
The Neural Basis of Skull Vibration Induced Nystagmus (SVIN)


Abstract
:1. Introduction
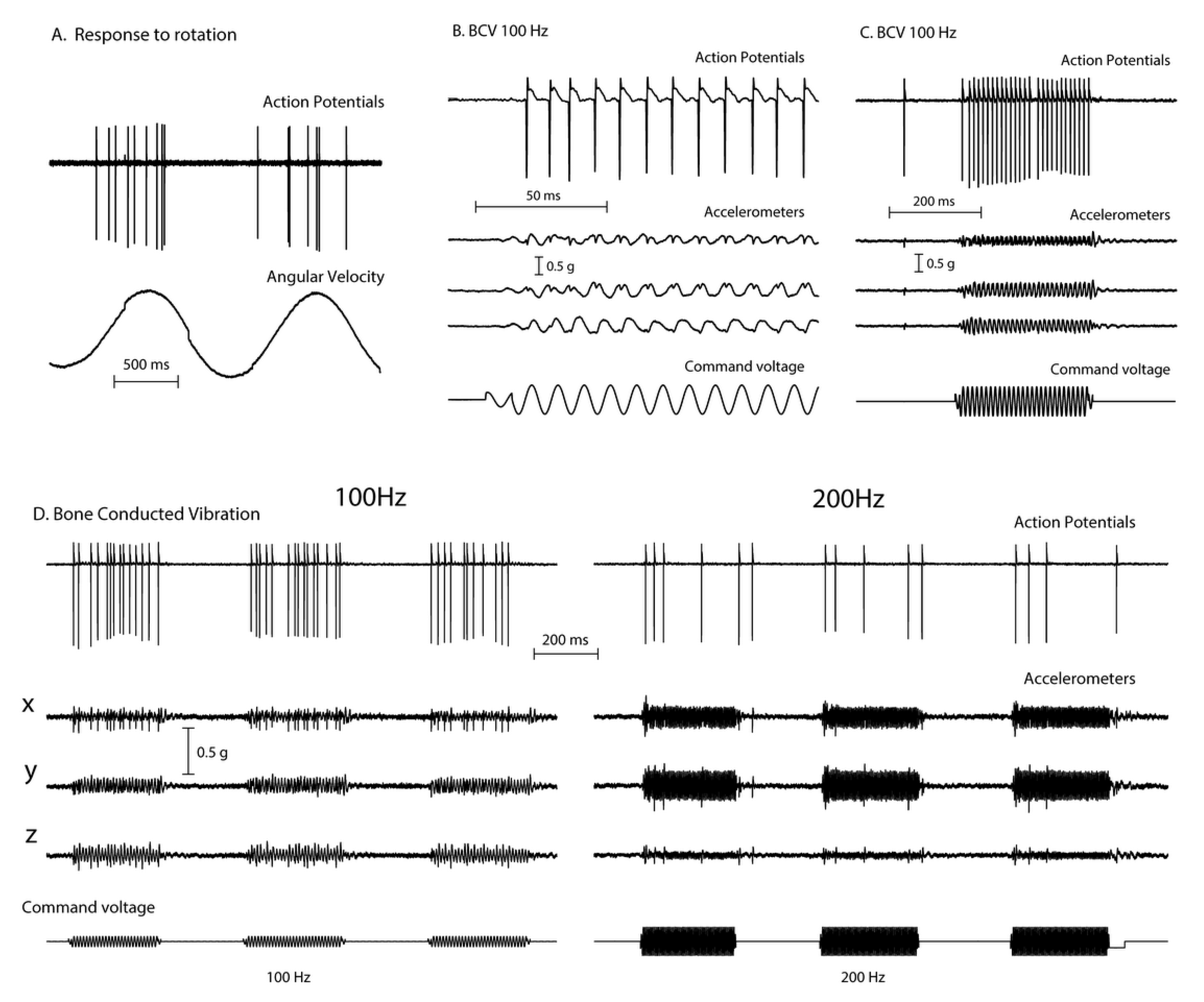
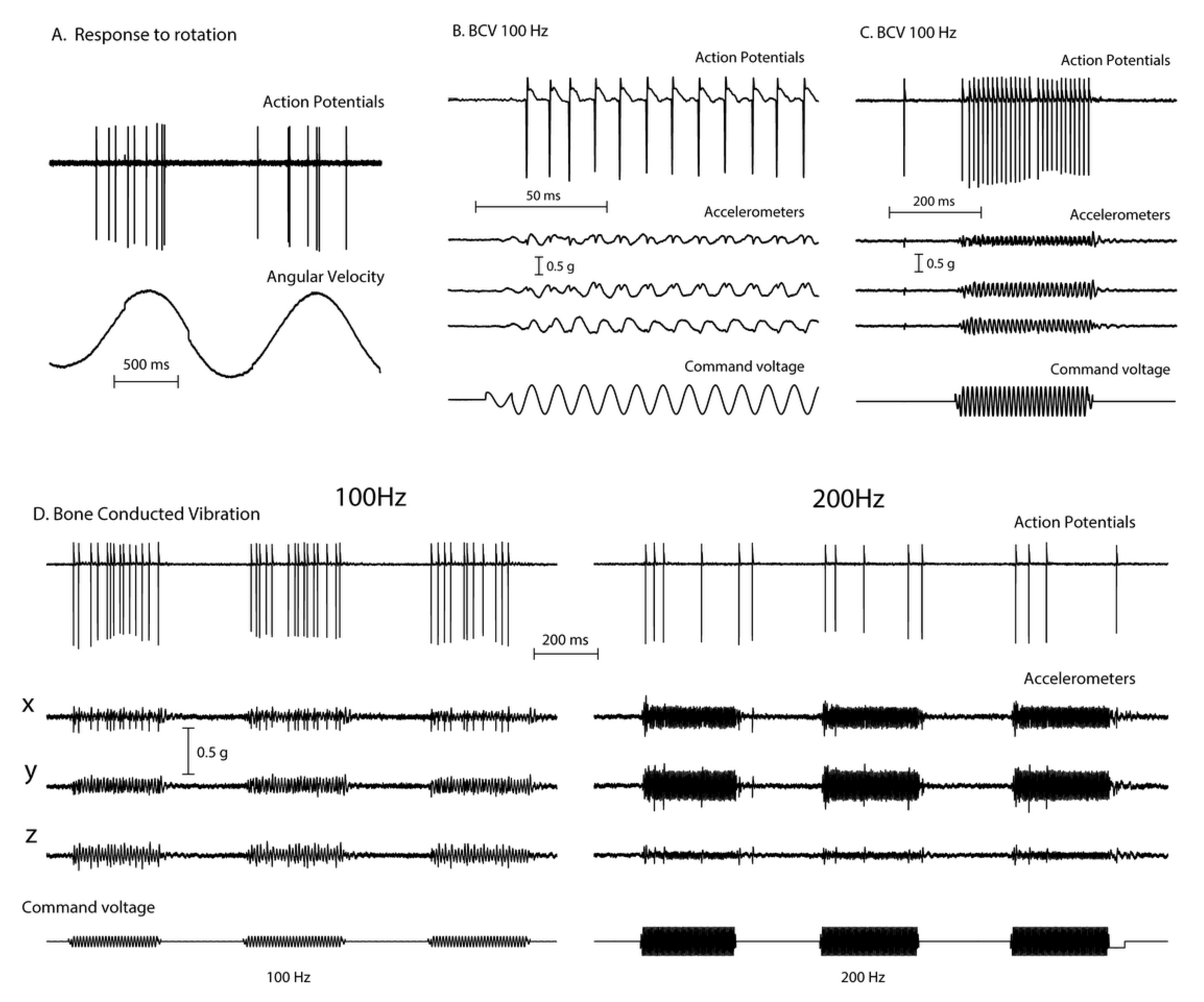
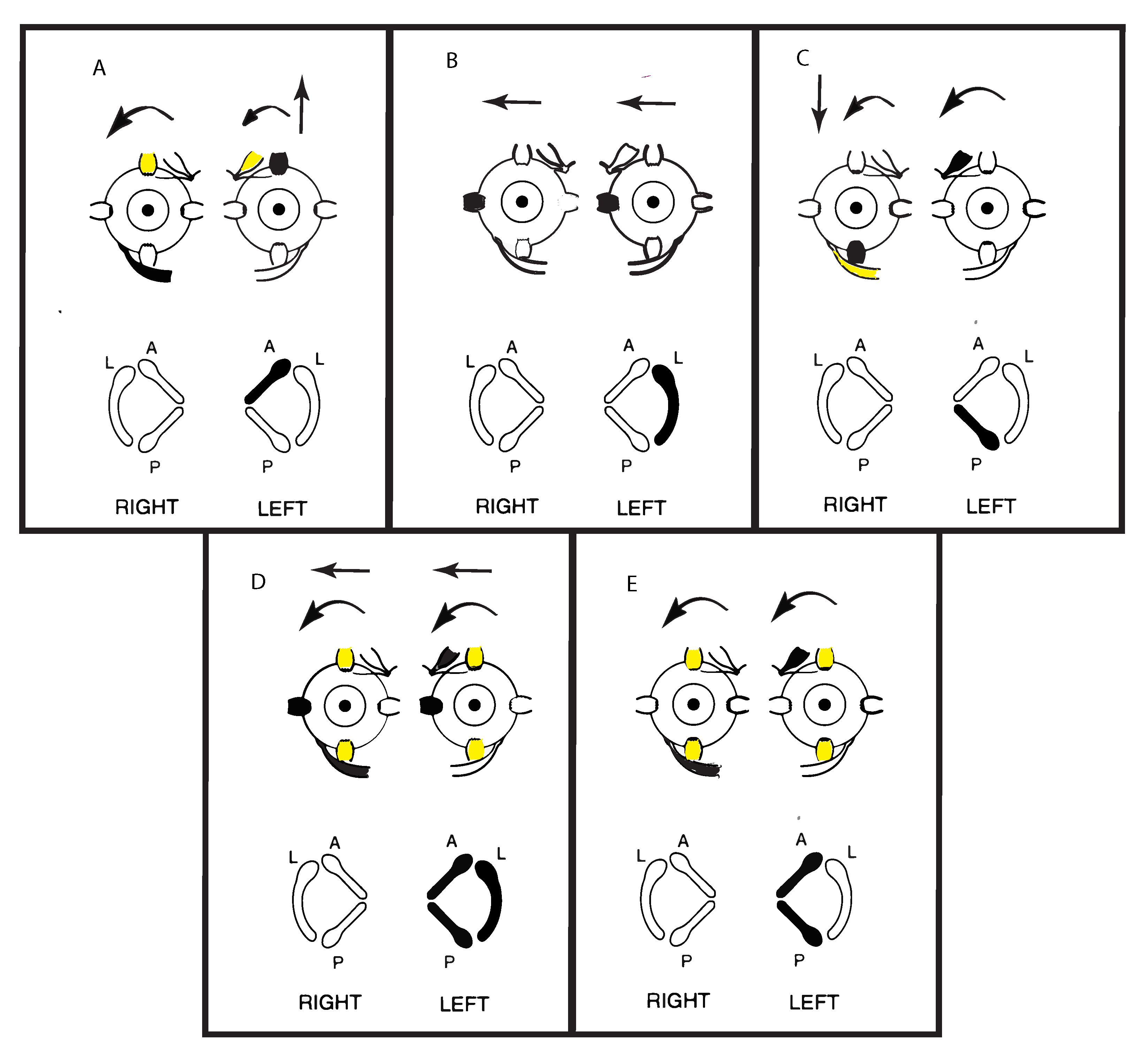
2. Vibration—Physiological Evidence
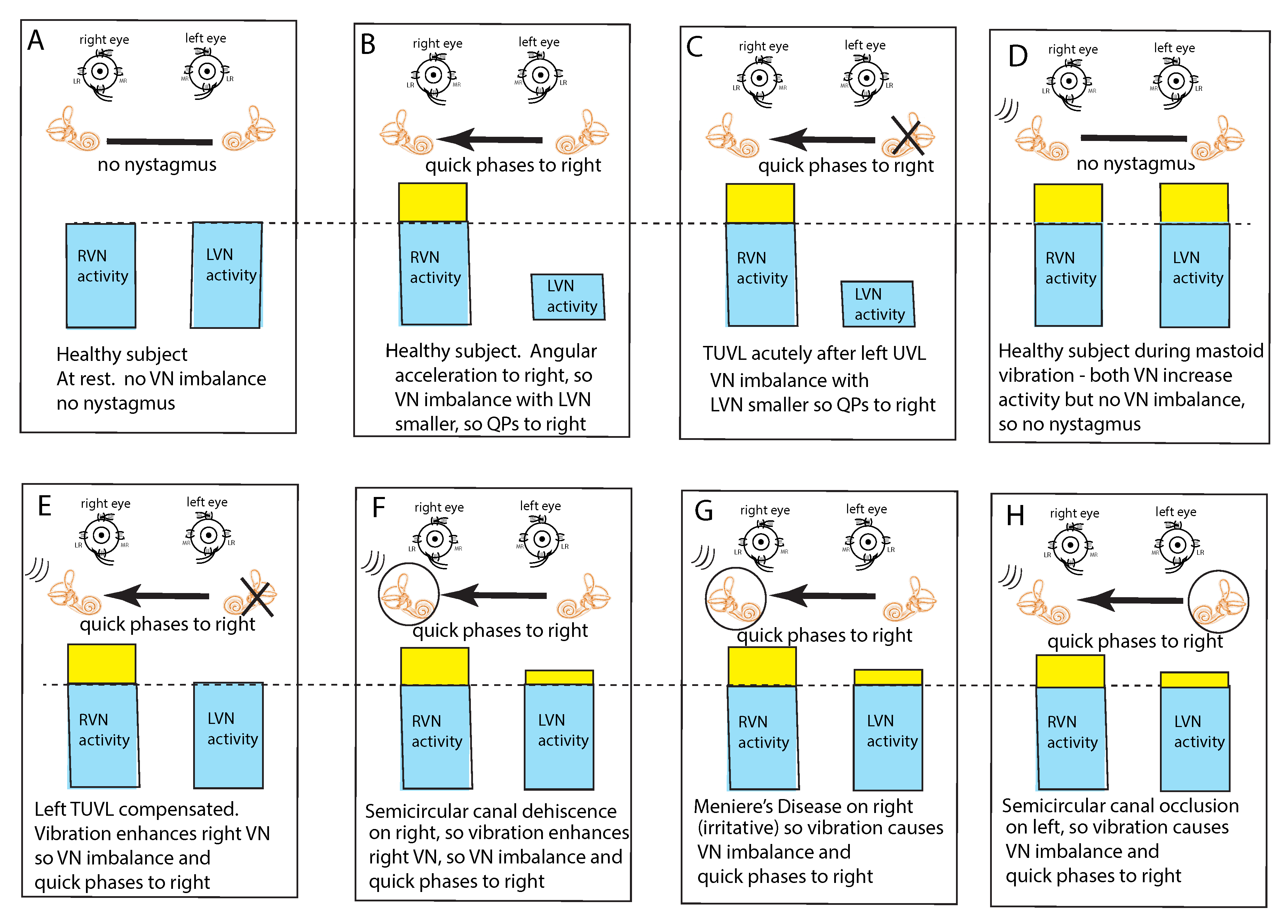
3. A Simplified Schematic Model of Nystagmus Generation
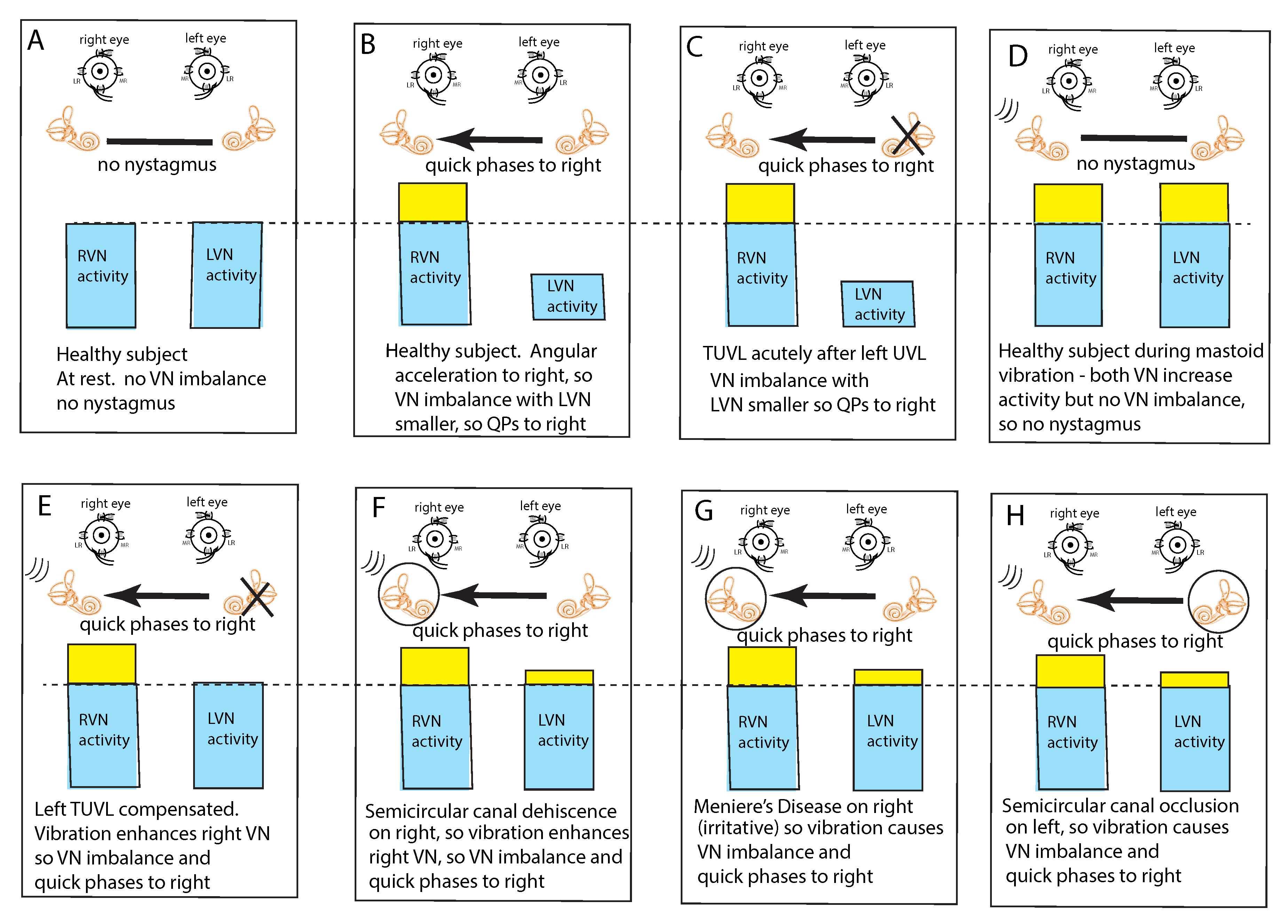
Nystagmus Is Due to an Asymmetry of Average Neural Activity between the Two Vestibular Nuclei (VN)
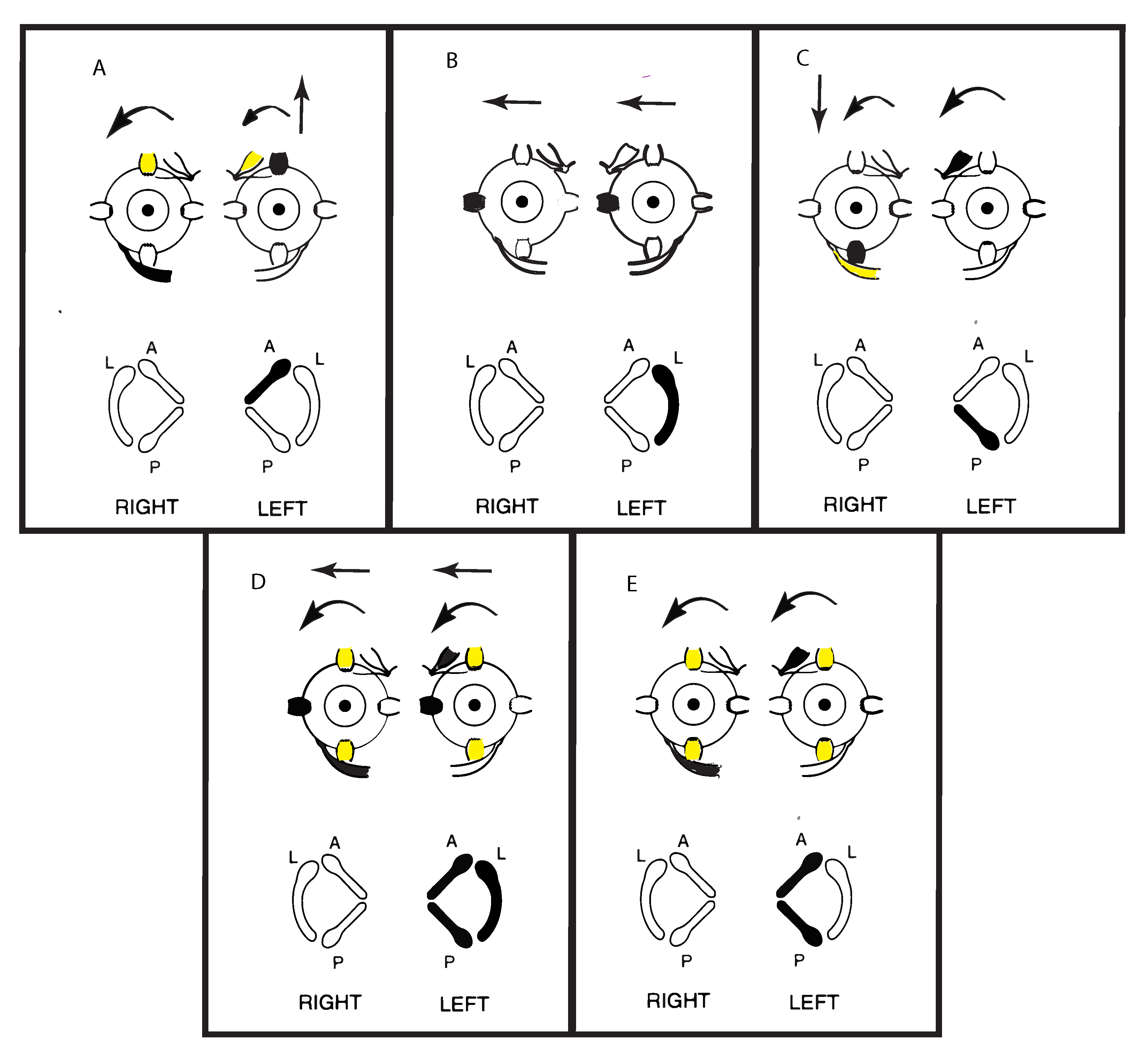
4. Clinical Value and Interpretation of SVIN
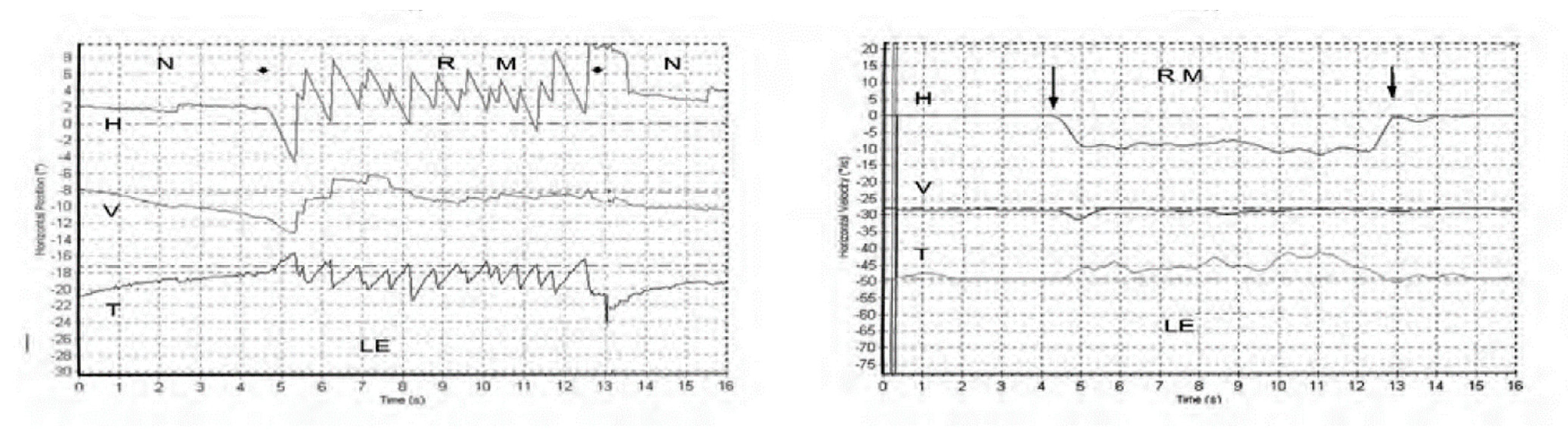
5. SVIN—The Slow Phase Eye Velocity
6. Discussion
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MD | Menière’s Disease |
| QP | quick phases |
| SCD | semicircular canal dehiscence |
| SPV | slow phase eye velocity |
| SVIN | skull vibration induced nystagmus |
| TUVL | total unilateral vestibular loss |
| VN | vestibular nuclei |
References
- Dumas, G.; Curthoys, I.S.; Lion, A.; Perrin, P.; Schmerber, S. The skull vibration-induced nystagmus test of vestibular function-a review. Front. Neurol. 2017, 8, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curthoys, I.S. Generation of the quick phase of horizontal vestibular nystagmus. Exp. Brain Res. 2002, 143, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Curthoys, I.S. The new vestibular stimuli: Sound and vibration-anatomical, physiological and clinical evidence. Exp. Brain Res. 2017, 235, 957–972. [Google Scholar] [CrossRef] [PubMed]
- Dlugaiczyk, J.; Burgess, A.M.; Curthoys, I.S. Activation of guinea pig irregular semicircular canal afferents by 100 hz vibration: Clinical implications for vibration-induced nystagmus and vestibular-evoked myogenic potentials. Otol. Neurotol. 2020, 41, e961–e970. [Google Scholar] [CrossRef] [PubMed]
- Cohen, B.; Suzuki, J.; Bender, M.B. Eye movements from semicircular canal nerve stimulation in cat. Ann. Otol. Rhinol. Laryngol. 1964, 73, 153–169. [Google Scholar] [CrossRef] [PubMed]
- Dumas, G.; Fabre, C.; Charpiot, A.; Fath, L.; Chaney-Vuong, H.; Perrin, P.; Schmerber, S. Skull vibration-induced nystagmus test in a human model of horizontal canal plugging. Audiol. Res. 2021, 11, 301–312. [Google Scholar] [CrossRef]
- Dumas, G.; Lion, A.; Karkas, A.; Perrin, P.; Perottino, F.; Schmerber, S. Skull vibration-induced nystagmus test in unilateral superior canal dehiscence and otosclerosis: A vestibular Weber test. Acta Otolaryngol. Stockh. 2014, 134, 588–600. [Google Scholar] [CrossRef]
- Dumas, G.; Tan, H.; Dumas, L.; Perrin, P.; Lion, A.; Schmerber, S. Skull vibration induced nystagmus in patients with superior semicircular canal dehiscence. Eur. Ann. Otorhinolaryngol. Head Neck Dis. 2019, 136, 263–272. [Google Scholar] [CrossRef]
- Fernandez, C.; Goldberg, J.M.; Baird, R.A. The vestibular nerve of the chinchilla. III. Peripheral innervation patterns in the utricular macula. J. Neurophysiol. 1990, 63, 767–780. [Google Scholar] [CrossRef] [PubMed]
- Curthoys, I.S.; Vulovic, V.; Sokolic, L.; Pogson, J.; Burgess, A.M. Irregular primary otolith afferents from the guinea pig utricular and saccular maculae respond to both bone conducted vibration and to air conducted sound. Brain Res. Bull. 2012, 89, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Curthoys, I.S.; Vulovic, V.; Burgess, A.M.; Sokolic, L.; Goonetilleke, S.C. The response of guinea pig primary utricular and saccular irregular neurons to bone-conducted vibration (BCV) and air-conducted, sound (ACS). Hear. Res. 2016, 331, 131–143. [Google Scholar] [CrossRef]
- Goldberg, J.M.; Fernandez, C. Conduction times and background discharge of vestibular afferents. Brain Res. 1977, 122, 545–550. [Google Scholar] [CrossRef]
- Yagi, T.; Markham, C.H.; Simpson, N.E. Relationship of conduction-velocity to other physiological properties of cats horizontal canal neurons. Exp. Brain Res. 1977, 30, 587–600. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.F.; Curthoys, I.S. Neuronal activity in the ipsilateral medial vestibular nucleus of the guinea pig following unilateral labyrinthectomy. Brain Res. 1988, 444, 308–319. [Google Scholar] [CrossRef]
- Smith, P.F.; Curthoys, I.S. Neuronal activity in the contralateral medial vestibular nucleus of the guinea pig following unilateral labyrinthectomy. Brain Res. 1988, 444, 295–307. [Google Scholar] [CrossRef]
- Curthoys, I.S.; Smith, P.F.; Darlington, C.L. Postural compensation in the guinea pig following unilateral labyrinthectomy. Prog. Brain Res. 1988, 76, 375–384. [Google Scholar]
- Stenfelt, S.; Goode, R.L. Bone-conducted sound: Physiological and clinical aspects. Otol. Neurotol. 2005, 26, 1245–1261. [Google Scholar] [CrossRef]
- Rosowski, J.J.; Songer, J.E.; Nakajima, H.H.; Brinsko, K.M.; Merchant, S.N. Clinical, experimental, and theoretical investigations of the effect of superior semicircular canal dehiscence on hearing mechanisms. Otol. Neurotol. 2004, 25, 323–332. [Google Scholar] [CrossRef]
- Dlugaiczyk, J.; Burgess, A.M.; Goonetilleke, S.C.; Sokolic, L.; Curthoys, I.S. Superior Canal Dehiscence syndrome: Relating clinical findings with vestibular neural responses from a guinea pig model. Otol. Neurotol. 2019, 40, e406–e414. [Google Scholar] [CrossRef]
- Manzari, L.; Burgess, A.M.; MacDougall, H.G.; Bradshaw, A.P.; Curthoys, I.S. Rapid fluctuations in dynamic semicircular canal function in early Meniere’s disease. Eur. Arch. Otorhinolaryngol. 2011, 268, 637–639. [Google Scholar] [CrossRef]
- Bance, M.; Mai, M.; Tomlinson, D.; Rutka, J. The changing direction of nystagmus in acute menieres-disease—Pathophysiological implications. Laryngoscope 1991, 101, 197–201. [Google Scholar] [CrossRef]
- McClure, J.A.; Copp, J.C.; Lycett, P. Recovery nystagmus in menieres-disease. Laryngoscope 1981, 91, 1727–1737. [Google Scholar] [CrossRef]
- Sadeghi, S.G.; Goldberg, J.M.; Minor, L.B.; Cullen, K.E. Effects of canal plugging on the vestibuloocular reflex and vestibular nerve discharge during passive and active head rotations. J. Neurophysiol. 2009, 102, 2693–2703. [Google Scholar] [CrossRef] [Green Version]
- Cremer, P.D.; Halmagyi, G.M.; Aw, S.T.; Curthoys, I.S.; McGarvie, L.A.; Todd, M.J.; Black, R.A.; Hannigan, I.P. Semicircular canal plane head impulses detect absent function of individual semicircular canals. Brain 1998, 121 Pt 4, 699–716. [Google Scholar] [CrossRef] [Green Version]
- Aw, S.T.; Halmagyi, G.M.; Haslwanter, T.; Curthoys, I.S.; Yavor, R.A.; Todd, M.J. Three-dimensional vector analysis of the human vestibuloocular reflex in response to high-acceleration head rotations.2. Responses in subjects with unilateral vestibular loss and selective semicircular canal occlusion. J. Neurophysiol. 1996, 76, 4021–4030. [Google Scholar] [CrossRef]
- Curthoys, I.S.; Grant, J.W.; Pastras, C.J.; Frohlich, L.; Brown, D.J. Similarities and differences between vestibular and cochlear systems—A review of clinical and physiological evidence. Front. Neurosci. 2021. [Google Scholar] [CrossRef]
- Karlberg, M.; Aw, S.T.; Black, R.A.; Todd, M.J.; MacDougall, H.G.; Halmagyi, G.M. Vibration-induced ocular torsion and nystagmus after unilateral vestibular deafferentation. Brain 2003, 126, 956–964. [Google Scholar] [CrossRef] [PubMed]
- Rubin, A.M.; Liedgren, S.R.C.; Milne, A.C.; Young, J.A.; Fredrickson, J.M. Vestibular and somatosensory interaction in cat vestibular nuclei. Pflug. Arch. Eur. J. Physiol. 1977, 371, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Ohkawa, T.; Uchino, Y.; Wilson, V.J. Excitatory connections between neurons of the central cervical nucleus and vestibular neurons in the cat. Exp. Brain Res. 1997, 115, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Wilson, V.J.; Wylie, R.M.; Marco, L.A. Synaptic inputs to cells in medial vestibular nucleus. J. Neurophysiol. 1968, 31, 176. [Google Scholar] [CrossRef] [PubMed]
- McConville, K.M.V.; Tomlinson, R.D.; Na, E.Q. Behavior of eye-movement-related cells in the vestibular nuclei during combined rotational and translational stimuli. J. Neurophysiol. 1996, 76, 3136–3148. [Google Scholar] [CrossRef] [PubMed]
- Curthoys, I.S.; Markham, C.H. Convergence of labyrinthine influences on units in the vestibular nuclei of the cat. I. Natural stimulation. Brain Res. 1971, 35, 469–490. [Google Scholar] [CrossRef]
- Curthoys, I.S.; Wearne, S.L.; Dai, M.; Halmagyi, G.M.; Holden, J.R. Linear acceleration modulates the nystagmus induced by angular acceleration stimulation of the horizontal canal. Ann. N. Y. Acad. Sci. 1992, 656, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Soper, J.; Lohse, C.M.; Eggers, S.D.Z.; Kaufman, K.R.; McCaslin, D.L. Agreement between the Skull Vibration-Induced Nystagmus Test and Semicircular Canal and Otolith Asymmetry. J. Am. Acad. Audiol. 2021, 32, 283–289. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Main Clinical Results from SVIN Studies of the Response of Patients with Total Unilateral Vestibular Loss (TUVL) |
|---|
|
|
|
|
|
|
| Other major results |
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Curthoys, I.S. The Neural Basis of Skull Vibration Induced Nystagmus (SVIN). Audiol. Res. 2021, 11, 557-566. https://doi.org/10.3390/audiolres11040050
Curthoys IS. The Neural Basis of Skull Vibration Induced Nystagmus (SVIN). Audiology Research. 2021; 11(4):557-566. https://doi.org/10.3390/audiolres11040050
Chicago/Turabian StyleCurthoys, Ian S. 2021. "The Neural Basis of Skull Vibration Induced Nystagmus (SVIN)" Audiology Research 11, no. 4: 557-566. https://doi.org/10.3390/audiolres11040050
APA StyleCurthoys, I. S. (2021). The Neural Basis of Skull Vibration Induced Nystagmus (SVIN). Audiology Research, 11(4), 557-566. https://doi.org/10.3390/audiolres11040050