
Blood Coagulation Favors Anti-Inflammatory Immune Responses in Whole Blood

 , , ,
, , , 
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Collection of Whole Blood Samples
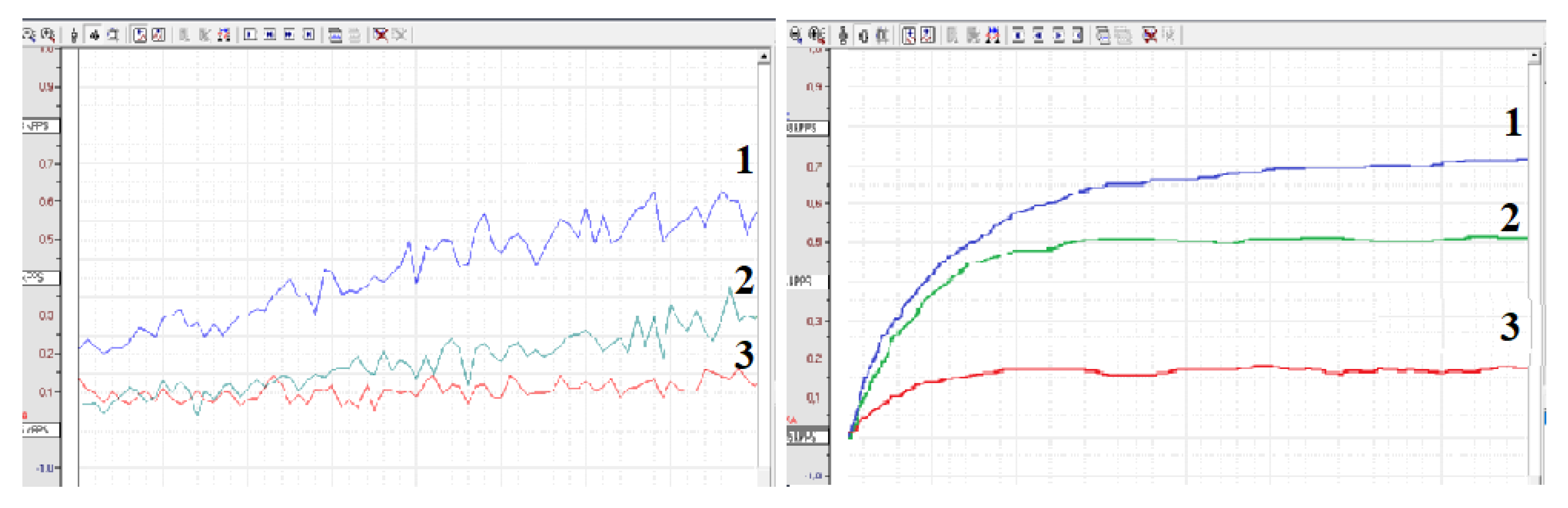
2.2. Measurements of Reactive Oxygen Species (ROS)
2.3. Assaying Bioactive Molecules
2.4. Statistical Analysis
3. Results
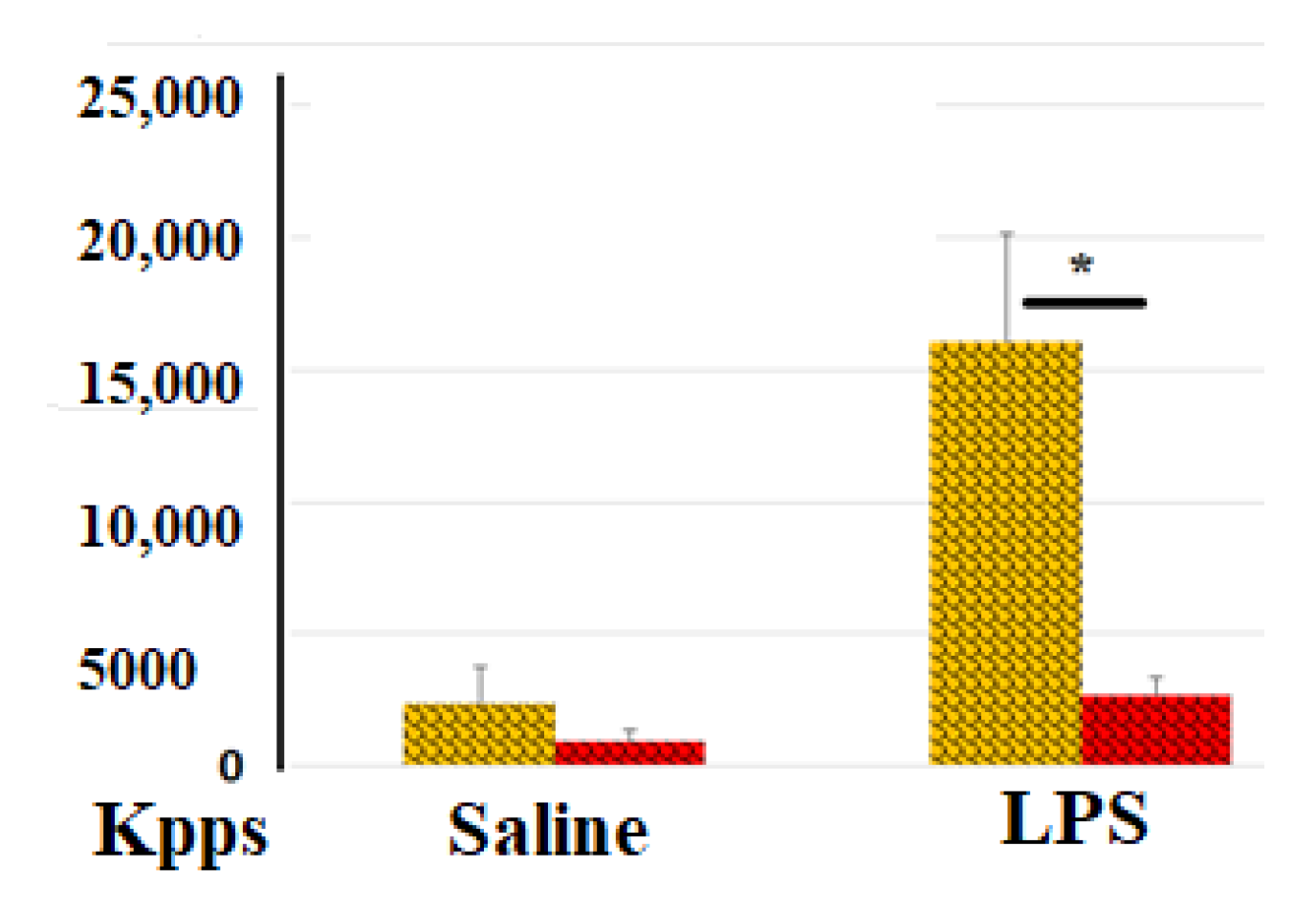
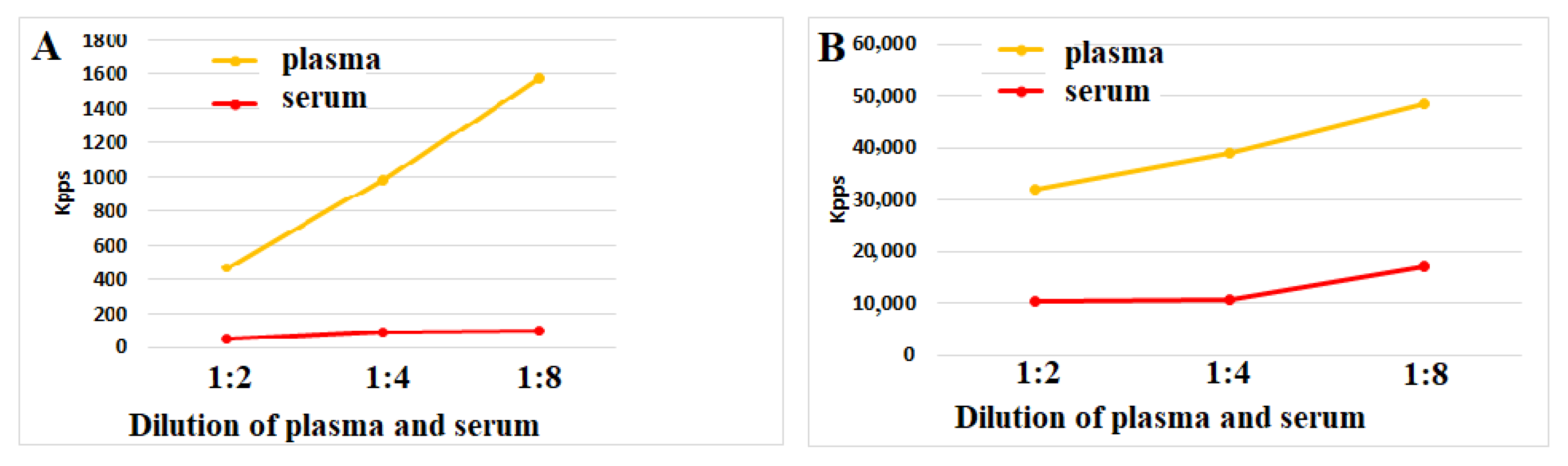
Antioxidant Effect of Blood Clotting
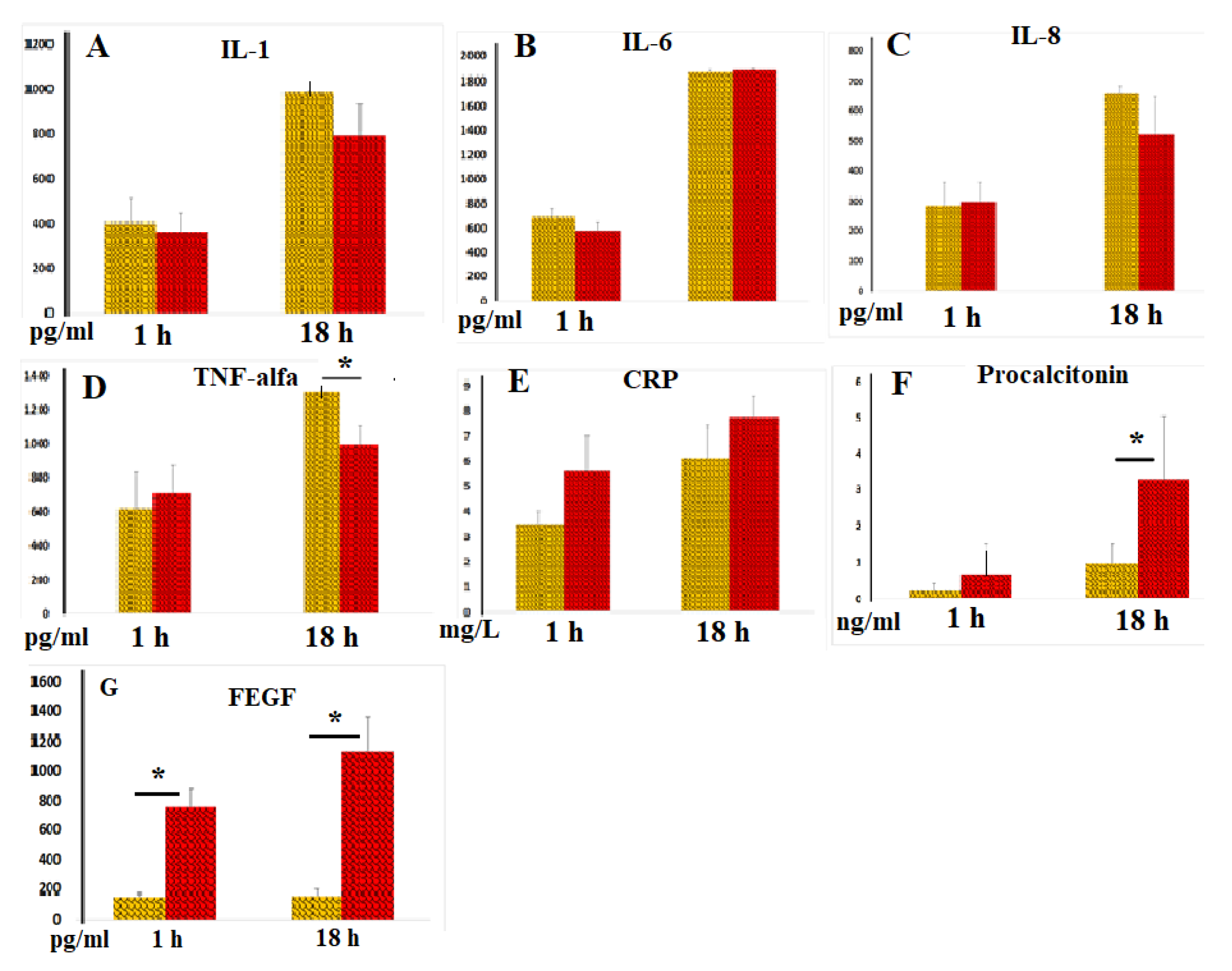
Anti-Inflammatory Effects of Blood Clotting
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Łukasik, Z.M.; Makowski, M.; Makowska, J.S. From blood coagulation to innate and adaptive immunity: The role of platelets in the physiology and pathology of autoimmune disorders. Rheumatol. Int. 2018, 38, 959–974. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Antoniak, S. The coagulation system in host defense. Res. Pract. Thromb. Haemost. 2018, 2, 549–557. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Xiong, Z.; Wang, Q.; Li, W.; Huang, L.; Zhang, J.; Zhu, J.; Xie, B.; Wang, S.; Kuang, H.; Lin, X.; et al. Corrigendum: Platelet-derived growth factor-D activates complement system to propagate macrophage polarization and neovascularization. Front. Cell Dev. Biol. 2022, 10, 848292, Erratum in Front. Cell Dev. Biol. 2021, 9, 686886. 10.3389/fcell.2021.686886. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Vitali, M.; Ometti, M.; Drossinos, A.; Pironti, P.; Santoleri, L.; Salini, V. Autologous conditioned serum: Clinical and functional results using a novel disease modifying agent for the management of knee osteoarthritis. J. Drug Assess. 2020, 9, 43–51. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Suárez-Álvarez, B.; Liapis, H.; Anders, H.J. Links between coagulation, inflammation, regeneration, and fibrosis in kidney pathology. Lab. Invest. 2016, 96, 378–390. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, N.; Hilger, A.; Zarbock, A.; Rossaint, J. Platelets at the crossroads of pro-inflammatory and resolution pathways during inflammation. Cells 2022, 11, 1957. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Seledtsov, V.I.; von Delwig, A.A. Oxygen therapy in traditional and immunotherapeutic treatment protocols of cancer patients: Current reality and future prospects. Expert. Rev. Anticancer Ther. 2022, 22, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Bester, J.; Matshailwe, C.; Pretorius, E. Simultaneous presence of hypercoagulation and increased clot lysis time due to IL-1β, IL-6 and IL-8. Cytokine 2018, 110, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.H.; Conway, E.M. Cross talk pathways between coagulation and inflammation. Circ. Res. 2016, 118, 1392–1408. [Google Scholar] [CrossRef] [PubMed]
- Seledtsov, V.I.; von Delwig, A.A. Therapeutic stimulation of glycolytic ATP production for treating ROS-mediated cellular senescence. Metabolites 2022, 12, 1160. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Gros, A.; Ollivier, V.; Ho-Tin-Noé, B. Platelets in inflammation: Regulation of leukocyte activities and vascular repair. Front. Immunol. 2015, 5, 678. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Li, N.; Ji, Q.; Hjemdahl, P. Platelet-lymphocyte conjugation differs between lymphocyte subpopulations. J. Thromb. Haemost. 2006, 4, 874–881. [Google Scholar] [CrossRef] [PubMed]
- Zamora, C.; Cantó, E.; Nieto, J.C.; Ortiz, M.A.; Diaz-Torné, C.; Diaz-Lopez, C.; Llobet, J.M.; Juarez, C.; Vidal, S. Functional consequences of platelet binding to T lymphocytes in inflammation. J. Leukoc. Biol. 2013, 94, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Gerdes, N.; Zhu, L.; Ersoy, M.; Hermansson, A.; Hjemdahl, P.; Hu, H.; Hansson, G.K.; Li, N. Platelets regulate CD4⁺ T-cell differentiation via multiple chemokines in humans. Thromb. Haemost. 2011, 106, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Huang, Z.; Stålesen, R.; Hansson, G.K.; Li, N. Platelets provoke distinct dynamics of immune responses by differentially regulating CD4+ T-cell proliferation. J. Thromb. Haemost. 2014, 12, 1156–1165. [Google Scholar] [CrossRef] [PubMed]
- Li, N. Platelet-lymphocyte cross-talk. J. Leukoc. Biol. 2008, 83, 1069–1078. [Google Scholar] [CrossRef] [PubMed]
- Ciesielska, A.; Matyjek, M.; Kwiatkowska, K. TLR4 and CD14 trafficking and its influence on LPS-induced pro-inflammatory signaling. Cell. Mol. Life Sci. 2021, 78, 1233–1261. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Armstrong, M.T.; Rickles, F.R.; Armstrong, P.B. Capture of lipopolysaccharide (endotoxin) by the blood clot: A comparative study. PLoS ONE 2013, 8, e80192. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wilhelm, G.; Mertowska, P.; Mertowski, S.; Przysucha, A.; Strużyna, J.; Grywalska, E.; Torres, K. The Crossroads of the Coagulation System and the Immune System: Interactions and Connections. Int. J. Mol. Sci. 2023, 24, 12563. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seledtsov, V.I.; Pyshenko, A.A.; Lyubavskaya, T.Y.; Seledtsova, I.A.; von Delwig, A.A. Blood Coagulation Favors Anti-Inflammatory Immune Responses in Whole Blood. Hematol. Rep. 2025, 17, 19. https://doi.org/10.3390/hematolrep17020019
Seledtsov VI, Pyshenko AA, Lyubavskaya TY, Seledtsova IA, von Delwig AA. Blood Coagulation Favors Anti-Inflammatory Immune Responses in Whole Blood. Hematology Reports. 2025; 17(2):19. https://doi.org/10.3390/hematolrep17020019
Chicago/Turabian StyleSeledtsov, Victor I., Anatoly A. Pyshenko, Tatyana Ya. Lyubavskaya, Irina A. Seledtsova, and Alexei A. von Delwig. 2025. "Blood Coagulation Favors Anti-Inflammatory Immune Responses in Whole Blood" Hematology Reports 17, no. 2: 19. https://doi.org/10.3390/hematolrep17020019
APA StyleSeledtsov, V. I., Pyshenko, A. A., Lyubavskaya, T. Y., Seledtsova, I. A., & von Delwig, A. A. (2025). Blood Coagulation Favors Anti-Inflammatory Immune Responses in Whole Blood. Hematology Reports, 17(2), 19. https://doi.org/10.3390/hematolrep17020019