 11.72 ± 0.68 bc |
| MS1 | 25.57 ± 0.08 b | 1.09 ± 0.06 e | 23.54 ± 2.22 abc | 11.48 ± 0.04 bc |
| S1 | 12.71 ± 0.50 c | 2.50 ± 0.04 bc | 20.42 ± 0.49 bc | 10.52 ± 0.20 c |
| CK2 | 14.60 ± 0.21 c | 2.30 ± 0.12 c | 17.92 ± 2.57 c | 10.58 ± 0.24 c |
| M2 | 26.20 ± 0.04 b | 1.91 ± 0.01 d | 18.06 ± 1.60 c | 16.08 ± 2.02 a |
| MS2 | 25.61 ± 1.85 b | 1.65 ± 0.02 d | 24.92 ± 1.11 ab | 16.08 ± 0.22 a |
| S2 | 13.51 ± 1.35 c | 3.56 ± 0.24 a | 20.42 ± 3.68 bc | 14.15 ± 0.29 ab |
| Environmental Variable | r-Value |
|---|---|
| pH + TC | 0.5407 |
| pH + C:N | 0.2881 |
| pH + NH4+ | 0.5266 |
| pH + TN | 0.5005 |
| pH + NO3− | 0.414 |
| AP + TP + AK | 0.5898 |
| NO3− + NH4+ | 0.5971 |
| TN + TC | 0.6033 |
| NO3− + NH4+ + TN + AN | 0.6564 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tayyab, M.; Islam, W.; Arafat, Y.; Pang, Z.; Zhang, C.; Lin, Y.; Waqas, M.; Lin, S.; Lin, W.; Zhang, H. Effect of Sugarcane Straw and Goat Manure on Soil Nutrient Transformation and Bacterial Communities. Sustainability 2018, 10, 2361. https://doi.org/10.3390/su10072361
Tayyab M, Islam W, Arafat Y, Pang Z, Zhang C, Lin Y, Waqas M, Lin S, Lin W, Zhang H. Effect of Sugarcane Straw and Goat Manure on Soil Nutrient Transformation and Bacterial Communities. Sustainability. 2018; 10(7):2361. https://doi.org/10.3390/su10072361
Chicago/Turabian StyleTayyab, Muhammad, Waqar Islam, Yasir Arafat, Ziqin Pang, Caifang Zhang, Yu Lin, Muhammad Waqas, Sheng Lin, Wenxiong Lin, and Hua Zhang. 2018. "Effect of Sugarcane Straw and Goat Manure on Soil Nutrient Transformation and Bacterial Communities" Sustainability 10, no. 7: 2361. https://doi.org/10.3390/su10072361
APA StyleTayyab, M., Islam, W., Arafat, Y., Pang, Z., Zhang, C., Lin, Y., Waqas, M., Lin, S., Lin, W., & Zhang, H. (2018). Effect of Sugarcane Straw and Goat Manure on Soil Nutrient Transformation and Bacterial Communities. Sustainability, 10(7), 2361. https://doi.org/10.3390/su10072361