Comparative Sequence Analysis of Hsp70 Gene from Mytilus edulis desolationis and Aulacomya ater of the Kerguelen Islands †

Introduction
Materials and Methods
Collection of specimens
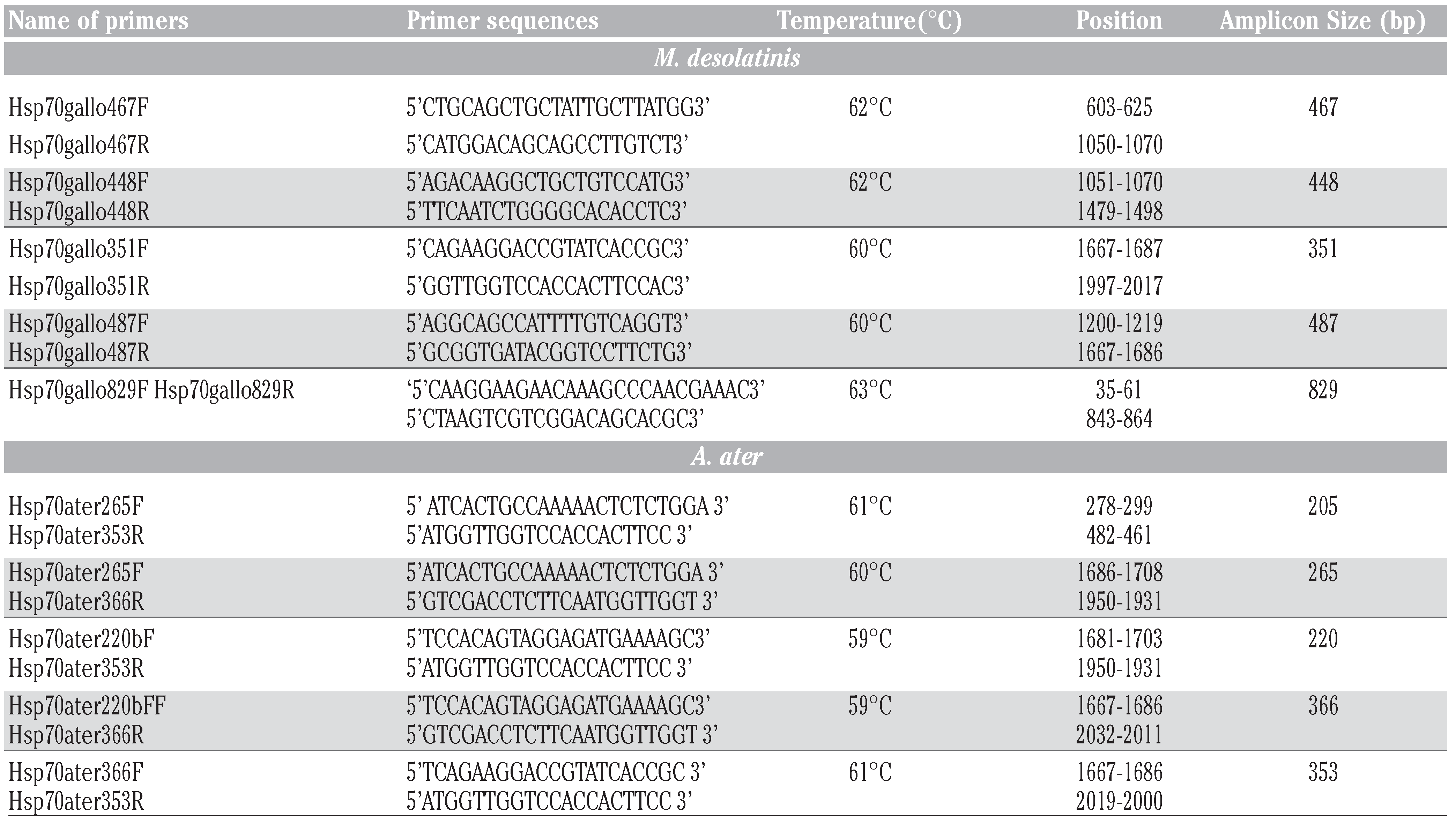
RT-PCR and sequencing
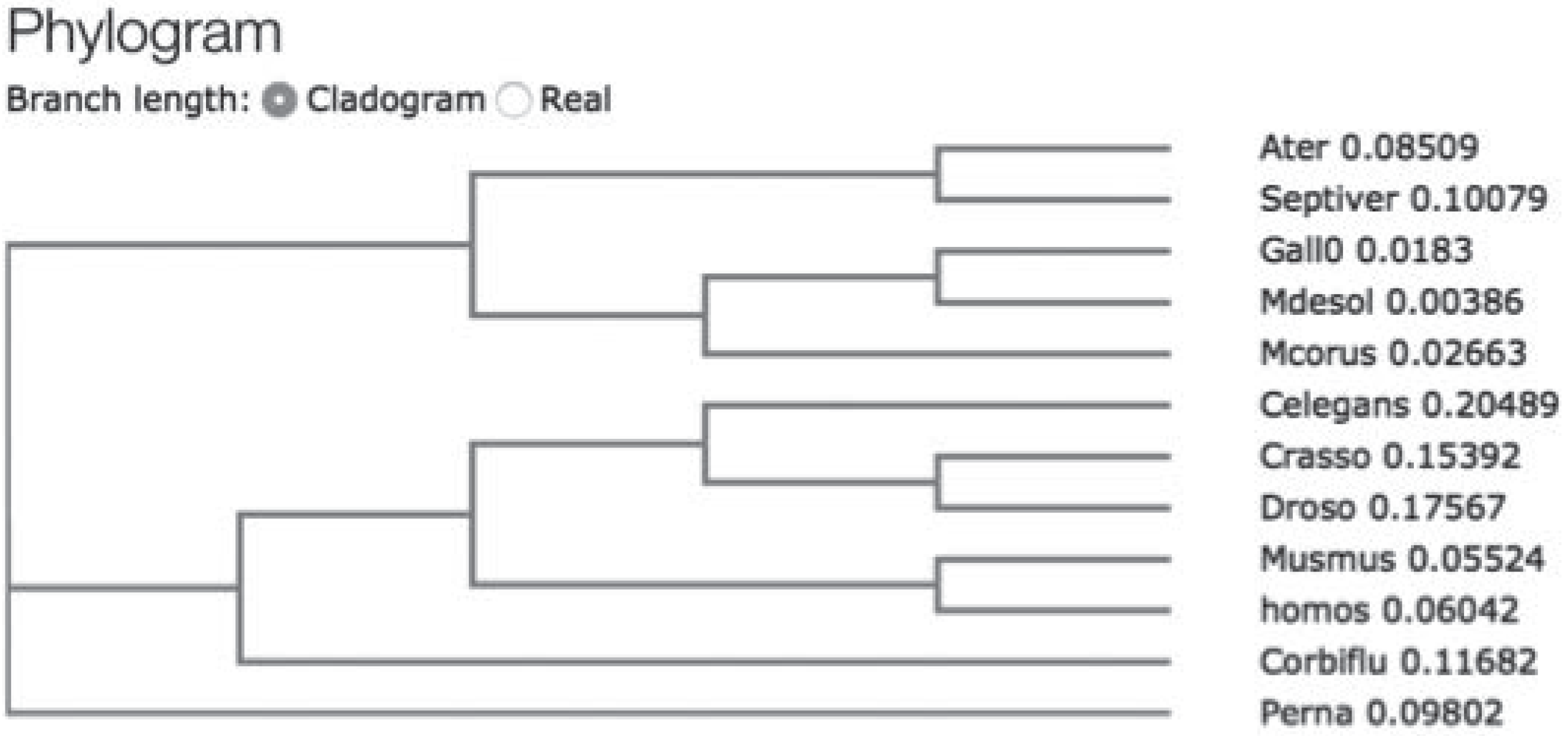
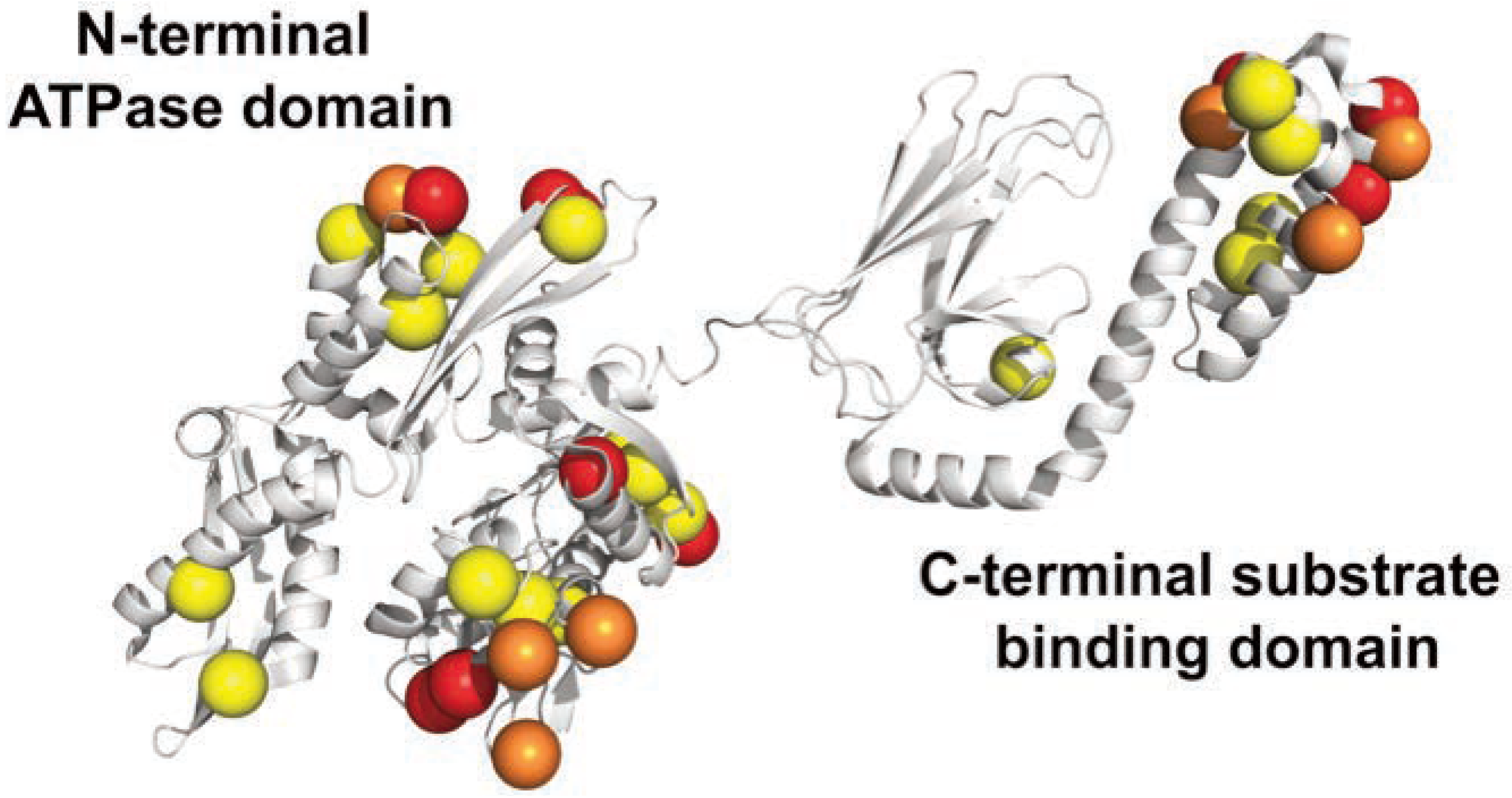
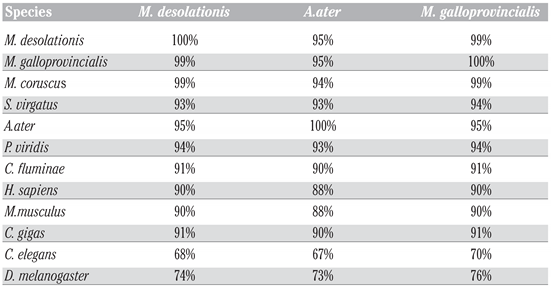
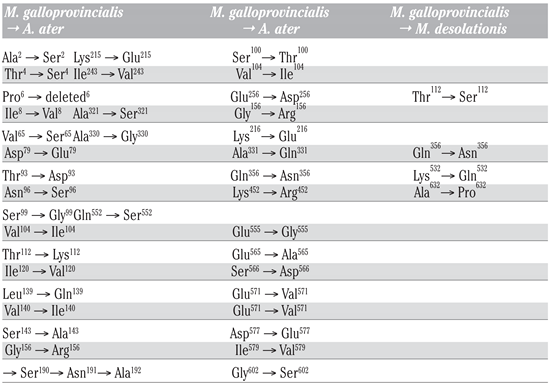
Results
Discussion
Conclusions
Funding
Acknowledgments
References
- Goldberg, E.D. The mussel watch - a first step in global marine monitoring. Mar. Poll. Bull. 1975, 6, 111. [Google Scholar] [CrossRef]
- Robinson, C.D.; Webster, L.; Martínez-Gómez, C.; Burgeot, T.; Gubbins, M.J.; Thain, J.E.; et al. Assessment of contaminant concentrations in sediments, fish and mussels sampled from the North Atlantic and European regional seas within the ICON project. Mar. Environ. Res. 2017, 124, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Oliva, A.L.; Ovaert, J.; Arias, A.H.; Souissi, S.; Marcovecchio, J.E. Mussels as bioindicators of PAHs pollution within Argentinean Coastal Environments, South America. Int. J. Environ. Res. 2015, 9, 1293–1304. [Google Scholar]
- Philipp, E.E.; Kraemer, L.; Melzner, F.; Poustka, A.J.; Thieme, S.; Findeisen, U.; et al. Massively parallel RNA sequencing identifies a complex immune gene repertoire in the lophotrochozoan Mytilus edulis. PLoS ONE 2012, 7, e33091. [Google Scholar] [CrossRef] [PubMed]
- Poynton, H.C.; Robinson, W.E.; Blalock, B.J.; Hannigan, R.E. Correlation of transcriptomic responses and metal bioaccumulation in Mytilus edulis L. reveals early indicators of stress. Aqua Toxicol. 2014, 155, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Campos, A.; Apraiz, I.; da Fonseca, R.R.; Cristobal, S. Shotgun analysis of the marine mussel Mytilus edulis hemolymph proteome and mapping the innate immunity elements. Proteomics 2015, 15, 4021–4029. [Google Scholar] [CrossRef] [PubMed]
- Caza, F.; Cledon, M.; St-Pierre, Y. Biomonitoring climate change and pollution in marine ecosystems: A review on Aulacomya ater. J. Mar. Biol. 2016, e2016. [Google Scholar] [CrossRef]
- Caza, F.; Betoulle, S.; Auffret, M.; Brousseau, P.; Fournier, M.; St-Pierre, Y. Comparative analysis of hemocyte properties from Mytilus edulis desolationis and Aulacomya ater in the Kerguelen Islands. Mar. Environ. Res. 2015, 110, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Lebouvier, M.; Laparie, M.; Hulle, M.; Marais, A.; Cozic, Y.; Lalouette, L.; et al. The significance of the sub-Antarctic Kerguelen Islands for the assessment of the vulnerability of native communities to climate change, alien insect invasions and plant viruses. Biol. Invasions 2011, 13, 1195–1208. [Google Scholar] [CrossRef]
- Feder, M.E.; Hofmann, G.E. Heat shock proteins, molecular chaperones and their stress response: Evolutionary and ecological physiology. Annu. Rev. Physiol. 1999, 61, 243–282. [Google Scholar] [CrossRef] [PubMed]
- Gross, M. Emergency services: A bird’s eye perspective on the many different functions of stress proteins. Curr. Protein Pept. Sci. 2004, 5, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Snyder, M.J.; Girvetz, E.; Mulder, E.P. Induction of marine mollusc stress proteins by chemical or physical stress. Arch. Environ. Contam. Toxicol. 2001, 41, 22–29. [Google Scholar] [PubMed]
- Bodin, N.; Burgeot, T.; Stanisiere, J.Y.; Bocquené, G.; Menard, D.; Minier, C.; et al. Seasonal variations of a battery of biomarkers and physiological indices for the mussel Mytilus galloprovincialis transplanted into the northwest Mediterranean Sea. Comp Biochem Physiol Part C: Toxicol. Pharmacol. 2004, 138, 411–427. [Google Scholar]
- Hamer, B.; Hamer, D.P.; Müller, W.E.; Batel, R. Stress-70 proteins in marine mussel Mytilus galloprovincialis as biomarkers of environmental pollution: A field study. Environ. Int. 2004, 30, 873–882. [Google Scholar] [CrossRef]
- Clark, M.S.; Peck, L.S. HSP70 heat shock proteins and environmental stress in Antarctic marine organisms: A minireview. Mar. Genomics 2009, 2, 11–18. [Google Scholar] [CrossRef]
- Miovi, V.; Bulog, A.; Kui, N.; Jakovac, H.; Radoševi-Staši, B. Metallothioneins and heat shock proteins 70 in marine mussels as sensors of environmental pollution in Northern Adriatic Sea. Env. Toxicol. Pharmacol. 2009, 28, 439–447. [Google Scholar]
- Clerico, E.M.; Tilitsky, J.M.; Meng, W.; Gierasch, L.M. How hsp70 molecular machines interact with their substrates to mediate diverse physiological functions. J. Mol. Biol. 2015, 427, 1575–1588. [Google Scholar] [CrossRef] [PubMed]
- Rippmann, F.; Taylor, W.R.; Rothbard, J.B.; Green, N.M. A hypothetical model for the peptide binding domain of hsp70 based on the peptide binding domain of HLA. EMBO J. 1991, 10, 1053–1059. [Google Scholar] [CrossRef] [PubMed]
- Mack, K.L.; Shorter, J. Engineering and evolution of molecular chaperones and protein disaggregases with enhanced activity. Front. Mol. Biosci. 2016, e3. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
©Copyright F. Caza et al., 2016 Licensee PAGEPress, Italy. This work is licensed under a Creative Commons Attribution NonCommercial 4.0 License (CC BYNC 4.0).
Share and Cite
Caza, F.; Betoulle, S.; Auffret, M.; Brousseau, P.; Fournier, M.; St-Pierre, Y. Comparative Sequence Analysis of Hsp70 Gene from Mytilus edulis desolationis and Aulacomya ater of the Kerguelen Islands. J. Xenobiot. 2016, 6, 6721. https://doi.org/10.4081/xeno.2016.6721
Caza F, Betoulle S, Auffret M, Brousseau P, Fournier M, St-Pierre Y. Comparative Sequence Analysis of Hsp70 Gene from Mytilus edulis desolationis and Aulacomya ater of the Kerguelen Islands. Journal of Xenobiotics. 2016; 6(2):6721. https://doi.org/10.4081/xeno.2016.6721
Chicago/Turabian StyleCaza, F., S. Betoulle, M. Auffret, P. Brousseau, M. Fournier, and Y. St-Pierre. 2016. "Comparative Sequence Analysis of Hsp70 Gene from Mytilus edulis desolationis and Aulacomya ater of the Kerguelen Islands" Journal of Xenobiotics 6, no. 2: 6721. https://doi.org/10.4081/xeno.2016.6721
APA StyleCaza, F., Betoulle, S., Auffret, M., Brousseau, P., Fournier, M., & St-Pierre, Y. (2016). Comparative Sequence Analysis of Hsp70 Gene from Mytilus edulis desolationis and Aulacomya ater of the Kerguelen Islands. Journal of Xenobiotics, 6(2), 6721. https://doi.org/10.4081/xeno.2016.6721