
The Importance of the Glomus Genus as a Potential Candidate for Sustainable Agriculture Under Arid Environments: A Review

 , , ,
, , ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
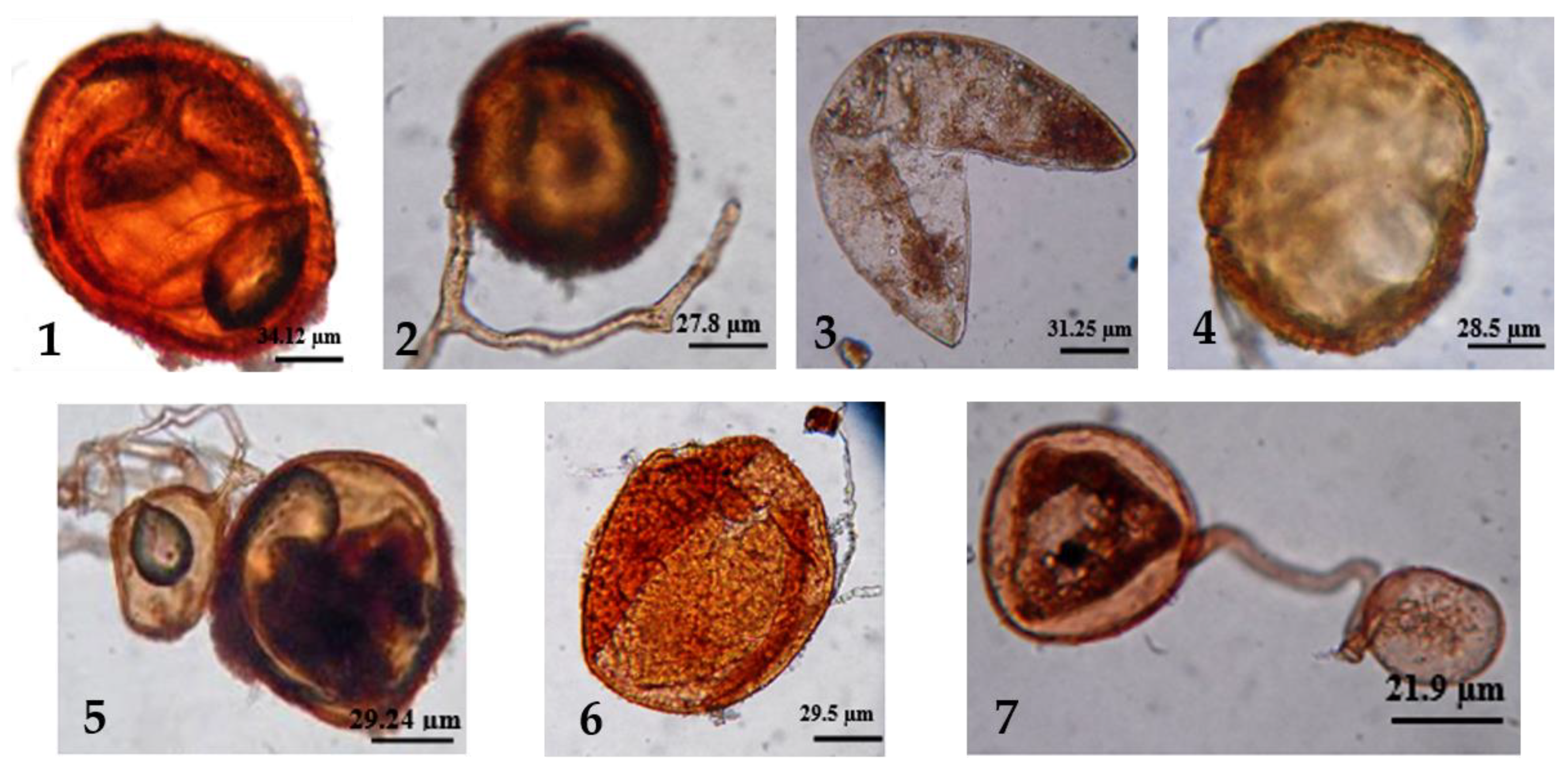
2. Glomus Genus
2.1. Phylogenetic Relationships
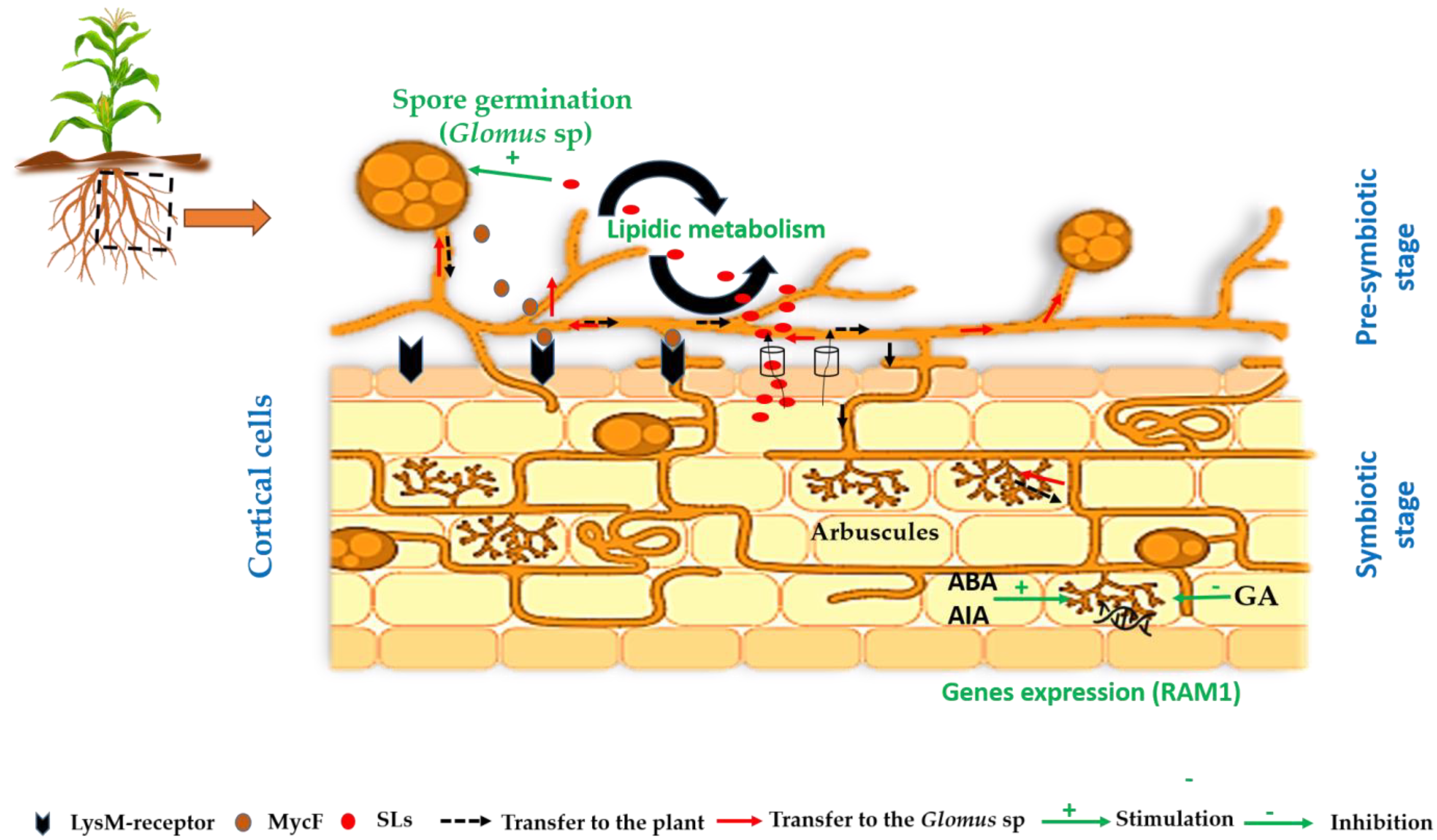
2.2. Life Cycle
2.3. Molecular Signaling Pathways in Plant–Glomus Symbiosis
3. Effect of Salt and Drought Stress on Crop Production
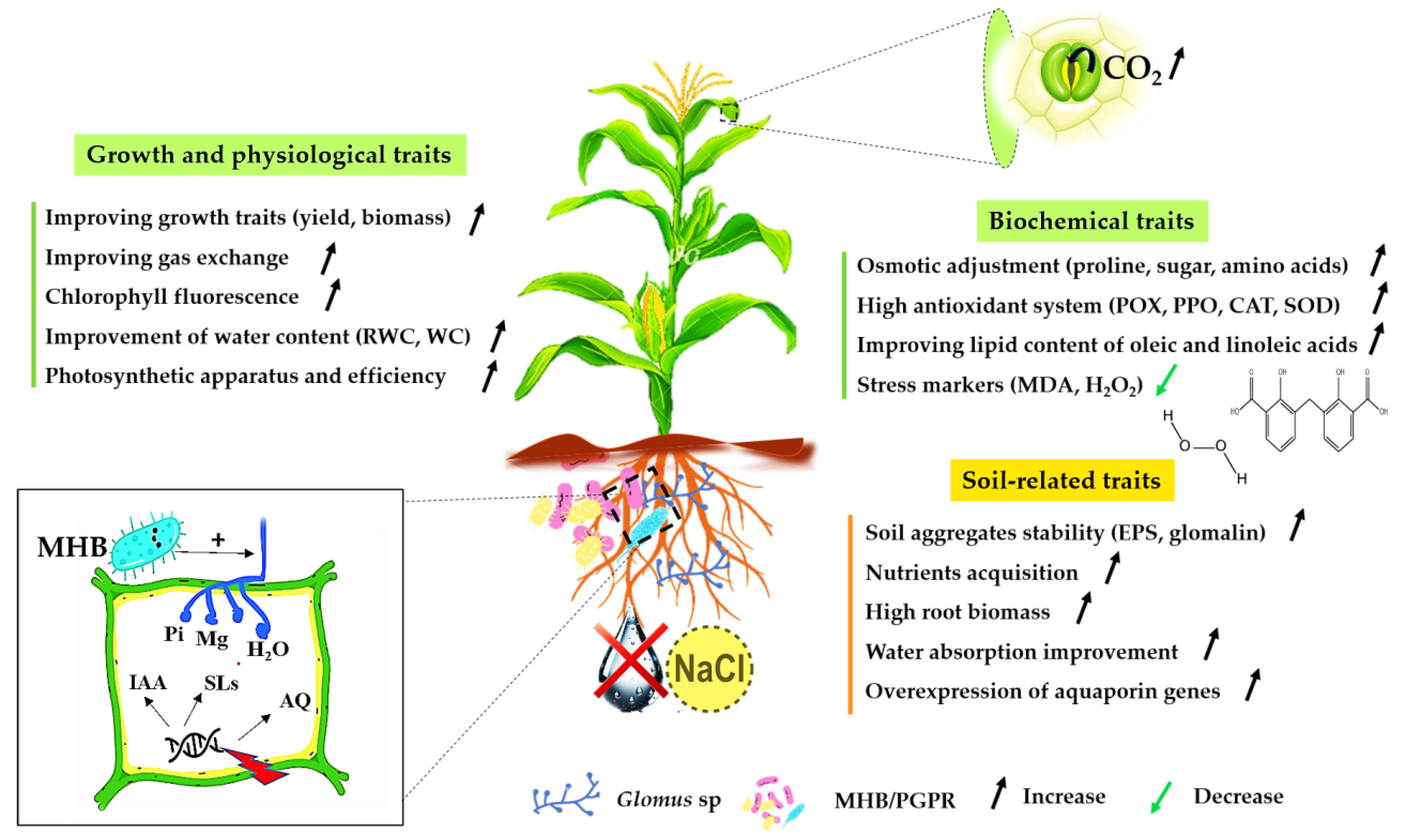
4. Interaction of Glomus and Abiotic Stress
4.1. Drought
4.1.1. Glomus and Drought Stress
4.1.2. Mechanisms of Tolerance to Drought Stress
4.2. Salinity
4.2.1. Glomus and Salt Stress
4.2.2. Salt Stress Tolerance Mechanisms
5. Interaction of Glomus and PGPR Under Abiotic Stresses
6. Factors Affecting Glomus Responsiveness to Mycorrhizal Symbiosis
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Saleem, A.; Anwar, S.; Nawaz, T.; Fahad, S.; Saud, S.; Ur Rahman, T.; Khan, M.N.R.; Nawaz, T. Securing a Sustainable Future: The Climate Change Threat to Agriculture, Food Security, and Sustainable Development Goals. J. Umm Al-Qura Univ. Appl. Sci. 2024, 1–17. [Google Scholar] [CrossRef]
- FAOSTAT. FAO Statistical Yearbook 2020—World Food and Agriculture. In Food and Agriculture Organization of the United Nations; FAOSTAT: Rome, Italy, 2022; ISBN 9789251333945. [Google Scholar]
- Biswas, A.; Sarkar, S.; Das, S.; Dutta, S.; Roy Choudhury, M.; Giri, A.; Bera, B.; Bag, K.; Mukherjee, B.; Banerjee, K.; et al. Water Scarcity: A Global Hindrance to Sustainable Development and Agricultural Production—A Critical Review of the Impacts and Adaptation Strategies. Camb. Prism. Water 2025, 3, e4. [Google Scholar] [CrossRef]
- Haseeb, M.; Zandi, G.; Hartani, N.H.; Pahi, M.H.; Nadeem, S.; Kedah, S. Environmental Analysis of the Effect of Population Growth Rate on Supply Chain Performance and Economic Growth of Indonesia. Ekoloji Derg. 2019, 28, 417–426. [Google Scholar]
- Zhang, X.; Han, L.; Wei, H.; Tan, X.; Zhou, W.; Li, W.; Qian, Y. Linking Urbanization and Air Quality Together: A Review and a Perspective on the Future Sustainable Urban Development. J. Clean. Prod. 2022, 346, 130988. [Google Scholar] [CrossRef]
- Ekka, P.; Patra, S.; Upreti, M.; Kumar, G.; Kumar, A.; Saikia, P. Land Degradation and Its Impacts on Biodiversity and Ecosystem Services. In Land and Environmental Management Through Forestry; Wiley Online Library: Hoboken, NJ, USA, 2023; pp. 77–101. [Google Scholar] [CrossRef]
- Dincă, L.C.; Grenni, P.; Onet, C.; Onet, A. Fertilization and Soil Microbial Community: A Review. Appl. Sci. 2022, 12, 1198. [Google Scholar] [CrossRef]
- Bai, Y.C.; Chang, Y.Y.; Hussain, M.; Lu, B.; Zhang, J.P.; Song, X.B.; Lei, X.S.; Pei, D. Soil Chemical and Microbiological Properties Are Changed by Long-Term Chemical Fertilizers That Limit Ecosystem Functioning. Microorganisms 2020, 8, 694. [Google Scholar] [CrossRef]
- Sallam, A.; Alqudah, A.M.; Dawood, M.F.A.; Baenziger, P.S.; Börner, A. Drought Stress Tolerance in Wheat and Barley: Advances in Physiology, Breeding and Genetics Research. Int. J. Mol. Sci. 2019, 20, 3137. [Google Scholar] [CrossRef] [PubMed]
- Ait-Rahou, Y.; Boutaj, H.; Boutasknit, A.; Douira, A.; Benkirane, R.; El Modafar, C.; Abdelilah, M. Colonization of Tomato Roots with Arbuscular Mycorrhizal Fungi Changes of Antioxidative Activity and Improves Tolerance to Verticillium dahliae. Plant Cell Biotechnol. Mol. Biol. 2021, 22, 65–81. [Google Scholar]
- Ouhaddou, R.; Ech-chatir, L.; Anli, M.; Ben-Laouane, R.; Boutasknit, A.; Meddich, A. Secondary Metabolites, Osmolytes and Antioxidant Activity as the Main Attributes Enhanced by Biostimulants for Growth and Resilience of Lettuce to Drought Stress. Gesunde Pflanz. 2023, 75, 1737–1753. [Google Scholar] [CrossRef]
- Arif, Y.; Singh, P.; Siddiqui, H.; Bajguz, A.; Hayat, S. Plant Physiology and Biochemistry Salinity Induced Physiological and Biochemical Changes in Plants: An Omic Approach towards Salt Stress Tolerance. Plant Physiol. Biochem. 2020, 156, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Majidian, P.; Ghorbani, H. Salinity Stress in Plants: Challenges in View of Physiological Aspects. In Abiotic Stress in Crop Plants—Ecophysiological Responses and Molecular Approaches; IntechOpen: Boston, MA, USA, 2024. [Google Scholar]
- Secomandi, E.; De Gregorio, M.A.; Castro-Cegrí, A.; Lucini, L. Biochemical, Photosynthetic and Metabolomics Insights of Single and Combined Effects of Salinity, Heat, Cold and Drought in Arabidopsis. Physiol. Plant. 2025, 177, e70062. [Google Scholar] [CrossRef]
- Fischer, S.; Wilckens, R.; Jara, J.; Aranda, M.; Valdivia, W.; Bustamante, L.; Graf, F.; Obal, I. Protein and Antioxidant Composition of Quinoa (Chenopodium quinoa Willd.) Sprout from Seeds Submitted to Water Stress, Salinity and Light Conditions. Ind. Crops Prod. 2017, 107, 558–564. [Google Scholar] [CrossRef]
- Anwaar, H.A.; Perveen, R.; Mansha, M.Z.; Abid, M.; Sarwar, Z.M.; Aatif, H.M.; Umar, U.U.D.; Sajid, M.; Aslam, H.M.U.; Alam, M.M.; et al. Assessment of Grain Yield Indices in Response to Drought Stress in Wheat (Triticum aestivum L.). Saudi J. Biol. Sci. 2020, 27, 1818–1823. [Google Scholar] [CrossRef]
- Chakhchar, A.; Haworth, M.; El Modafar, C.; Lauteri, M.; Mattioni, C.; Wahbi, S.; Centritto, M. An Assessment of Genetic Diversity and Drought Tolerance in Argan Tree (Argania spinosa) Populations: Potential for the Development of Improved Drought Tolerance. Front. Plant Sci. 2017, 8, 276. [Google Scholar] [CrossRef]
- Arenas-Castro, S.; Gonçalves, J.F.; Moreno, M.; Villar, R. Projected Climate Changes Are Expected to Decrease the Suitability and Production of Olive Varieties in Southern Spain. Sci. Total Environ. 2020, 709, 136161. [Google Scholar] [CrossRef] [PubMed]
- Zaib, M.; Zeeshan, A.; Aslam, S.; Bano, S.; Ilyas, A.; Abbas, Z.; Nazar, A.; Mumtaz, S. Drought Stress and Plants Production: A Review with Future Prospects. Int. J. Sci. Res. Eng. Dev. 2023, 6, 1278–1293. [Google Scholar]
- Janni, M.; Maestri, E.; Gullì, M.; Marmiroli, M.; Marmiroli, N. Plant Responses to Climate Change, How Global Warming May Impact on Food Security: A Critical Review. Front. Plant Sci. 2023, 14, 1297569. [Google Scholar] [CrossRef]
- Ayed, S.; Othmani, A.; Bouhaouel, I.; Teixeira da Silva, J.A. Multi-Environment Screening of Durum Wheat Genotypes for Drought Tolerance in Changing Climatic Events. Agronomy 2021, 11, 875. [Google Scholar] [CrossRef]
- Del Pozo, A.; Brunel-Saldias, N.; Engler, A.; Ortega-Farias, S.; Acevedo-Opazo, C.; Lobos, G.A.; Jara-Rojas, R.; Molina-Montenegro, M.A. Climate Change Impacts and Adaptation Strategies of Agriculture in Mediterranean-Climate Regions (MCRs). Sustainability 2019, 11, 2769. [Google Scholar] [CrossRef]
- Zaremehrjardy, M.; Victor, J.; Park, S.; Smerdon, B.; Alessi, D.S.; Faramarzi, M. Assessment of Snowmelt and Groundwater-Surface Water Dynamics in Mountains, Foothills, and Plains Regions in Northern Latitudes. J. Hydrol. 2022, 606, 127449. [Google Scholar] [CrossRef]
- Maghsoudi, K.; Emam, Y.; Pessarakli, M. Effect of Silicon on Photosynthetic Gas Exchange, Photosynthetic Pigments, Cell Membrane Stability and Relative Water Content of Different Wheat Cultivars under Drought Stress Conditions. J. Plant Nutr. 2016, 39, 1001–1015. [Google Scholar] [CrossRef]
- Soussani, F.E.; Boutasknit, A.; Ben-Laouane, R.; Benkirane, R.; Baslam, M.; Meddich, A. Arbuscular Mycorrhizal Fungi and Compost-Based Biostimulants Enhance Fitness, Physiological Responses, Yield, and Quality Traits of Drought-Stressed Tomato Plants. Plants 2023, 12, 1856. [Google Scholar] [CrossRef]
- Boutasknit, A.; Baslam, M.; Anli, M.; Ait-El-Mokhtar, M.; Ben-Laouane, R.; Ait-Rahou, Y.; El Modafar, C.; Douira, A.; Wahbi, S.; Meddich, A. Impact of Arbuscular Mycorrhizal Fungi and Compost on the Growth, Water Status, and Photosynthesis of Carob (Ceratonia siliqua) under Drought Stress and Recovery. Plant Biosyst. 2021, 156, 994–1010. [Google Scholar] [CrossRef]
- Boutasknit, A.; Baslam, M.; Ait-El-mokhtar, M.; Anli, M.; Ben-Laouane, R.; Douira, A.; Modafar, C.E.; Mitsui, T.; Wahbi, S.; Meddich, A. Arbuscular Mycorrhizal Fungi Mediate Drought Tolerance and Recovery in Two Contrasting Carob (Ceratonia siliqua L.) Ecotypes by Regulating Stomatal, Water Relations, and (in)Organic Adjustments. Plants 2020, 9, 80. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Parvin, K.; Bhuiyan, T.F.; Anee, T.I.; Nahar, K.; Hossen, M.S.; Zulfiqar, F.; Alam, M.M.; Fujita, M. Regulation of ROS Metabolism in Plants under Environmental Stress: A Review of Recent Experimental Evidence. Int. J. Mol. Sci. 2020, 21, 8695. [Google Scholar] [CrossRef]
- He, J.D.; Zou, Y.N.; Wu, Q.S.; Kuča, K. Mycorrhizas Enhance Drought Tolerance of Trifoliate Orange by Enhancing Activities and Gene Expression of Antioxidant Enzymes. Sci. Hortic. 2020, 262, 108745. [Google Scholar] [CrossRef]
- Schwarzott, D.; Walker, C.; Schu, A. Glomus, the Largest Genus of the Arbuscular Mycorrhizal Fungi (Glomales), Is Nonmonophyletic. Mol. Phylogenet. Evol. 2001, 21, 190–197. [Google Scholar] [CrossRef]
- Singh, G.; Kaur, M.; Kaushik, P.; Nasser, M.; Abdullah, A.; Ahmad, P. Arbuscular Mycorrhiza in Combating Abiotic Stresses in Vegetables: An Eco-Friendly Approach. Saudi J. Biol. Sci. 2021, 28, 1465–1476. [Google Scholar] [CrossRef]
- Sarkodee-addo, E.; Yasuda, M.; Lee, C.G.; Kanasugi, M.; Fujii, Y.; Omari, R.A.; Abebrese, S.O.; Bam, R.; Asuming-brempong, S.; Mohammad, K.; et al. Arbuscular Mycorrhizal Fungi Associated with Rice (Oryza sativa L.) in Ghana: Effet of Regional Locations and Soil Factors on Diversity and Community Assembly. Agronomy 2020, 10, 559. [Google Scholar] [CrossRef]
- Datta, K.; Chakraborty, S.; Roychoudhury, A. Management of Soil, Waste and Water in the Context of Global Climate Change. In Environmental Nexus for Resource Management; CRC Press: Boca Raton, FL, USA, 2024; pp. 1–26. [Google Scholar]
- Ikehi, M.E.; Ifeanyieze, F.O.; Onu, F.M.; Ejiofor, T.E.; Nwankwo, C.U. Assessing Climate Change Mitigation and Adaptation Strategies and Agricultural Innovation Systems in the Niger Delta. GeoJournal 2023, 88, 209–224. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Basche, A.; Traylor, E.; Roy, T. The Efficacy of Conservation Practices in Reducing Floods and Improving Water Quality. Front. Environ. Sci. 2023, 11, 1136989. [Google Scholar] [CrossRef]
- Khan, S.; Irshad, S.; Mehmood, K.; Hasnain, Z.; Nawaz, M.; Rais, A.; Gul, S.; Wahid, M.A.; Hashem, A.; Abd_Allah, E.F.; et al. Biochar Production and Characteristics, Its Impacts on Soil Health, Crop Production, and Yield Enhancement: A Review. Plants 2024, 13, 166. [Google Scholar] [CrossRef] [PubMed]
- Anand, U.; Pal, T.; Yadav, N.; Singh, V.K.; Tripathi, V.; Choudhary, K.K.; Shukla, A.K.; Sunita, K.; Kumar, A.; Bontempi, E.; et al. Current Scenario and Future Prospects of Endophytic Microbes: Promising Candidates for Abiotic and Biotic Stress Management for Agricultural and Environmental Sustainability; Springer: Berlin/Heidelberg, Germany, 2023; Volume 86, ISBN 0123456789. [Google Scholar]
- Abbaspour, H.; Saeidi-Sar, S.; Afshari, H.; Abdel-Wahhab, M.A. Tolerance of Mycorrhiza Infected Pistachio (Pistacia vera L.) Seedling to Drought Stress under Glasshouse Conditions. J. Plant Physiol. 2012, 169, 704–709. [Google Scholar] [CrossRef]
- Meddich, A.; Oihabi, A.; Abbas, Y.; Bizid, E. Rôle des Champignons Mycorhiziens à Arbuscules de Zones Arides dans La Résistance du Trèfle (Trifolium alexandrinum L.) au Déficit Hydrique. Agronomie 2000, 20, 283–295. [Google Scholar] [CrossRef]
- Hartoyo, B.; Trisilawati, O. Diversity of Arbuscular Mycorrhiza Fungi (AMF) in the Rhizosphere of Sugarcane. IOP Conf. Ser. Earth Environ. Sci. 2021, 653, 012066. [Google Scholar] [CrossRef]
- Meddich, A.; El Mokhtar, M.A.; Wahbi, S.; Boumezzough, A. Évaluation des Potentialités Mycorhizogènes en Lien Avec les Paramètres Physico-Chimiques des Sols de Palmeraies du Maroc (Marrakech et Tafilalet). Cah. Agric. 2017, 26, 45012. [Google Scholar] [CrossRef]
- Afrangan, F.; Kazemeini, S.A.; Alinia, M.; Mastinu, A. Glomus versiforme and Micrococcus yunnanensis Reduce the Negative Effects of Salinity Stress by Regulating the Redox State and Ion Homeostasis in Brassica napus L. Crops. Biologia 2023, 78, 3049–3061. [Google Scholar] [CrossRef]
- Alarcón-Zayas, A.; Hernández-Montiel, L.G.; Medina-Hernández, D.; Rueda-Puente, E.O.; Ceiro-Catasú, W.G.; Holguín-Peña, R.J. Effects of Glomus fasciculatum, Azotobacter chroococcum and Vermicompost Leachate on the Production and Quality of Tomato Fruit. Microbiol. Res. 2024, 15, 187–195. [Google Scholar] [CrossRef]
- Msairi, S.; Rais, C.; Maazouzi, S.; Artib, M.; El Gabardi, S.; Mouden, N.; Selmaoui, K.; Benkirane, R.; Ouazzani Touhami, A.; Douira, A. Arbuscular Mycorrhizal Symbiosis in Two Cannabis Varieties (Khardala and Critical) in Morocco. Ecol. Eng. Environ. Technol. 2023, 24, 30–35. [Google Scholar] [CrossRef]
- Belechheb, T. Mycorrhizal Fungi Status Associated with the Rhizosphere of Cytisus monspessulanus in the North West of Morocco. Int. J. Pure Appl. Biosci. 2016, 4, 1–8. [Google Scholar] [CrossRef]
- Kachkouch, W.; Touhami, A.O.; Filali-maltouf, A.; El Modafar, C.; Moukhli, A.; Oukabli, A.; Benkirane, R.; Douira, A. Diversity of Arbuscular Mycorrhizal Fungi in the Rhizosphere of Olea europaea in Three Regions of Morocco (Tafilalt, Zagora and Taounate). Int. J. Pure Appl. Biosci. 2014, 2, 178–195. [Google Scholar]
- Lahbouki, S.; Anli, M.; El Gabardi, S.; Ait-El-Mokhtar, M.; Ben-Laouane, R.; Boutasknit, A.; Ait-Rahou, Y.; Outzourhit, A.; Wahbi, S.; Douira, A.; et al. Evaluation of Arbuscular Mycorrhiost Supplementation on Growth, Phenolic Content and Antioxidant Activity of Prickly Pear Cactus (Opuntia ficus-indica). Plant Biosyst. 2021, 156, 882–892. [Google Scholar] [CrossRef]
- Magurno, F.; Uszok, S.; Bierza, K.; Bakr, J.; Kende, Z.; Bessa de Queiroz, M.; Goto, B.T. Glomus highlandensis and G. mongioie, Two New Arbuscular Mycorrhizal Fungi from Saltmarshes, Dunes and Mountains of Europe. Ecol. Evol. Behav. Syst. 2024. [Google Scholar] [CrossRef]
- Błaszkowski, J.; Kovács, G.M.; Balázs, T.K.; Orłowska, E.; Sadravi, M.; Wubet, T.; Buscot, F. Glomus africanum and G. iranicum, Two New Species of Arbuscular Mycorrhizal Fungi (Glomeromycota). Mycologia 2010, 102, 1450–1462. [Google Scholar] [CrossRef]
- Stürmer, S.L.; Kemmelmeier, K. The Glomeromycota in the Neotropics. Front. Microbiol. 2021, 11, 553679. [Google Scholar] [CrossRef]
- Arif, S.; YusnainI, S.; Niswati, A.; Setiawan, A.; Tuchida, K.; Katou, T.; Touji, Y.; Nonaka, M. Population of Arbuscular Mycorrhizal Fungi (AMF) by Different Land Use in Sumatra, Indonesia. Comparison of AMF Spore Numbers in Primary Forest, Secondary Forest, Fields Growing Coffee and Native Grass. Microbes Environ. 1999, 14, 9–17. [Google Scholar] [CrossRef]
- Gerdemann, J.W.; Trappe, J.M. The Endogonaceae in the Pacific Northwest. Mycol. Mem. 1974, 5, 76. [Google Scholar]
- Morton, J.B.; Benny, G.L. Revised Classification of Arbuscular Mycorrhizal Fungi (Zygomycetes): A New Order, Glomales, Two New Suborders, Glomineae and Gigasporineae, and Two New Families, Acaulosporaceae and Gigasporaceae, with an Emendation of Glomaceae. Mycotaxon 1990, 37, 471–491. [Google Scholar]
- Droh, G.; Meliton Djezou, K.; Tuo, S.; Touré, M.; Kouassi, A.-B. Morphometric Characterization of Endomycorrhizal Fungi (Glomeraceae and Acaulosporaceae) from the Bouaflé and Niellé Areas in Côte D'Ivoire. Am. J. Biosci. 2023, 11, 1–10. [Google Scholar] [CrossRef]
- Schüßler, A.; Walker, C. The Glomeromycota: A Species List with New Families and Genera; CreateSpace Independent Publishing Platform: North Charleston, SC, USA, 2010. [Google Scholar]
- Oehl, F.; Goto, B.T.; Sieverding, E. Glomeromycota: Three New Genera and Glomoid Species Reorganized. Mycotaxon 2011, 116, 75–120. [Google Scholar] [CrossRef]
- Magurno, F.; Uszok, S.; Bierza, K.; Bakr, J.; Kende, Z.; Bessa de Queiroz, M.; Casieri, L. Glomus mongioiense, a New Species of Arbuscular Mycorrhizal Fungi from Italian Alps and the Phylogeny-Spoiling Issue of Ribosomal Variants in the Glomus Genus. Agronomy 2024, 14, 1350. [Google Scholar] [CrossRef]
- Souza, T. An Old Relationship. Handbook of Arbuscular Mycorrhizal Fungi; Springer: Cham, Switzerland, 2015; pp. 9–41. ISBN 9783319248509. [Google Scholar]
- Stürmer, S.L. A History of the Taxonomy and Systematics of Arbuscular Mycorrhizal Fungi Belonging to the Phylum Glomeromycota. Mycorrhiza 2012, 22, 247–258. [Google Scholar] [CrossRef]
- Pirozynski, K.A.; Dalpe, Y. Geological History of the Glomaceae with Particular Reference to Mycorrhizal Symbiosis. Symbiosis 1989, 7, 1–36. [Google Scholar]
- Taylor, A.T.N.; Remy, W.; Hass, H.; Kerp, H.; Mycologia, S.; Aug, J.; Aug, N.J. Fossil Arbuscular Mycorrhizae from the Early Devonian. Mycologia 1995, 87, 560–573. [Google Scholar] [CrossRef]
- Redecker, D.; Redecker, D.; Kodner, R.; Graham, L.E. Glomalean Fungi from the Ordovician. Science 2000, 289, 1920–1921. [Google Scholar] [CrossRef]
- Bouwmeester, H.J.; Roux, C.; Lopez-raez, J.A. Rhizosphere Communication of Plants, Parasitic Plants and AM Fungi. Trends Plant Sci. 2007, 12, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Pérez-López, U.; Miranda-Apodaca, J.; Muñoz-Rueda, A.; Mena-Petite, A. Lettuce Production and Antioxidant Capacity Are Differentially Modified by Salt Stress and Light Intensity under Ambient and Elevated CO2. J. Plant Physiol. 2013, 170, 1517–1525. [Google Scholar] [CrossRef] [PubMed]
- Błaszkowski, J.; Czerniawska, B. Glomus eburneum and Scutellospora fulgida, Species of Arbuscular Mycorrhizal Fungi (Glomeromycota) New for Europe. Acta Mycol. 2013, 43, 57–65. [Google Scholar] [CrossRef]
- Priyadharsini, P.; Muthukumar, T.; Pandey, R.R. Arbuscular Mycorrhizal and Dark Septate Fungal Associations in Shallot (Allium cepa L. Var. Aggregatum) under Conventional Agriculture. Acta Bot. Croat. 2012, 71, 159–175. [Google Scholar] [CrossRef]
- Dalpé, Y.; de Souza, F.A.; Declerck, S. Life Cycle of Glomus Species in Monoxenic Culture. In In Vitro Culture of Mycorrhizas; Springer: Berlin/Heidelberg, Germany, 2005; pp. 49–71. [Google Scholar] [CrossRef]
- Acoltzi-conde, M.C.; Chimal-sánchez, E.; Tovar-soto, A.; Díaz-reyes, J. Arbuscular Mycorrhizal Fungi Consortia in Six Vegetable Crops in the Tepeaca Valley, Puebla, Mexico. Rev. Terra Latinoam. 2024, 42, 1–11. [Google Scholar] [CrossRef]
- Retama-Ortiz, Y.; Ávila-Bello, C.H.; Alarcón, A.; Ferrera-Cerrato, R. Effectiveness of Native Arbuscular Mycorrhiza on the Growth of Four Tree Forest Species from the Santa Marta Mountain, Veracruz (Mexico). For. Syst. 2017, 26, e001. [Google Scholar] [CrossRef]
- Noreen, S.; Yaseen, T.; Iqbal, J.; Abbasi, B.A.; Farouk Elsadek, M.; Eldin, S.M.; Ijaz, S.; Ali, I. Morphological and Molecular Characterizations of Arbuscular Mycorrhizal Fungi and Their Influence on Soil Physicochemical Properties and Plant Nutrition. ACS Omega 2023, 8, 32468–32482. [Google Scholar] [CrossRef]
- Scervino, J.M.; Ponce, M.A.; Erra-bassells, R.; Vierheilig, H. Flavonoids Exhibit Fungal Species and Genus Specific Effects on the Presymbiotic Growth of Gigaspora and Glomus. Mycol. Res. 2005, 109, 789–794. [Google Scholar] [CrossRef] [PubMed]
- Liao, D.; Sun, C.; Li, S.; Tauqeer, A.; Bian, X.; Shen, J.; Wu, S. The Utilization and Molecular Mechanism of Arbuscular Mycorrhizal Symbiosis in Vegetables. Veg. Res. 2023, 3, 1–7. [Google Scholar] [CrossRef]
- Naseer, M.A.; Zhang, Z.Q.; Mukhtar, A.; Asad, M.S.; Wu, H.Y.; Yang, H.; Zhou, X.B. Strigolactones: A Promising Tool for Nutrient Acquisition through Arbuscular Mycorrhizal Fungi Symbiosis and Abiotic Stress Tolerance. Plant Physiol. Biochem. 2024, 215, 109057. [Google Scholar] [CrossRef] [PubMed]
- Yolanda, N.-G.; Ferrera-Cerrato, R.; Santamaría, J.M. Glomus intraradices Attenuates the Negative Effect of Low Pi Supply on Photosynthesis and Growth of Papaya Maradol Plants. J. Bot. 2012, 2012, 129591. [Google Scholar] [CrossRef]
- Steinkellner, S.; Lendzemo, V.; Langer, I.; Schweiger, P.; Khaosaad, T.; Toussaint, J.P.; Vierheilig, H. Flavonoids and Strigolactones in Root Exudates as Signals in Symbiotic and Pathogenic Plant-Fungus Interactions. Molecules 2007, 12, 1290–1306. [Google Scholar] [CrossRef]
- Gust, A.A.; Willmann, R.; Desaki, Y.; Grabherr, H.M.; Nu, T. Plant LysM Proteins: Modules Mediating Symbiosis and Immunity. Trends Plant Sci. 2012, 17, 495–502. [Google Scholar] [CrossRef]
- Nagahashi, G.; Douds, D.D. The Effects of Hydroxy Fatty Acids on the Hyphal Branching of Germinated Spores of AM Fungi. Fungal Biol. 2011, 115, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Azmat, R.; Hamid, N.; Moin, S.; Saleem, A. Glomus fasciculatum Fungi as a Bio-Convertor and Bio-Activator of Inorganic and Organic P in Dual Symbiosis. Recent Pat. Biotechnol. 2015, 9, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Wewer, V.; Brands, M.; Dörmann, P. Fatty Acid Synthesis and Lipid Metabolism in the Obligate Biotrophic Fungus Rhizophagus irregularis during Mycorrhization of Lotus japonicus. Plant J. 2014, 79, 398–412. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, A.M.; Harrison, M.J. Signaling Events during Initiation of Arbuscular Mycorrhizal Symbiosis. J. Integr. Plant Biol. 2014, 56, 250–261. [Google Scholar] [CrossRef]
- Nadal, M.; Sawers, R.; Naseem, S.; Bassin, B.; Kulicke, C.; Sharman, A.; An, G.; An, K.; Ahern, K.R.; Romag, A.; et al. An N -Acetylglucosamine Transporter Required for Arbuscular Mycorrhizal Symbioses in Rice and Maize. Nat. Plants. 2017, 3, 17073. [Google Scholar] [CrossRef] [PubMed]
- Steinkellner, S.; Vierheilig, H.; Antonio, J.; Bote, O. Abscisic Acid Determines Arbuscule Development and Functionality in the Tomato Arbuscular Mycorrhiza. New Phytol. 2007, 175, 554–564. [Google Scholar] [CrossRef]
- Feng, F.; Sun, J.; Radhakrishnan, G.V.; Lee, T.; Bozsóki, Z.; Fort, S.; Gavrin, A.; Gysel, K.; Thygesen, M.B.; Andersen, K.R.; et al. A Combination of Chitooligosaccharide and Lipochitooligosaccharide Recognition Promotes Arbuscular Mycorrhizal Associations in Medicago truncatula. Nat. Commun. 2019, 10, 5047. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.R.; Füchtbauer, W.; Novero, M.; Volpe, V.; Malkov, N.; Genre, A.; Bonfante, P.; Stougaard, J.; Radutoiu, S. Intraradical Colonization by Arbuscular Mycorrhizal Fungi Triggers Induction of a Lipochitooligosaccharide Receptor. Sci. Rep. 2016, 6, 29733. [Google Scholar] [CrossRef]
- Zhang, X.; Dong, W.; Sun, J.; Feng, F.; Deng, Y.; He, Z.; Oldroyd, G.E.D.; Wang, E. The Receptor Kinase CERK1 Has Dual Functions in Symbiosis and Immunity Signalling. Plant J. 2015, 81, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Stracke, S.; Kistner, C.; Yoshida, S.; Mulder, L.; Sato, S.; Kaneko, T.; Tabata, S.; Sandal, N.; Stougaard, J.; Szczyglowski, K.; et al. A Plant Receptor-like Kinase Required for Both Bacterial and Fungal Symbiosis. Nature 2002, 417, 959–962. [Google Scholar] [CrossRef] [PubMed]
- Charpentier, M.; Sun, J.; Martins, T.V.; Radhakrishnan, G.V.; Findlay, K.; Soumpourou, E.; Thouin, J.; Véry, A.; Sanders, D.; Morris, R.J.; et al. Nuclear-Localized Cyclic Nucleotide–Gated Channels Mediate Symbiotic Calcium Oscillations. Science 2016, 352, 1102–1105. [Google Scholar] [CrossRef]
- Yano, K.; Yoshida, S.; Müller, J.; Singh, S.; Banba, M.; Vickers, K.; Markmann, K.; White, C.; Schuller, B.; Sato, S.; et al. CYCLOPS, a Mediator of Symbiotic Intracellular Accommodation. Proc. Natl. Acad. Sci. USA 2008, 105, 20540–20545. [Google Scholar] [CrossRef]
- Etemadi, M.; Gutjahr, C.; Couzigou, J.M.; Zouine, M.; Lauressergues, D.; Timmers, A.; Audran, C.; Bouzayen, M.; Bécard, G.; Combier, J.P. Auxin Perception Is Required for Arbuscule Development in Arbuscular Mycorrhizal Symbiosis. Plant Physiol. 2014, 166, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Ren, A.T.; Zhu, Y.; Chen, Y.L.; Ren, H.X.; Li, J.Y.; Kay Abbott, L.; Xiong, Y.C. Arbuscular Mycorrhizal Fungus Alters Root-Sourced Signal (Abscisic Acid) for Better Drought Acclimation in Zea mays L. Seedlings. Environ. Exp. Bot. 2019, 167, 103824. [Google Scholar] [CrossRef]
- Rich, M.K.; Schorderet, M.; Bapaume, L.; Falquet, L.; Morel, P.; Vandenbussche, M.; Reinhardt, D. The Petunia GRAS Transcription Factor ATA/RAM1 Regulates Symbiotic Gene Expression and Fungal Morphogenesis in Arbuscular Mycorrhiza. Plant Physiol. 2015, 168, 788–797. [Google Scholar] [CrossRef]
- Hossain, M.S.; Dietz, K.J. Tuning of Redox Regulatory Mechanisms, Reactive Oxygen Species and Redox Homeostasis under Salinity Stress. Front. Plant Sci. 2016, 7, 548. [Google Scholar] [CrossRef] [PubMed]
- Ouhaddou, R.; Meddich, A.; Ikan, C.; Lahlali, R.; Ait Barka, E.; Hajirezaei, M.R.; Duponnois, R.; Baslam, M. Enhancing Maize Productivity and Soil Health under Salt Stress through Physiological Adaptation and Metabolic Regulation Using Indigenous Biostimulants. Plants 2023, 12, 3703. [Google Scholar] [CrossRef] [PubMed]
- Kreslavski, V.D.; Los, D.A.; Allakhverdiev, S.I.; Kuznetsov, V.V. Signaling Role of Reactive Oxygen Species in Plants under Stress. Russ. J. Plant Physiol. 2012, 59, 141–154. [Google Scholar] [CrossRef]
- Babar, M.; Saif-ur-Rehman; Rasul, S.; Aslam, K.; Abbas, R.; Athar, H.U.R.; Manzoor, I.; Kashif Hanif, M.; Naqqash, T. Mining of Halo-Tolerant Plant Growth Promoting Rhizobacteria and Their Impact on Wheat (Triticum aestivum L.) under Saline Conditions. J. King Saud Univ.-Sci. 2021, 33, 101372. [Google Scholar] [CrossRef]
- Lu, C.; Li, L.; Liu, X.; Chen, M.; Wan, S.; Li, G. Salt Stress Inhibits Photosynthesis and Destroys Chloroplast Structure by Downregulating Chloroplast Development–Related Genes in Robinia pseudoacacia Seedlings. Plants 2023, 12, 1283. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive Oxygen Species (ROS) and Response of Antioxidants as ROS-Scavengers during Environmental Stress in Plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Hu, Y.; Zhao, H.; Xue, L.; Nie, N.; Zhang, H.; Zhao, N.; He, S.; Liu, Q.; Gao, S.; Zhai, H. IbMYC2 Contributes to Salt and Drought Stress Tolerance via Modulating Anthocyanin Accumulation and ROS-Scavenging System in Sweet Potato. Int. J. Mol. Sci. 2024, 25, 2096. [Google Scholar] [CrossRef]
- Sairam, R.K.; Rao, K.; Srivastava, G.C. Differential Response of Wheat Genotypes to Long Term Salinity Stress in Relation to Oxidative Stress, Antioxidant Activity and Osmolyte Concentration. Plant Sci. 2002, 163, 1037–1046. [Google Scholar] [CrossRef]
- Sahab, S.; Suhani, I.; Srivastava, V.; Chauhan, P.S.; Singh, R.P.; Prasad, V. Potential Risk Assessment of Soil Salinity to Agroecosystem Sustainability: Current Status and Management Strategies. Sci. Total Environ. 2020, 764, 144164. [Google Scholar] [CrossRef]
- Sahin, U.; Ekinci, M.; Ors, S.; Turan, M.; Yildiz, S.; Yildirim, E. Effects of Individual and Combined Effects of Salinity and Drought on Physiological, Nutritional and Biochemical Properties of Cabbage (Brassica oleracea Var. Capitata). Sci. Hortic. 2018, 240, 196–204. [Google Scholar] [CrossRef]
- Marzec, M.; Daszkowska-Golec, A.; Collin, A.; Melzer, M.; Eggert, K.; Szarejko, I. Barley Strigolactone Signalling Mutant Hvd14.d Reveals the Role of Strigolactones in Abscisic Acid-Dependent Response to Drought. Plant Cell Environ. 2020, 43, 2239–2253. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Reichheld, J.P.; Foyer, C.H. ROS-Related Redox Regulation and Signaling in Plants. Semin. Cell Dev. Biol. 2018, 80, 3–12. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef]
- Panda, S.K.; Gupta, D.; Patel, M.; Vyver, C.V.D.; Koyama, H. Functionality of Reactive Oxygen Species (ROS) in Plants: Toxicity and Control in Poaceae Crops Exposed to Abiotic Stress. Plants 2024, 13, 2071. [Google Scholar] [CrossRef]
- Jafari, M.; Shahsavar, A.R.; Talebi, M.; Hesami, M. Exogenous Melatonin Protects Lime Plants from Drought Stress-Induced Damage by Maintaining Cell Membrane Structure, Detoxifying ROS and Regulating Antioxidant Systems. Horticulturae 2022, 8, 257. [Google Scholar] [CrossRef]
- Dandan, Z.; Zhiwei, Z. Biodiversity of Arbuscular Mycorrhizal Fungi in the Hot-Dry Valley of the Jinsha River, Southwest China. Appl. Soil Ecol. 2007, 37, 118–128. [Google Scholar] [CrossRef]
- Jefwa, J.M.; Mung’atu, J.; Okoth, P.; Muya, E.; Roimen, H.; Njuguini, S. Influence of Land Use Types on Occurrence of Arbuscular Mycorrhizal Fungi in the High Altitude Regions of Mt. Kenya. Trop. Subtrop. Agroecosyst. 2009, 11, 277–290. [Google Scholar]
- Abdelhalim, T.S.; Finckh, M.R.; Babiker, A.G.; Oehl, F. Species Composition and Diversity of Arbuscular Mycorrhizal Fungi in White Nile state, Central Sudan. Arch. Agron. Soil Sci. 2014, 60, 37–41. [Google Scholar] [CrossRef]
- Mahmoudi, N.; Cruz, C.; Mahdhi, M.; Mars, M.; Caeiro, M.F. Arbuscular Mycorrhizal Fungi in Soil, Roots and Rhizosphere of Medicago truncatula: Diversity and Heterogeneity under Semi-Arid Conditions. PeerJ 2019, 7, e6401. [Google Scholar] [CrossRef] [PubMed]
- Sellal, Z.; Ouazzani, T.A.; Dahmani, J.; Maazouzi, S.; Najoua, M.; Mohamed, C.; Karima, S.; Rachid, B.; Cherkaoui, E.M.; Douira, A. Diversity of Arbuscular Mycorrhizal Fungi in the Rhizosphere of Argania spinosa in Morocco. In Arbuscular Mycorrhizal Fungi in Agriculture-New Insights; IntechOpen: Boston, MA, USA, 2022. [Google Scholar] [CrossRef]
- Bâ, A.M.; Guissou, T.; Dalpé, Y. Les Glomales d’Acacia Holosericea et d’Acacia Mangium. Bois For. Trop. 1996, 250, 5–17. [Google Scholar]
- Yooyongwech, S.; Phaukinsang, N.; Cha-um, S.; Supaibulwatana, K. Arbuscular Mycorrhiza Improved Growth Performance in Macadamia tetraphylla L. Grown under Water Deficit Stress Involves Soluble Sugar and Proline Accumulation. Plant Growth Regul. 2013, 69, 285–293. [Google Scholar] [CrossRef]
- Mena-Violante, H.G.; Ocampo-Jiménez, O.; Dendooven, L.; Martínez-soto, G.; González-castañeda, J.; Fred, T.D.J.; Olalde-Portugal, V. Arbuscular Mycorrhizal Fungi Enhance Fruit Growth and Quality of Chile Ancho (Capsicum annuum L. Cv San Luis) Plants Exposed to Drought. Mycorrhiza 2006, 16, 261–267. [Google Scholar] [CrossRef]
- Gobat, J.M.; Aragno, M.; Matthey, W. The Living Soil: Basic Pedology-Soil Biology; Presses Polytechniques et Universitaires Romandes: Lausanne, Switzerland, 2003; p. 569. ISBN 2-88074-501-2. [Google Scholar]
- Frey, S.D. Mycorrhizal Fungi as Mediators of Soil Organic Matter Dynamics. Annu. Rev. Ecol. Evol. Syst. 2019, 50, 237–259. [Google Scholar] [CrossRef]
- Al-Maliki, S.; Al-Amery, A.; Sallal, M.; Radhi, A.; Al-Taey, D.K.A. Effects of Arbuscular Mycorrhiza and Organic Wastes on Soil Carbon Mineralisation, Actinomycete s and Nutrient Content in Maize Plants (Zea mays L.). Malays. J. Soil Sci. 2021, 25, 107–124. [Google Scholar]
- Boutasknit, A.; Ait-El-Mokhtar, M.; Fassih, B.; Ben-Laouane, R.; Wahbi, S.; Meddich, A. Effect of Arbuscular Mycorrhizal Fungi and Rock Phosphate on Growth, Physiology, and Biochemistry of Carob under Water Stress and after Rehydration in Vermicompost-Amended Soil. Metabolites 2024, 14, 202. [Google Scholar] [CrossRef]
- Putri, D.A.L.P.; Widyastuti, R.; Idris, I.; Ikhwani, A.Z.N.; Nugroho, S.; Sudiana, I.M.; Kanti, A.; Purnaningsih, I.; Ochiai, K.; Kobayashi, M.; et al. Unraveling the Mechanisms of Drought Tolerance Enhancement in Sorghum Bicolor through Glomus mosseae Inoculation: Insights from Comparative Analysis of Super 2 and Konawe Selatan Accessions. S. Afr. J. Bot. 2023, 161, 293–304. [Google Scholar] [CrossRef]
- Benaffari, W.; Boutasknit, A.; Anli, M.; Ait-el-mokhtar, M.; Ait-Rahou, Y.; Ben-Laouane, R.; Ahmed, H.B.; Mitsui, T.; Baslam, M.; Meddich, A. The Native Arbuscular Mycorrhizal Fungi and Vermicompost-Based Organic Amendments Enhance Soil Fertility, Growth Performance, and the Drought Stress Tolerance of Quinoa. Plants 2022, 11, 393. [Google Scholar] [CrossRef] [PubMed]
- Spinoso-Castillo, J.L.; Moreno-Hernández, M.D.R.; Mancilla-Álvarez, E.; Sánchez-Segura, L.; Sánchez-Páez, R.; Bello-Bello, J.J. Arbuscular Mycorrhizal Symbiosis Improves Ex Vitro Acclimatization of Sugarcane Plantlets (Saccharum spp.) under Drought Stress Conditions. Plants 2023, 12, 687. [Google Scholar] [CrossRef] [PubMed]
- Ganjeali, A.; Ashiani, E.; Zare, M.; Tabasi, E. Influences of the Arbuscular Mycorrhizal Fungus Glomus mosseae on Morphophysiological Traits and Biochemical Compounds of Common Bean (Phaseolus vulgaris) under Drought Stress. S. Afr. J. Plant Soil. 2018, 35, 121–127. [Google Scholar] [CrossRef]
- He, F.; Sheng, M.; Tang, M. Effects of Rhizophagus irregularis on Photosynthesis and Antioxidative Enzymatic System in Robinia pseudoacacia L. under Drought Stress. Front. Plant Sci. 2017, 8, 183. [Google Scholar] [CrossRef]
- Meddich, A.; Jaiti, F.; Bourzik, W.; Asli, A.E.; Hafidi, M. Use of Mycorrhizal Fungi as a Strategy for Improving the Drought Tolerance in Date Palm (Phoenix dactylifera). Sci. Hortic. 2015, 192, 468–474. [Google Scholar] [CrossRef]
- Ouledali, S.; Ennajeh, M.; Ferrandino, A.; Khemira, H.; Schubert, A.; Secchi, F. Influence of Arbuscular Mycorrhizal Fungi Inoculation on the Control of Stomata Functioning by Abscisic Acid (ABA) in Drought-Stressed Olive Plants. S. Afr. J. Bot. 2019, 121, 152–158. [Google Scholar] [CrossRef]
- Meddich, A.; Ouhaddou, R.; Anli, M.; Boutasknit, A. Role of Phosphorus and Arbuscular Mycorrhizal Fungi in the Growth Performances and Tolerance of Barley to Water Stress. Plant Cell Biotechnol. Mol. Biol. 2021, 22, 45–67. [Google Scholar]
- Baslam, M.; Goicoechea, N. Water Deficit Improved the Capacity of Arbuscular Mycorrhizal Fungi (AMF) for Inducing the Accumulation of Antioxidant Compounds in Lettuce Leaves. Mycorrhiza 2012, 22, 347–359. [Google Scholar] [CrossRef]
- Wu, Q.S.; Zou, Y.N.; Xia, R.X. Effects of Water Stress and Arbuscular Mycorrhizal Fungi on Reactive Oxygen Metabolism and Antioxidant Production by Citrus (Citrus tangerine) Roots. Eur. J. Soil Biol. 2006, 42, 166–172. [Google Scholar] [CrossRef]
- Begum, N.; Ahanger, M.A.; Zhang, L. AMF Inoculation and Phosphorus Supplementation Alleviates Drought Induced Growth and Photosynthetic Decline in Nicotiana tabacum by Up-Regulating Antioxidant Metabolism and Osmolyte Accumulation. Environ. Exp. Bot. 2020, 176, 104088. [Google Scholar] [CrossRef]
- Ghasemi, M.; Zahedi, M.; Gheysari, M.; Sabzalian, M.R. Effects of Inoculation with Four Mycorrhizal Species on Seed Phenolic and Fatty Acids of Sesame Plants Grown under Different Irrigation Regimes. Sci. Rep. 2023, 13, 16482. [Google Scholar] [CrossRef] [PubMed]
- Del Mar Alguacil, M.; Kohler, J.; Caravaca, F.; Roldán, A. Differential Effects of Pseudomonas mendocina and Glomus intraradices on Lettuce Plants Physiological Response and Aquaporin PIP2 Gene Expression under Elevated Atmospheric CO2 and Drought. Microb. Ecol. 2009, 58, 942–951. [Google Scholar] [CrossRef]
- Das, S.; Sarkar, S. Arbuscular Mycorrhizal Fungal Contribution towards Plant Resilience to Drought Conditions. Front. Fungal Biol. 2024, 5, 1355999. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Zhou, L.J.; Xu, Y.J.; Hashem, A.; Abd_Allah, E.F.; Wu, Q.S. Growth Performance and Osmolyte Regulation of Drought-Stressed Walnut Plants Are Improved by Mycorrhiza. Agriculture 2024, 14, 367. [Google Scholar] [CrossRef]
- Chen, M.; Yang, G.; Sheng, Y.; Li, P.; Qiu, H.; Zhou, X.; Huang, L.; Chao, Z. Glomus mosseae Inoculation Improves the Root System Architecture, Photosynthetic Efficiency and Flavonoids Accumulation of Liquorice under Nutrient Stress. Front. Plant Sci. 2017, 8, 931. [Google Scholar] [CrossRef] [PubMed]
- Sagar, A.; Rathore, P.; Ramteke, P.W.; Ramakrishna, W.; Reddy, M.S.; Pecoraro, L. Plant Growth Promoting Rhizobacteria, Arbuscular Mycorrhizal Fungi and Their Synergistic Interactions to Counteract the Negative Effects of Saline Soil on Agriculture: Key Macromolecules and Mechanisms. Microorganisms 2021, 9, 1491. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.L.; Mooney, S.J.; Dickinson, M.J.; West, H.M. The Effects of Simultaneous Root Colonisation by Three Glomus Species on Soil Pore Characteristics. Soil Biol. Biochem. 2012, 49, 167–173. [Google Scholar] [CrossRef]
- Lei, A.Q.; Yang, Q.H.; Zhang, Y.; Liao, W.Y.; Xie, Y.C.; Srivastava, A.K.; Hashem, A.; Alqahtani, M.D.; Abd_Allah, E.F.; Wu, Q.S.; et al. Agronomic Practices Alter Regulated Effects of Easily Extractable Glomalin-Related Soil Protein on Fruit Quality and Soil Properties of Satsuma Mandarin. Agronomy 2023, 13, 881. [Google Scholar] [CrossRef]
- Singh, A.K.; Zhu, X.; Chen, C.; Wu, J.; Yang, B.; Zakari, S.; Jiang, X.J.; Singh, N.; Liu, W. The Role of Glomalin in Mitigation of Multiple Soil Degradation Problems. Crit. Rev. Environ. Sci. Technol. 2022, 52, 1604–1638. [Google Scholar] [CrossRef]
- El-Sherbeny, T.M.S.; Mousa, A.M.; El-Sayed, E.S.R. Use of Mycorrhizal Fungi and Phosphorus Fertilization to Improve the Yield of Onion (Allium cepa L.) Plant. Saudi J. Biol. Sci. 2022, 29, 331–338. [Google Scholar] [CrossRef]
- Jabborova, D.; Annapurna, K.; Al-Sadi, A.M.; Alharbi, S.A.; Datta, R.; Zuan, A.T.K. Biochar and Arbuscular Mycorrhizal Fungi Mediated Enhanced Drought Tolerance in Okra (Abelmoschus esculentus) Plant Growth, Root Morphological Traits and Physiological Properties. Saudi J. Biol. Sci. 2021, 28, 5490–5499. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Xu, Y.; Han, G.; Wang, W.; Li, X.; Cheng, B. Identification and Functional Characterization of a Maize Phosphate Transporter Induced by Mycorrhiza Formation. Plant Cell Physiol. 2018, 59, 1683–1694. [Google Scholar] [CrossRef] [PubMed]
- Volpe, V.; Chitarra, W.; Cascone, P.; Volpe, M.G.; Bartolini, P.; Moneti, G.; Pieraccini, G.; Serio, C.D.; Maserti, B.; Guerrieri, E.; et al. The Association With Two Different Arbuscular Mycorrhizal Fungi Differently Affects Water Stress Tolerance in Tomato. Front. Plant Sci. 2018, 9, 1480. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Meng, S.; Li, M.; Zhao, Z. Genomic Identification and Expression Analysis of the Phosphate Transporter Gene Family in Poplar. Front. Plant Sci. 2016, 7, 1398. [Google Scholar] [CrossRef]
- Balestrini, R.; Rosso, L.C.; Veronico, P.; Melillo, M.T.; De Luca, F.; Fanelli, E.; Colagiero, M.; Di Fossalunga, A.S.; Ciancio, A.; Pentimone, I. Transcriptomic Responses to Water Deficit and Nematode Infection in Mycorrhizal Tomato Roots. Front. Microbiol. 2019, 10, 1807. [Google Scholar] [CrossRef]
- Al-Arjani, A.B.F.; Hashem, A.; Abd_Allah, E.F. Arbuscular Mycorrhizal Fungi Modulates Dynamics Tolerance Expression to Mitigate Drought Stress in Ephedra foliata Boiss. Saudi J. Biol. Sci. 2020, 27, 380–394. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Zhao, Q.; Zaman, S.; Anwar, A.; Li, S. The Transcriptomic Analysis Revealed the Molecular Mechanism of Arbuscular Mycorrhizal Fungi (AMF) Inoculation in Watermelon. Sci. Hortic. 2024, 332, 113184. [Google Scholar] [CrossRef]
- Bahadur, A.; Batool, A.; Nasir, F.; Jiang, S.; Mingsen, Q.; Zhang, Q.; Pan, J.; Liu, Y.; Feng, H. Mechanistic Insights into Arbuscular Mycorrhizal Fungi-Mediated Drought Stress Tolerance in Plants. Int. J. Mol. Sci. 2019, 20, 4199. [Google Scholar] [CrossRef]
- Xie, W.; Hao, Z.; Zhou, X.; Jiang, X.; Xu, L.; Wu, S.; Zhao, A.; Zhang, X.; Chen, B. Arbuscular Mycorrhiza Facilitates the Accumulation of Glycyrrhizin and Liquiritin in Glycyrrhiza uralensis under Drought Stress. Mycorrhiza 2018, 28, 285–300. [Google Scholar] [CrossRef]
- Sharma, K.; Gupta, S.; Thokchom, S.D.; Jangir, P.; Kapoor, R. Arbuscular Mycorrhiza-Mediated Regulation of Polyamines and Aquaporins During Abiotic Stress: Deep Insights on the Recondite Players. Front. Plant Sci. 2021, 12, 642101. [Google Scholar] [CrossRef]
- El-Mesbahi, M.N.; Azcón, R.; Ruiz-Lozano, J.M.; Aroca, R. Plant Potassium Content Modifies the Effects of Arbuscular Mycorrhizal Symbiosis on Root Hydraulic Properties in Maize Plants. Mycorrhiza 2012, 22, 555–564. [Google Scholar] [CrossRef]
- Gong, M.; Bai, N.; Wang, P.; Su, J.; Chang, Q.; Zhang, Q. Co-Inoculation with Arbuscular Mycorrhizal Fungi and Dark Septate Endophytes under Drought Stress: Synergistic or Competitive Effects on Maize Growth, Photosynthesis, Root Hydraulic Properties and Aquaporins? Plants 2023, 12, 2596. [Google Scholar] [CrossRef] [PubMed]
- Quiroga, G.; Erice, G.; Aroca, R.; Delgado-Huertas, A.; Ruiz-Lozano, J.M. Elucidating the Possible Involvement of Maize Aquaporins and Arbuscular Mycorrhizal Symbiosis in the Plant Ammonium and Urea Transport under Drought Stress Conditions. Plants 2020, 9, 148. [Google Scholar] [CrossRef]
- Li, T.; Hu, Y.; Hao, Z.; Li, H.; Chen, B. Aquaporin Genes GintAQPF1 and GintAQPF2 from Glomus intraradices Contribute to Plant Drought Tolerance. Plant Signal. Behav. 2013, 8, e24030. [Google Scholar] [CrossRef]
- Gong, M.; Tang, M.; Chen, H.; Zhang, Q.; Feng, X. Effects of Two Glomus Species on the Growth and Physiological Performance of Sophora davidii Seedlings under Water Stress. New For. 2013, 44, 399–408. [Google Scholar] [CrossRef]
- Wu, H.; Xiang, W.; Chen, L.; Ouyang, S.; Xiao, W.; Li, S.; Forrester, D.I.; Lei, P.; Zeng, Y.; Deng, X.; et al. Soil Phosphorus Bioavailability and Recycling Increased with Stand Age in Chinese Fir Plantations. Ecosystems 2020, 23, 973–988. [Google Scholar] [CrossRef]
- Wahid, F.; Sharif, M.; Fahad, S.; Ali, A.; Adnan, M.; Rafiullah; Saud, S.; Danish, S.; Arif Ali, M.; Ahmed, N.; et al. Mycorrhiza and Phosphate Solubilizing Bacteria: Potential Bioagents for Sustainable Phosphorus Management in Agriculture. Phyton 2022, 91, 257–278. [Google Scholar] [CrossRef]
- Alonso, L.M.; Kleiner, D. Spores of the Mycorrhizal Fungus Glomus mosseae Host Yeasts That Solubilize Phosphate and Accumulate Polyphosphates. Mycorrhiza 2008, 18, 197–204. [Google Scholar] [CrossRef]
- Shi, Q.; Pang, J.; Yong, J.W.H.; Bai, C.; Pereira, C.G.; Song, Q.; Wu, D.; Dong, Q.; Cheng, X.; Wang, F.; et al. Phosphorus-Fertilisation Has Differential Effects on Leaf Growth and Photosynthetic Capacity of Arachis hypogaea L. Plant Soil 2020, 447, 99–116. [Google Scholar] [CrossRef]
- Salamatullah, A.M.; Soufan, W.; Nafidi, H.; Ouahmane, L. Mycorrhizal Fungi Inoculation Improves Capparis spinosa’s Yield, Nutrient Uptake and Photosynthetic Efficiency under Water Deficit. Agronomy 2022, 12, 149. [Google Scholar] [CrossRef]
- Hammer, E.C.; Rillig, M.C. The Influence of Different Stresses on Glomalin Levels in an Arbuscular Mycorrhizal Fungus-Salinity Increases Glomalin Content. PLoS ONE 2011, 6, 28426. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Ma, S.; Liu, J.; Qin, S.; Liu, X.; Li, T.; Liao, Y.; Shi, Y.; Zhang, J. Organic Materials and AMF Addition Promote Growth of Taxodium ‘Zhongshanshan’ by Improving Soil Structure. Forests 2023, 14, 731. [Google Scholar] [CrossRef]
- Channavar, V.R.; Hussain, K.N.J.; Vyas, R.D.V.; Malappannavar, N.; Radder, V.S.; Jagadeesh, B.R. The Hidden Powers of Glomalin: Insights into Soil Health and Functionality. Arch. Curr. Res. Int. 2024, 24, 469–479. [Google Scholar] [CrossRef]
- Serrano, R.; Rodriguez-navarro, A. Ion Homeostasis during Salt Stress in Plants. Curr. Opin. Cell Biol. 2001, 13, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Samiappan, S.C.; Mahalakshmi, P.; Pandiyan, R. Impact of Mycorrhizal Soil Fertility Proteins and Arbuscular Mycorrhizal Application to Combat Drought Stress in Maize Plants. J. Plant Biochem. Biotechnol. 2021, 30, 906–917. [Google Scholar] [CrossRef]
- Prasad, K.; Khare, A.; Rawat, P. Glomalin Arbuscular Mycorrhizal Fungal Reproduction, Lifestyle and Dynamic Role in Global Sustainable Agriculture for Future Generation. In Fungal Reproduction and Growth; IntechOpen: Boston, MA, USA, 2022; pp. 1–22. [Google Scholar]
- Gao, W.Q.; Wang, P.; Wu, Q.S. Functions and Application of Glomalin-Related Soil Proteins: A Review. Sains Malays. 2019, 48, 111–119. [Google Scholar] [CrossRef]
- Nawaz, A.; Qamar, Z.U.; Marghoob, M.U.; Imtiaz, M.; Imran, A.; Mubeen, F. Contribution of Potassium Solubilizing Bacteria in Improved Potassium Assimilation and Cytosolic K+/Na+ Ratio in Rice (Oryza sativa L.) under Saline-Sodic Conditions. Front. Microbiol. 2023, 14, 1196024. [Google Scholar] [CrossRef]
- Evelin, H.; Giri, B.; Kapoor, R. Contribution of Glomus intraradices Inoculation to Nutrient Acquisition and Mitigation of Ionic Imbalance in NaCl-Stressed Trigonella foenum-graecum. Mycorrhiza 2012, 22, 203–217. [Google Scholar] [CrossRef] [PubMed]
- Giri, B.; Kapoor, R.; Mukerji, K.G. Improved Tolerance of Acacia nilotica to Salt Stress by Arbuscular Mycorrhiza, Glomus fasciculatum May Be Partly Related to Elevated K/Na Ratios in Root and Shoot Tissues. Microb. Ecol. 2007, 54, 753–760. [Google Scholar] [CrossRef]
- Karimi, R.; Noori, A. Streptomyces rimosus Rhizobacteria and Glomus mosseae Mycorrhizal Fungus Inoculation Alleviate Salinity Stress in Grapevine through Morphophysiological Changes and Nutritional Balance. Sci. Hortic. 2022, 305, 111433. [Google Scholar] [CrossRef]
- Zou, Y.; Wu, Q. Sodium Chloride Stress Induced Changes in Leaf Osmotic Adjustment of Trifoliate Orange (Poncirus trifoliata) Seedlings Inoculated with Mycorrhizal Fungi. Not. Bot. Horti Agrobot. Cluj-Napoca 2011, 39, 64–69. [Google Scholar] [CrossRef]
- Ouhaddou, R.; Anli, M.; Ech-Chatir, L.; Ben-Laouane, R.; Boutasknit, A.; Benaffari, W.; Meddich, A. Boosting Agricultural Productivity in the Mediterranean: Harnessing Natural Biostimulants to Alleviate Abiotic Constraints. In Agricultural Research Updates; Nova Science Publishers: Hauppauge, NY, USA, 2023; Volume 46, pp. 181–216. ISBN 9798891133785. [Google Scholar]
- Sharma, V.; Sharma, D.P.; Salwan, R. Surviving the Stress: Understanding the Molecular Basis of Plant Adaptations and Uncovering the Role of Mycorrhizal Association in Plant Abiotic Stresses. Microb. Pathog. 2024, 193, 106772. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, S.; Wani, K.I.; Naeem, M.; Khan, M.M.A.; Aftab, T. Cellular Responses, Osmotic Adjustments, and Role of Osmolytes in Providing Salt Stress Resilience in Higher Plants: Polyamines and Nitric Oxide Crosstalk. J. Plant Growth Regul. 2023, 42, 539–553. [Google Scholar] [CrossRef]
- Ye, L.; Zhao, X.; Bao, E.; Cao, K.; Zou, Z. Effects of Arbuscular Mycorrhizal Fungi on Watermelon Growth, Elemental Uptake, Antioxidant, and Photosystem Ii Activities and Stress-Response Gene Expressions under Salinity-Alkalinity Stresses. Front. Plant Sci. 2019, 10, 863. [Google Scholar] [CrossRef]
- Evelin, H.; Kapoor, R. Arbuscular Mycorrhizal Symbiosis Modulates Antioxidant Response in Salt-Stressed Trigonella foenum-graecum Plants. Mycorrhiza 2014, 24, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Kohler, J.; Hernández, J.A.; Caravaca, F.; Roldán, A. Induction of Antioxidant Enzymes Is Involved in the Greater Effectiveness of a PGPR versus AM Fungi with Respect to Increasing the Tolerance of Lettuce to Severe Salt Stress. Environ. Exp. Bot. 2009, 65, 245–252. [Google Scholar] [CrossRef]
- Ouhaddou, R.; Ben-Laouane, R.; Lahlali, R.; Anli, M.; Ikan, C.; Boutasknit, A.; Slimani, A.; Oufdou, K.; Baslam, M.; Ait Barka, E.; et al. Application of Indigenous Rhizospheric Microorganisms and Local Compost as Enhancers of Lettuce Growth, Development, and Salt Stress Tolerance. Microorganisms 2022, 10, 1625. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, S.R.Z.; Seifi, E.; Varasteh, F.; Akbarpour, V. Mitigating Salinity Stress in Pomegranate: Effects of Pseudomonas fluorescens and Glomus mosseae on Stress Responses of Red Angel and Wonderful Cultivars. Sci. Hortic. 2024, 330, 113036. [Google Scholar] [CrossRef]
- Yilmaz, A.; Yildirim, E.; Yilmaz, H.; Soydemir, H.E.; Güler, E.; Ciftci, V.; Yaman, M. Use of Arbuscular Mycorrhizal Fungi for Boosting Antioxidant Enzyme Metabolism and Mitigating Saline Stress in Sweet Basil (Ocimum basilicum L.). Sustainability 2023, 15, 5982. [Google Scholar] [CrossRef]
- Huang, S.; Gill, S.; Ramzan, M.; Ahmad, M.Z.; Danish, S.; Huang, P.; Al Obaid, S.; Alharbi, S.A. Uncovering the Impact of AM Fungi on Wheat Nutrient Uptake, Ion Homeostasis, Oxidative Stress, and Antioxidant Defense under Salinity Stress. Sci. Rep. 2023, 13, 8249. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Shi, F.; Zheng, X.; Pan, H.; Wen, Y.; Song, F. Effects of AMF Compound Inoculants on Growth, Ion Homeostasis, and Salt Tolerance-Related Gene Expression in Oryza sativa L. Under Salt Treatments. Rice 2023, 16, 18. [Google Scholar] [CrossRef]
- Garg, N.; Baher, N. Role of Arbuscular Mycorrhizal Symbiosis in Proline Biosynthesis and Metabolism of Cicer arietinum L. (Chickpea) Genotypes Under Salt Stress. J. Plant Growth Regul. 2013, 32, 767–778. [Google Scholar] [CrossRef]
- Altuntas, O.; Dasgan, H.Y.; Akhoundnejad, Y.; Nas, Y. Unlocking the Potential of Pepper Plants under Salt Stress: Mycorrhizal Effects on Physiological Parameters Related to Plant Growth and Gas Exchange across Tolerant and Sensitive Genotypes. Plants 2024, 13, 1380. [Google Scholar] [CrossRef] [PubMed]
- Léopold, I.; Diatta, D.; Kane, A.; Agbangba, C.E.; Sagna, M. Inoculation with Arbuscular Mycorrhizal Fungi Improves Seedlings Growth of Two Sahelian Date Palm Cultivars (Phoenix dactylifera L., Cv. Nakhla Hamra and Cv. Tijib) under Salinity Stresses. Adv. Biosci. Biotechnol. 2014, 5, 64–72. [Google Scholar] [CrossRef]
- Estrada, B.; Barea, J.M.; Aroca, R.; Ruiz-Lozano, J.M. A Native Glomus intraradices Strain from a Mediterranean Saline Area Exhibits Salt Tolerance and Enhanced Symbiotic Efficiency with Maize Plants under Salt Stress Conditions. Plant Soil 2013, 366, 333–349. [Google Scholar] [CrossRef]
- Ikan, C.; Ben-Laouane, R.; Ouhaddou, R.; Ghoulam, C.; Meddich, A. Co-Inoculation of Arbuscular Mycorrhizal Fungi and Plant Growth-Promoting Rhizobacteria Can Mitigate the Effects of Drought in Wheat Plants (Triticum durum). Plant Biosyst. 2023, 157, 907–919. [Google Scholar] [CrossRef]
- Akensous, F.; Anli, M.; Meddich, A. Biostimulants as Innovative Tools to Boost Date Palm (Phoenix dactylifera L.) Performance under Drought, Salinity, and Heavy Metal(Oid)s’ Stresses: A Concise Review. Sustainability 2022, 14, 15484. [Google Scholar] [CrossRef]
- Ghadbane, M.; Medjekal, S.; Benderradji, L.; Belhadj, H.; Daoud, H. Assessment of Arbuscular Mycorrhizal Fungi Status and Rhizobium on Date Palm (Phoenix dactylifera L.) Cultivated in a Pb Contaminated Soil. In Recent Advances in Environmental Science from the Euro-Mediterranean and Surrounding Regions, 2nd ed.; Environmental Science and Engineering; Springer: Cham, Switzerland, 2019; pp. 703–707. [Google Scholar] [CrossRef]
- Begum, N.; Wang, L.; Ahmad, H.; Akhtar, K.; Roy, R.; Khan, M.I.; Zhao, T. Co-Inoculation of Arbuscular Mycorrhizal Fungi and the Plant Growth-Promoting Rhizobacteria Improve Growth and Photosynthesis in Tobacco Under Drought Stress by Up-Regulating Antioxidant and Mineral Nutrition Metabolism. Microb. Ecol. 2022, 83, 971–988. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Soni, S.K.; Bajpai, A. Cooperative Interaction of Glomus intraradices with Plant Growth-Promoting Rhizobacteria Promotes Plant Development and Essential Oil Yield of Pogostemon Cablin and Reduces Disease Occurrence under Organic Field Conditions. Australas. Plant Pathol. 2023, 52, 595–607. [Google Scholar] [CrossRef]
- Li, W.; Li, W.; Xing, L.; Guo, S. Effect of Arbuscular Mycorrhizal Fungi (AMF) and Plant Growth-Promoting Rhizobacteria (PGPR) on Microorganism of Phenanthrene and Pyrene Contaminated Soils. Int. J. Phytoremediat. 2022, 25, 240–251. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.W.; Tong, C.L.; Sun, M.F. Effects and Molecular Mechanism of Mycorrhiza on the Growth, Nutrient Absorption, Quality of Fresh Leaves, and Antioxidant System of Tea Seedlings Suffering from Salt Stress. Agronomy 2022, 12, 1163. [Google Scholar] [CrossRef]
- Postgraduados, C.; Montecillo, C.; Postgraduados, C.; Montecillo, C.; Texcoco, M.; Postgraduados, C.; Hidalgo, S.; Cotaxtla-inifap, C.E. Quality of Floral Stems of Lisianthus (Eustoma grandiflorum Raf.) Inoculated with Bacillus subtilis and Glomus intraradices. Ornam. Hortic. 2022, 28, 414–422. [Google Scholar]
- Sharma, S.; Compant, S.; Ballhausen, M.B.; Ruppel, S.; Franken, P. The Interaction between Rhizoglomus irregulare and Hyphae Attached Phosphate Solubilizing Bacteria Increases Plant Biomass of Solanum lycopersicum. Microbiol. Res. 2020, 240, 126556. [Google Scholar] [CrossRef]
- Berger, F.; Gutjahr, C. Factors Affecting Plant Responsiveness to Arbuscular Mycorrhiza. Curr. Opin. Plant Biol. 2021, 59, 101994. [Google Scholar] [CrossRef]
- Ikan, C.; Ben-Laouane, R.; Ouhaddou, R.; Anli, M.; Boutasknit, A.; Lahbouki, S.; Benchakour, A.; Jaouad, A.; Bouchdoug, M.; El Moatasime, A.; et al. Interactions between Arbuscular Mycorrhizal Fungus and Indigenous Compost Improve Salt Stress Tolerance in Wheat (Triticum durum). S. Afr. J. Bot. 2023, 158, 417–428. [Google Scholar] [CrossRef]
- Yazici, M.A.; Asif, M.; Tutus, Y.; Ortas, I.; Ozturk, L.; Lambers, H.; Cakmak, I. Reduced Root Mycorrhizal Colonization as Affected by Phosphorus Fertilization Is Responsible for High Cadmium Accumulation in Wheat. Plant Soil 2021, 468, 19–35. [Google Scholar] [CrossRef]
- Watts-Williams, S.J.; Cavagnaro, T.R.; Tyerman, S.D. Variable Effects of Arbuscular Mycorrhizal Fungal Inoculation on Physiological and Molecular Measures of Root and Stomatal Conductance of Diverse Medicago truncatula Accessions. Plant. Cell Environ. 2018, 42, 285–294. [Google Scholar] [CrossRef]
- Koch, A.M.; Antunes, P.M.; Maherali, H.; Hart, M.M.; Klironomos, J.N. Evolutionary Asymmetry in the Arbuscular Mycorrhizal Symbiosis: Conservatism in Fungal Morphology Does Not Predict Host Plant Growth. New Phytol. 2017, 214, 1330–1337. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ouhaddou, R.; Anli, M.; Ben-Laouane, R.; Boutasknit, A.; Baslam, M.; Meddich, A. The Importance of the Glomus Genus as a Potential Candidate for Sustainable Agriculture Under Arid Environments: A Review. Int. J. Plant Biol. 2025, 16, 32. https://doi.org/10.3390/ijpb16010032
Ouhaddou R, Anli M, Ben-Laouane R, Boutasknit A, Baslam M, Meddich A. The Importance of the Glomus Genus as a Potential Candidate for Sustainable Agriculture Under Arid Environments: A Review. International Journal of Plant Biology. 2025; 16(1):32. https://doi.org/10.3390/ijpb16010032
Chicago/Turabian StyleOuhaddou, Redouane, Mohamed Anli, Raja Ben-Laouane, Abderrahim Boutasknit, Marouane Baslam, and Abdelilah Meddich. 2025. "The Importance of the Glomus Genus as a Potential Candidate for Sustainable Agriculture Under Arid Environments: A Review" International Journal of Plant Biology 16, no. 1: 32. https://doi.org/10.3390/ijpb16010032
APA StyleOuhaddou, R., Anli, M., Ben-Laouane, R., Boutasknit, A., Baslam, M., & Meddich, A. (2025). The Importance of the Glomus Genus as a Potential Candidate for Sustainable Agriculture Under Arid Environments: A Review. International Journal of Plant Biology, 16(1), 32. https://doi.org/10.3390/ijpb16010032