
Effect of Retinoic Acid on the Cerebral Vasculature: Analysis of the Vasoactive Response of Smooth Muscle Cells in Normal and Ischemic Contexts

 ,
,  ,
,  , and
, and 
Abstract

1. Introduction
2. Materials and Methods
2.1. Animals and Ethical Issues
2.2. Smooth Muscle Cell Culture
2.3. Ischemic Stroke Simulation
2.3.1. Primary Cortical Cultures
2.3.2. OGD and Reperfusion
2.4. PCSA Contractility Experiments in SMCs
- (1)
- Non-genomic effect of RA on vasculature: The rapid action of 10 µM of RA was analyzed in SMCs pre-contracted with noradrenaline (NA; 1 µM). Controls with the vehicle used to dissolve RA (0.1% ethanol) were always performed. Vascular rapid effects are described as the immediate effects that occur after application of a specific drug on SMCs [25].
- (2)
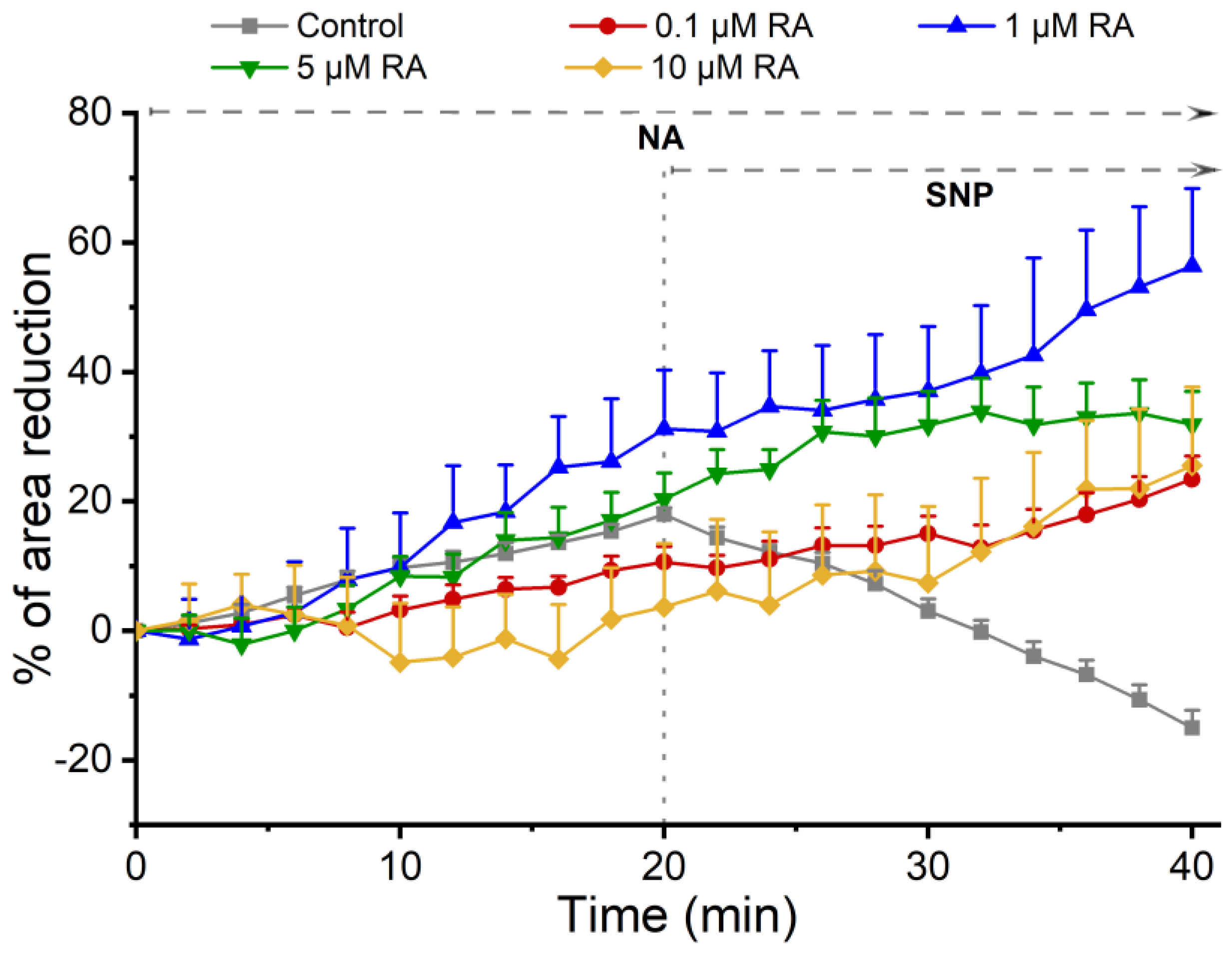
- Genomic effect of RA on vasculature: For this analysis, SMCs were pre-incubated for 24 h with 0.1% ethanol (control) or with RA at different concentrations of RA (0.1 µM, 1 µM, 5 µM, and 10 µM). Incubation for 24 h is adequate for genomic effects on contractility patterns of SMCs to occur [13]. Then, the vasoactive response of SMCs was assessed using the contractile agent noradrenaline (NA; 1 µM) and the relaxing agent sodium nitroprusside (SNP; 1 µM). Each experiment lasted 60 min. After the first observation, an image of the cells was recorded. For the first 20 min, the cells were kept on the microscope without the addition of any agent. Afterwards, NA was added, and changes in cell area were recorded for 20 min. Then, SNP was added, and changes in cell area were again recorded for 20 min. This period was necessary to obtain a maximum response of the cells to the agents used.
- (3)
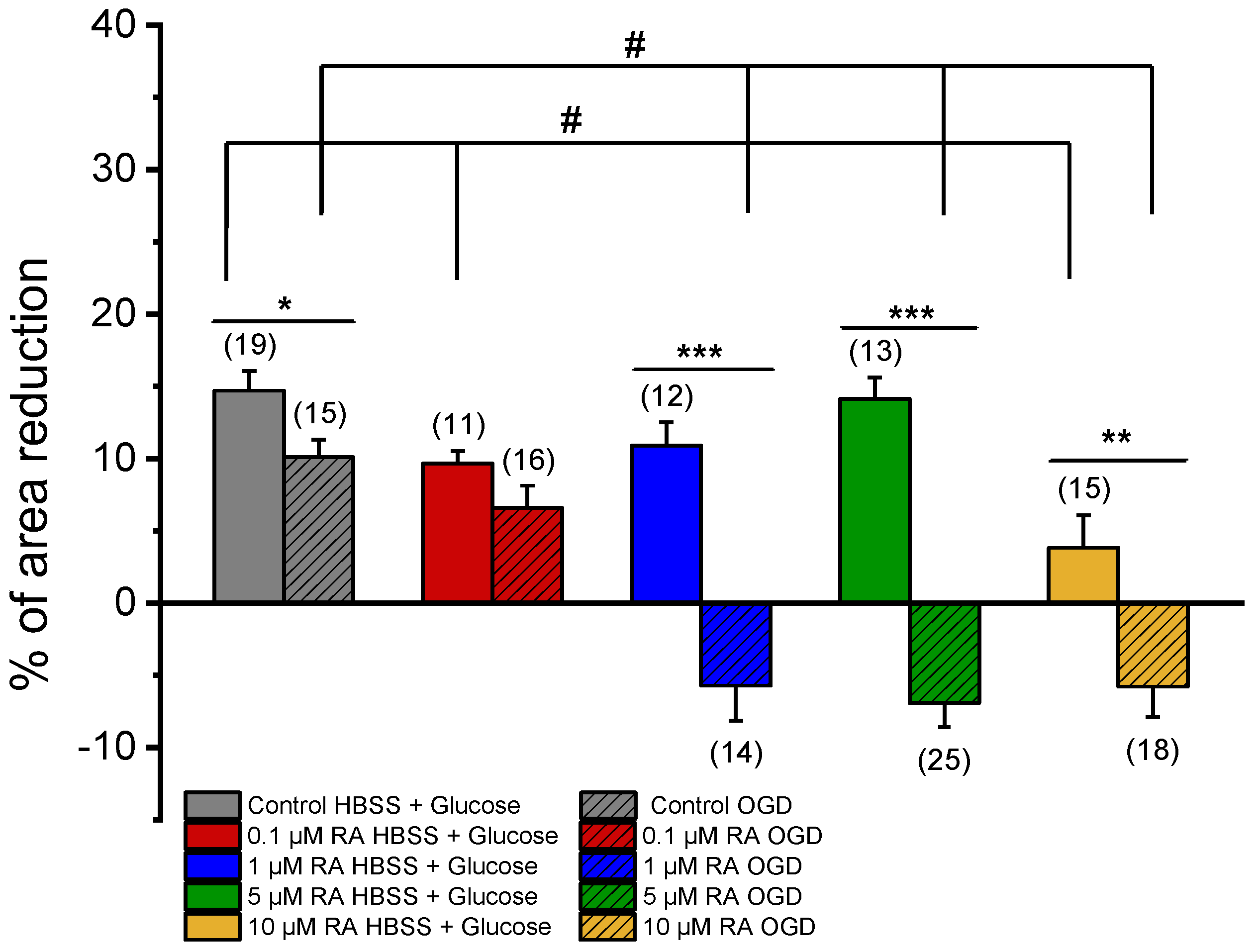
- Genomic effect of RA on ischemic events: For this analysis, SMCs were pre-incubated for 24 h with PCC secretome (Table 1). The setup strategy was the same as described in 2).
2.5. Drugs
2.6. Statistical Analysis
3. Results
3.1. Evaluation of the Non-Genomic (Rapid) Effects of RA on SMCs
3.2. Evaluation of the Genomic (Long) Effects of RA on SMCs
3.3. Effect of RA on the Vasoactive Response Regulation Conditioned by Neurons and Astrocytes
3.3.1. SMC Vascular Response to NA Contraction
3.3.2. SMC Vascular Response to the NO Effect
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SMC | Smooth muscle cells |
| RA | Retinoic acid |
| NVU | Neurovascular unit |
| EC | Endothelial cells |
| BBB | Blood–brain barrier |
| CBF | Cerebral blood flow |
| CNS | Central nervous system |
| NO | Nitric oxide |
| RAR | Retinoic acid receptor |
| RXR | Retinoic X receptor |
| MCA | Middle cerebral artery |
| PBS | Phosphate-buffered saline |
| CCM | Complete culture medium |
| FBS | Fetal bovine serum |
| FGF | Fibroblast growth factor |
| NBM | Neurobasal medium |
| OGD | Oxygen and glucose deprivation |
| HBSS | Hank’s buffered saline solution |
| PCSA | Planar cell surface area |
| NA | Noradrenaline |
| SNP | Sodium nitroprusside |
| sGC | Soluble guanylate cyclase |
References
- Muoio, V.; Persson, P.B.; Sendeski, M.M. The neurovascular unit—Concept review. Acta Physiol. 2014, 210, 790–798. [Google Scholar] [CrossRef]
- Mergenthaler, P.; Lindauer, U.; Dienel, G.A.; Meisel, A. Sugar for the brain: The role of glucose in physiological and pathological brain function. Trends Neurosci. 2013, 36, 587–597. [Google Scholar] [CrossRef]
- Iadecola, C. The Neurovascular Unit Coming of Age: A Journey through Neurovascular Coupling in Health and Disease. Neuron 2017, 96, 17–42. [Google Scholar] [CrossRef]
- Sweeney, M.D.; Ayyadurai, S.; Zlokovic, B.V. Pericytes of the neurovascular unit: Key functions and signaling pathways. Nat. Neurosci. 2016, 19, 771–783. [Google Scholar] [CrossRef]
- Yu, X.; Ji, C.; Shao, A. Neurovascular Unit Dysfunction and Neurodegenerative Disorders. Front. Neurosci. 2020, 14, 334. [Google Scholar] [CrossRef] [PubMed]
- Keaney, J.; Campbell, M. The dynamic blood-brain barrier. FEBS J. 2015, 282, 4067–4079. [Google Scholar] [CrossRef] [PubMed]
- Poittevin, M.; Lozeron, P.; Hilal, R.; Levy, B.I.; Merkulova-Rainon, T.; Kubis, N. Smooth muscle cell phenotypic switching in stroke. Transl. Stroke Res. 2014, 5, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Rensen, S.S.; Doevendans, P.A.; van Eys, G.J. Regulation and characteristics of vascular smooth muscle cell phenotypic diversity. Neth. Heart J. 2007, 15, 100–108. [Google Scholar] [CrossRef]
- Nishida, M.; Tanaka, T.; Mangmool, S.; Nishiyama, K.; Nishimura, A. Canonical Transient Receptor Potential Channels and Vascular Smooth Muscle Cell Plasticity. J. Lipid Atheroscler 2020, 9, 124–139. [Google Scholar] [CrossRef]
- Quelhas, P.; Baltazar, G.; Cairrao, E. The Neurovascular Unit: Focus on the Regulation of Arterial Smooth Muscle Cells. Curr. Neurovasc. Res. 2019, 16, 502–515. [Google Scholar] [CrossRef]
- Kuriakose, D.; Xiao, Z. Pathophysiology and Treatment of Stroke: Present Status and Future Perspectives. Int. J. Mol. Sci. 2020, 21, 7609. [Google Scholar] [CrossRef]
- Grysiewicz, R.A.; Thomas, K.; Pandey, D.K. Epidemiology of ischemic and hemorrhagic stroke: Incidence, prevalence, mortality, and risk factors. Neurol. Clin. 2008, 26, 871–895. [Google Scholar] [CrossRef] [PubMed]
- Mariana, M.; Roque, C.; Baltazar, G.; Cairrao, E. In Vitro Model for Ischemic Stroke: Functional Analysis of Vascular Smooth Muscle Cells. Cell Mol. Neurobiol. 2021, 42, 2289–2304. [Google Scholar] [CrossRef] [PubMed]
- Machado-Pereira, M.; Santos, T.; Ferreira, L.; Bernardino, L.; Ferreira, R. Challenging the great vascular wall: Can we envision a simple yet comprehensive therapy for stroke? J. Tissue Eng. Regen. Med. 2018, 12, e350–e354. [Google Scholar] [CrossRef] [PubMed]
- Roque, C.; Baltazar, G. Impact of Astrocytes on the Injury Induced by In Vitro Ischemia. Cell Mol. Neurobiol. 2017, 37, 1521–1528. [Google Scholar] [CrossRef]
- van Neerven, S.; Regen, T.; Wolf, D.; Nemes, A.; Johann, S.; Beyer, C.; Hanisch, U.K.; Mey, J. Inflammatory chemokine release of astrocytes in vitro is reduced by all-trans retinoic acid. J. Neurochem. 2010, 114, 1511–1526. [Google Scholar] [CrossRef]
- Kampmann, E.; Johann, S.; van Neerven, S.; Beyer, C.; Mey, J. Anti-inflammatory effect of retinoic acid on prostaglandin synthesis in cultured cortical astrocytes. J. Neurochem. 2008, 106, 320–332. [Google Scholar] [CrossRef]
- Barreto, G.; White, R.E.; Ouyang, Y.; Xu, L.; Giffard, R.G. Astrocytes: Targets for neuroprotection in stroke. Cent. Nerv. Syst. Agents Med. Chem. 2011, 11, 164–173. [Google Scholar] [CrossRef]
- Olmez, I.; Ozyurt, H. Reactive oxygen species and ischemic cerebrovascular disease. Neurochem. Int. 2012, 60, 208–212. [Google Scholar] [CrossRef]
- de Hoog, E.; Lukewich, M.K.; Spencer, G.E. Retinoic acid inhibits neuronal voltage-gated calcium channels. Cell Calcium 2018, 72, 51–61. [Google Scholar] [CrossRef]
- Pouso, M.R.; Cairrao, E. Effect of retinoic acid on the neurovascular unit: A review. Brain Res. Bull. 2022, 184, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Shmarakov, I.O. Retinoid-xenobiotic interactions: The Ying and the Yang. Hepatobiliary Surg. Nutr. 2015, 4, 243–267. [Google Scholar] [CrossRef] [PubMed]
- Powała, K.; Żołek, T.; Brown, G.; Kutner, A. Molecular Interactions of Selective Agonists and Antagonists with the Retinoic Acid Receptor γ. Int. J. Mol. Sci. 2024, 25, 6568. [Google Scholar] [CrossRef]
- Grignard, E.; Håkansson, H.; Munn, S. Regulatory needs and activities to address the retinoid system in the context of endocrine disruption: The European viewpoint. Reprod. Toxicol. 2020, 93, 250–258. [Google Scholar] [CrossRef]
- Quelhas, P.; Baltazar, G.; Cairrao, E. Characterization of culture from smooth muscle cells isolated from rat middle cerebral arteries. Tissue Cell 2020, 66, 101400. [Google Scholar] [CrossRef] [PubMed]
- Sommer, C.J. Ischemic stroke: Experimental models and reality. Acta Neuropathol. 2017, 133, 245–261. [Google Scholar] [CrossRef]
- Yang, C.; Hawkins, K.E.; Dore, S.; Candelario-Jalil, E. Neuroinflammatory mechanisms of blood-brain barrier damage in ischemic stroke. Am. J. Physiol. Cell Physiol. 2019, 316, C135–C153. [Google Scholar] [CrossRef]
- Kang, J.B.; Park, D.J.; Shah, M.A.; Koh, P.O. Retinoic acid exerts neuroprotective effects against focal cerebral ischemia by preventing apoptotic cell death. Neurosci. Lett. 2021, 757, 135979. [Google Scholar] [CrossRef] [PubMed]
- Cairrão, E.; Santos-Silva, A.J.; Alvarez, E.; Correia, I.; Verde, I. Isolation and culture of human umbilical artery smooth muscle cells expressing functional calcium channels. In Vitro Cell. Dev. Biol. Anim. 2009, 45, 175–184. [Google Scholar] [CrossRef]
- Santos, T.; Ferreira, R.; Maia, J.; Agasse, F.; Xapelli, S.; Cortes, L.; Braganca, J.; Malva, J.O.; Ferreira, L.; Bernardino, L. Polymeric nanoparticles to control the differentiation of neural stem cells in the subventricular zone of the brain. ACS Nano 2012, 6, 10463–10474. [Google Scholar] [CrossRef]
- Monge, L.; Fernández, N.; Salcedo, A.; García-Villalón, A.L.; Diéguez, G. Role of alpha-adrenoceptors and prostacyclin in the enhanced adrenergic reactivity of goat cerebral arteries after ischemia-reperfusion. Brain Res. 2010, 1346, 121–131. [Google Scholar] [CrossRef]
- Hirokawa, K.; O’Shaughnessy, K.M.; Ramrakha, P.; Wilkins, M.R. Inhibition of nitric oxide synthesis in vascular smooth muscle by retinoids. Br. J. Pharmacol. 1994, 113, 1448–1454. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, Y.; Zhang, D.; Wang, W.; Xie, H.; Ruan, J.; Jin, Y.; Li, T.; Li, X.; Zhao, B.; et al. Ca2+ oscillation in vascular smooth muscle cells control myogenic spontaneous vasomotion and counteract post-ischemic no-reflow. Commun. Biol. 2024, 7, 332. [Google Scholar] [CrossRef] [PubMed]
- Noman, M.A.A.; Kyzer, J.L.; Chung, S.S.W.; Wolgemuth, D.J.; Georg, G.I. Retinoic acid receptor antagonists for male contraception: Current status†. Biol. Reprod. 2020, 103, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Branka, J.E.; Vallette, G.; Jarry, A.; Laboisse, C.L. Stimulation of mucin exocytosis from human epithelial cells by nitric oxide: Evidence for a cGMP-dependent and a cGMP-independent pathway. Biochem. J. 1997, 323 Pt 2, 521–524. [Google Scholar] [CrossRef]
- da Silva Filho, P.M.; Paz, I.A.; Nascimento, N.; Abreu, D.S.; Lopes, L.G.F.; Sousa, E.H.S.; Longhinotti, E. Nitroprusside—Expanding the Potential Use of an Old Drug Using Nanoparticles. Mol. Pharm. 2023, 20, 6–22. [Google Scholar] [CrossRef]
- Mawson, A.R. Retinoids in the treatment of glioma: A new perspective. Cancer Manag. Res. 2012, 4, 233–241. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | RA Incubation | Medium |
|---|---|---|
| HBSS + Glucose | 0.1% ethanol (solvent control) | No-OGD-treated RMC |
| 0.1 µM RA | No-OGD-treated RMC | |
| 1 µM RA | No-OGD-treated RMC | |
| 5 µM RA | No-OGD-treated RMC | |
| 10 µM RA | No-OGD-treated RMC | |
| OGD (4 h) | 0.1% ethanol (solvent control) | OGD-treated RMC |
| 0.1 µM RA | OGD-treated RMC | |
| 1 µM RA | OGD-treated RMC | |
| 5 µM RA | OGD-treated RMC | |
| 10 µM RA | OGD-treated RMC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pouso, M.R.; Farinha, E.; Costa, H.E.; Lorigo, M.; Baltazar, G.; Cairrao, E. Effect of Retinoic Acid on the Cerebral Vasculature: Analysis of the Vasoactive Response of Smooth Muscle Cells in Normal and Ischemic Contexts. J. Xenobiot. 2025, 15, 69. https://doi.org/10.3390/jox15030069
Pouso MR, Farinha E, Costa HE, Lorigo M, Baltazar G, Cairrao E. Effect of Retinoic Acid on the Cerebral Vasculature: Analysis of the Vasoactive Response of Smooth Muscle Cells in Normal and Ischemic Contexts. Journal of Xenobiotics. 2025; 15(3):69. https://doi.org/10.3390/jox15030069
Chicago/Turabian StylePouso, Manuel R., Emanuel Farinha, Henrique E. Costa, Margarida Lorigo, Graça Baltazar, and Elisa Cairrao. 2025. "Effect of Retinoic Acid on the Cerebral Vasculature: Analysis of the Vasoactive Response of Smooth Muscle Cells in Normal and Ischemic Contexts" Journal of Xenobiotics 15, no. 3: 69. https://doi.org/10.3390/jox15030069
APA StylePouso, M. R., Farinha, E., Costa, H. E., Lorigo, M., Baltazar, G., & Cairrao, E. (2025). Effect of Retinoic Acid on the Cerebral Vasculature: Analysis of the Vasoactive Response of Smooth Muscle Cells in Normal and Ischemic Contexts. Journal of Xenobiotics, 15(3), 69. https://doi.org/10.3390/jox15030069