
Liriodenine and Its Probable Role as an Osmolyte during Water Stress in Annona lutescens (Annonaceae)
 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Growth Conditions and Treatments
2.2. Net CO2 Assimilation (An), Stomatal Conductance (gs) and Growth Analysis
2.3. Pressure–Volume Curves
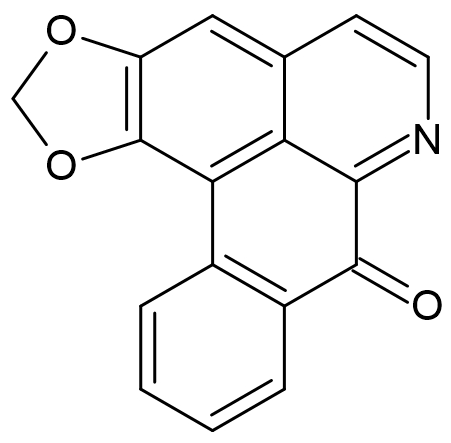
2.4. Liriodenine Content
2.5. Liriodenine Test and HCl Solubility
2.6. Test of pH Regulation by Liriodenine in Malic Acid
2.7. Statistical Analysis
3. Results
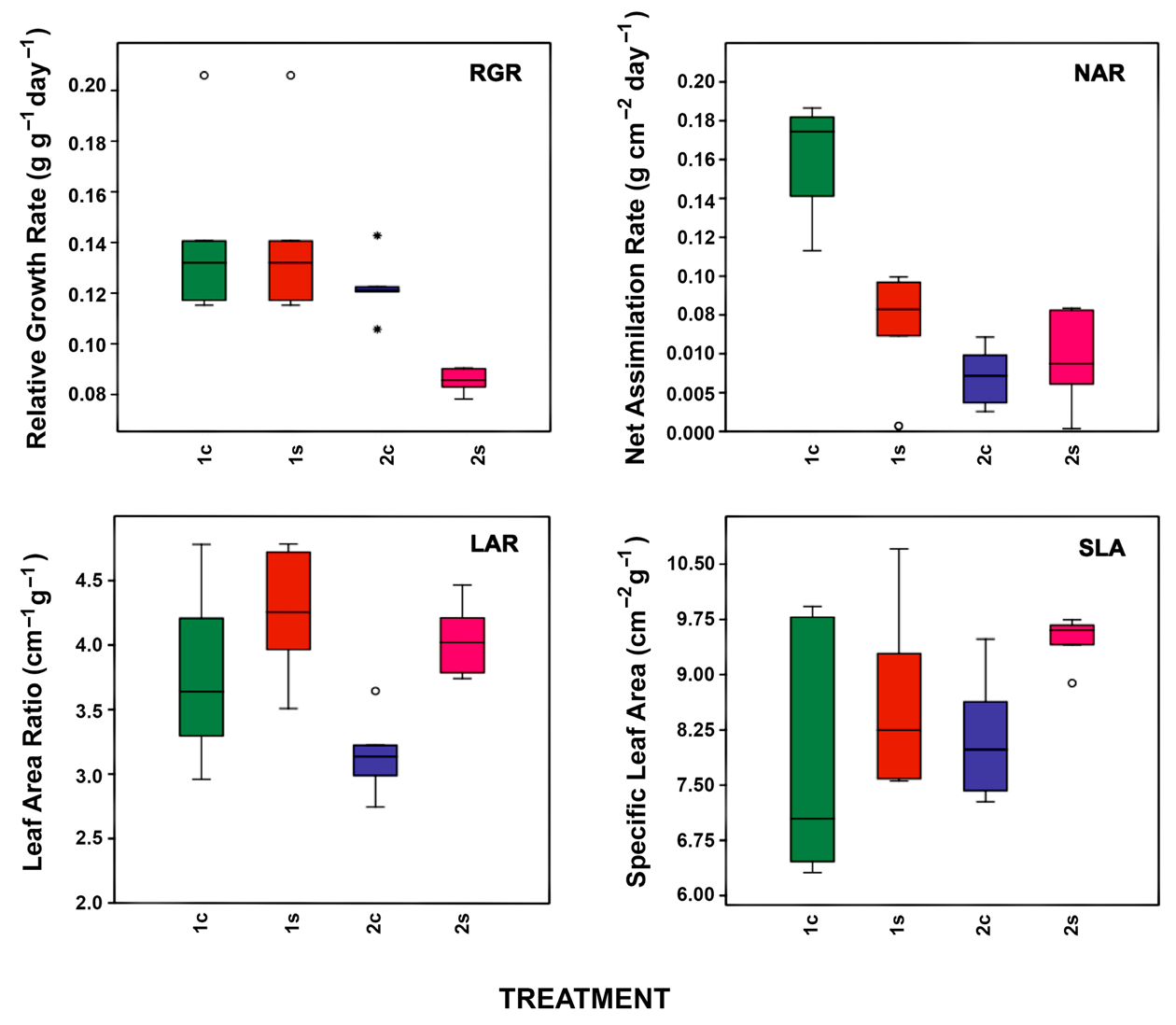
3.1. Net CO2 Assimilation Rate (An), Stomatal Conductance (gs) and Growth Analysis
3.2. Pressure–Volume Curves
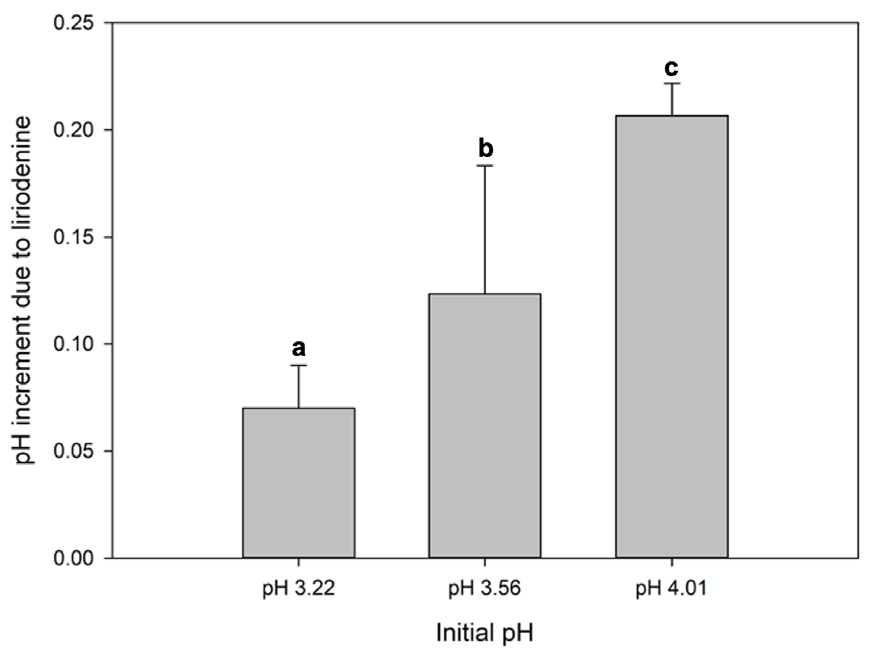
3.3. Liriodenine Content and Tests at Different pH Values
3.4. Non-Metric Multidimensional Scaling (NMDS)
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding plant responses to drought–from genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef] [PubMed]
- Lambers, H.; Chapin, F.S., III; Pons, T.L. Plant Physiological Ecology; Springer: New York, NY, USA, 2008; 571p. [Google Scholar]
- Mahajan, M.; Kuiry, R.; Pal, P.K. Understanding the consequence of environmental stress for accumulation of secondary metabolites in medicinal and aromatic plants. J. Appl. Res. Med. 2020, 18, 100255. [Google Scholar] [CrossRef]
- Oh, M.; Trick, H.N.; Rajasekar, C.B. Secondary metabolism and antioxidants are involved in environmental adaptation and stress tolerance in lettuce. J. Plant Physiol. 2009, 166, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Selmar, D.; Kleinwächter, M. Influencing the product quality by deliberately applying drought stress during the cultivation of medicinal plants. Ind. Crops Prod. 2013, 42, 558–566. [Google Scholar] [CrossRef]
- Skoneczny, D.; Weston, P.A.; Weston, L.A. Metabolomics and metabolic profiling: Investigation of dynamic plant-environment interactions at the functional level. In Advances in Plant Ecophysiology Techniques; Sánchez-Moreiras, A.M., Reigosa, M.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; pp. 323–345. [Google Scholar] [CrossRef]
- Kirakosyan, A.; Kaufman, P.; Warber, S.; Zick, S.; Aaronson, K.; Bolling, S. Applied environmental stresses to enhance the levels of polyphenolics in leaves of hawthorn plants. Physiol. Plant. 2004, 121, 182–186. [Google Scholar] [CrossRef] [PubMed]
- De Abreu, I.N.; Mazzafera, P. Effect of water temperature stress on the content of active constituents of Hypericum brasiliense Choisy. Plant Physiol. Biochem. 2005, 43, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Nowak, M.; Manderscheid, R.; Weigel, H.J.; Kleinwächter, M.; Selmar, D. Drought stress increases the accumulation of monoterpenes in sage (Salvia officinalis), an effect that is compensated by elevated carbon dioxide concentration. J. Appl. Bot. Food Qual. 2010, 83, 133–136. [Google Scholar]
- Manukyan, A. Effect of growing factors on productivity and quality of lemon catmint, lemon balm and sage under soilless greenhouse production: I. Drought stress. Med. Aromat. Plant Sci. Biotechnol. 2011, 5, 119–125. [Google Scholar]
- Zhu, Z.; Liang, Z.; Han, R.; Wang, X. Impact of fertilization a drought response in the medicinal herb Bupleurum chinense DC: Growth and saikosaponin production. Ind. Crops Prod. 2009, 29, 629–663. [Google Scholar] [CrossRef]
- Nogués, S.; Allen, D.J.; Morison, J.I.L.; Baker, N.R. Ultraviolet-B radiation effects on water relations, leaf development, and photosynthesis in droughted pea plants. Plant Physiol. 1998, 117, 173–181. [Google Scholar] [CrossRef]
- Cakir, R.; Cebi, U. The effect of irrigation scheduling and water stress on the maturity and chemical composition on Virginia tobacco leaf. Field Crops Res. 2010, 119, 269–276. [Google Scholar] [CrossRef]
- Kirk, H.; Vrieling, K.; Van Der Meijden, E.; Klinkhamer, P.G.L. Species by environment interaction after pyrrolizidine alkaloid expression in Senecio jacobaea, Senecio aquaticus, and their hybrid. J. Chem. Ecol. 2010, 36, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Castro-Moreno, M.; Tinoco-Ojangurén, C.L.; Cruz-Ortega, M.D.R.; González-Esquinca, A.R. Influence of seasonal variation on the phenology and liriodenine content of Annona lutescens (Annonaceae). J. Plant Res. 2013, 126, 529–537. [Google Scholar] [CrossRef]
- Szabó, B.; Tyihák, E.; Szabó, L.G.Y.; Botz, L. Mycotoxin and drought stress induced chance of alkaloids content of Papaver somniferum plantlets. Acta Bot. Hung. 2003, 45, 409–417. [Google Scholar] [CrossRef]
- Briske, D.D.; Camp, B.J. Water stress increases alkaloid concentrations in threadleaf groundsel (Senecio longilobus). Weed Sci. 1982, 30, 106–108. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Sankar, B.; Murali, P.V.; Gomathinayagam, M.; Lakshmanan, G.M.A.; Panneerselvam, R. Water deficit stress effects on reactive oxygen metabolism in Catharanthus roseus; impacts on ajmalicine accumulation. Colloids Surf. B. 2008, 62, 105–111. [Google Scholar] [CrossRef]
- Ababaf, M.; Omidi, H.; Bakhshandeh, A. Changes in antioxidant activities and alkaloid amount of Catharanthus roseus in response to plant growth regulators under drought condition. Ind. Crops Prod. 2021, 167, 11. [Google Scholar] [CrossRef]
- De-la -Cruz-Chacón, I.; Riley-Saldaña, C.A.; Arrollo-Gómez, S.; Sancristóbal-Domínguez, T.J.; Castro-Moreno, M.; González-Esquinca, A.R. Spatio-temporal variation of alkaloids in Annona purpurea and the associated influence on their antifungal activity. Chem. Biodivers. 2018, 16, e1800284. [Google Scholar] [CrossRef] [PubMed]
- Honório, A.B.M.; De-la-Cruz-Chacón, I.; Martínez-Vázquez, M.; da Silva, M.R.; Campos, F.G.; Martin, B.C.; da Silva, G.C.; Fernandes Boaro, C.S.; Ferreira, G. Impact of drought and flooding on alkaloid production in Annona crassiflora Mart. Horticulturae 2021, 7, 414. [Google Scholar] [CrossRef]
- De-La-Cruz-Chacón, I.; González-Esquinca, A.R. Liriodenine alkaloid in Annona diversifolia during early development. Nat. Prod. Res. 2012, 26, 42–49. [Google Scholar] [CrossRef]
- Cohen, J.; Langenthal, W.; Taylor, W. Notes- The alkaloids of Liriodendron tulipifera L. The structure and synthesis of the unnamed yellow alkaloid and the isolation of D-glucina. J. Org. Chem. 1961, 26, 4143–4144. [Google Scholar] [CrossRef]
- Taylor, W.I. The structure and synthesis of liriodenine, a new type of isoquinoline alkaloid. Tetrahedron 1961, 14, 42–45. [Google Scholar] [CrossRef]
- Ríos, J.L.; Máñez, S.; Giner, R.M.; Recio, M.C. Biological aspects of aporphinoid alkaloids. In The Alkaloids: Chemistry and Biology; Cordell, G.A., Ed.; Academic Press: Cambridge, MA, USA, 1999; Volume 53, pp. 57–117. [Google Scholar]
- González-Esquinca, A.R.; De-La-Cruz-Chacón, I.; Castro-Moreno, M.; Riley-Saldaña, C.A. Phenological strategies of Annona species from the tropical deciduous forest of Chiapas, México. Bot. Sci. 2016, 94, 531–541. [Google Scholar] [CrossRef]
- Facchini, P.J.; De Luca, V. Phloem-specific expression of tyrosine/dopa decarboxylase genes and the biosynthesis of isoquinoline alkaloids in Opium poppy. Plant Cell 1995, 7, 1811–1821. [Google Scholar] [CrossRef]
- Roberts, M.F.; Wink, M. Alkaloids: Biochemistry, Ecology and Medicinal Application; Plenum Press: New York, NY, USA, 1998. [Google Scholar] [CrossRef]
- Hunt, R.; Cornelissen, J.H.C. Components of relative growth rate and their interrelations in 59 temperate plant species. New Phytol. 1997, 135, 395–417. [Google Scholar] [CrossRef]
- Tyree, M.T.; Hammel, H.T. The measurement of the turgor pressure and the water relations of plant by the pressure-bomb technique. J. Exp. Bot. 1972, 23, 267–282. [Google Scholar] [CrossRef]
- Wilson, J.R.; Ludlow, M.M.; Fisher, M.J.; Schulze, E. Adaptation to water stress of the leaf water relation of four tropical forage species. Aus. J. Plant Physiol. 1980, 7, 207–220. [Google Scholar] [CrossRef]
- González-Esquinca, A.R.; De-La-Cruz-Chacón, I.; Castro-Moreno, M.; Orozco-Castillo, J.A.; Riley-Saldaña, C.A. Alkaloids and acetogenins in Annonaceae development: Biological considerations. Rev. Bras. Frutic. 2014, 36, 1–16. [Google Scholar] [CrossRef]
- Shriner, R.L.; Fuson, R.C.; Curtin, D.Y. Identificación Sistemática de Compuestos Orgánicos; Cap. 6. Comportamiento de la Solubilidad. ed. Limusa: Ciudad de México, Mexico, 1977. [Google Scholar]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics software package for education and data analysis. Palaeontol. Electrónica 2001, 4, 9. [Google Scholar]
- Jaleel, C.A.; Manivannan, P.; Kishorekumar, A.; Sankar, B.; Gopi, R.; Somasundaram, R.; Panneer, S. Alterations in osmoregulation, antioxidant enzymes and indole alkaloid levels in Catharanthus roseus exposed to water deficit. Colloids Surf. B. 2007, 59, 150–157. [Google Scholar] [CrossRef]
- Liu, J.; Gao, F.; Ren, J.; Lu, X.; Ren, G.; Wang, R. A novel AP2/ERF transcription factor CR1 regulates the accumulation of vindoline and serpentine in Catharanthus roseus. Front. Plant Sci. 2017, 8, 2082. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Meng, Q.; Duan, X.; Zhang, Z.; Li, D. Effects of PEG-induced drought stress on regulation of indole alkaloid biosynthesis in Catharanthus roseus. J. Plant Interact. 2017, 12, 87–91. [Google Scholar] [CrossRef]
- Mall, M.; Verma, R.K.; Gupta, M.M.; Shasany, A.K.; Khanuja, S.P.S.; Shukla, A.K. Influence of seasonal and ontogenic parameters on the pattern of key terpenoid indole alkaloids biosynthesized in the leaves of Catharanthus roseus. S. Afr. J. Bot. 2019, 123, 98–2014. [Google Scholar] [CrossRef]
- Liang, J.; Quan, M.; Chaowen, S.; He, A.; Xiang, X.; Feng, C. Effects of drought stress on growth, photosynthesis alkaloid accumulation of Lycoris Aurea. Pak. J. Bot. 2020, 52, 1137–1442. [Google Scholar] [CrossRef] [PubMed]
- Alinejad, S.; Sarabi, V.; Sadeghi, A.R.; Hashempour, H. Variation in physiological traits, yield and secondary metabolites of jimsonweed (Datura stramonium L.) under different irrigation regimes and nutrition systems. Ind. Crops Prod. 2020, 143, 8. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Y.; Guo, Q.; Zhu, G.; Wang, C.; Liu, Z. Effects of drought stress on the growth, physiology and secondary metabolite production in Pinellia ternate Thunb. Pak. J. Bot. 2021, 53, 833–840. [Google Scholar] [CrossRef] [PubMed]
- Maréchaux, I. Individual-Based Modeling of Tropical Forests: Role of Biodiversity and Responses to Drought. Ph.D. Thesis, Laboratoire Evolution et Diversité Biologique, Université de Toulouse 3 Paul Sabatier, Toulouse, France, 2016. [Google Scholar]
- Thilagavathy, A.; Naik, K.; Devaraj, V.R. microRNAs: Key Modulators of Drought stress responses in Plants. In Metabolic Adaptations in Plants during Abiotic Stress; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]
- Barrios-Gómez, E.; López-Castañeda, C.; Kohashi-Shibata, J. Relaciones hídricas y temperaturas altas en frijol del tipo “Flor de mayo”. Agron. Costarric. 2011, 35, 131–145. [Google Scholar] [CrossRef]
- Basu, S.; Ramegowda, V.; Kumar, A.; Pereira, A. Plant adaptation to drought stress. F1000Research 2016, 5, 1554. [Google Scholar] [CrossRef]
- Zivcak, M.; Brestic, M.; Sytar, O. Osmotic Adjustment and Plant Adaptation to drought Stress. In Drought Stress Tolerance in Plants, Volume 1; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar] [CrossRef]
- Roychoudhury, A.; Chakraborty, M. Biochemical and Molecular basis of varietal difference in plant salt tolerance. Annu. Rev. Res. Biol. 2013, 3, 422–454. [Google Scholar]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Roychoudhury, A.; Banerjee, A. Endogenous glycine betaine accumulation mediates abiotic stress tolerance in plants. Trop. Plant Res. 2016, 3, 105–111. [Google Scholar]
- Gomes, F.P.; Oliva, M.A.; Mielke, M.S.; Almeida, A.A.F.; Aquino, L.A. Osmotic adjustment, proline accumulation and cell membrane stability in leaves of Cocos nucifera submitted to drought stress. Sci. Hortic. 2010, 126, 379–384. [Google Scholar] [CrossRef]
- Nayyar, H. Accumulation of osmolytes and osmotic adjustment in water-stressed wheat (Triticum aestivum) and maize (Zea mays) as affected by calcium and its antagonist. Environ. Exp. Bot. 2003, 50, 253–264. [Google Scholar] [CrossRef]
- Liu, S.; Yan, Z.; Chen, Y.; Zhang, M.; Chen, J.; Han, W. Foliar pH, an emerging plant functional trait: Biogeography and variability across northern China. Glob. Ecol. Biogeogr. 2019, 28, 386–397. [Google Scholar] [CrossRef]
- Luo, Y.; Yan, Z.; Liu, S.; Chen, J.; Li, K.; Mohammat, A.; Han, W. Variation in desert shrub foliar pH in relation to drought and salinity in Xinjiang, China. J. Veg. Sci. 2021, 32, e13031. [Google Scholar] [CrossRef]
- Singh, L.R.; Dar, T.A. Cellular Osmolytes: From Chaperoning Protein Folding to Clinical Perspectives; Springer: Singapore, 2017. [Google Scholar]
- Matile, P. Localization of alkaloids and mechanism of their accumulation in vacuoles of Chelidonium majus laticifers. Nova Acta Leopold. Supplementum 1976, 1976. [Google Scholar]
- Nowak, M.; Selmar, D. Cellular distribution of alkaloids and their translocation via phloem and xylem: The importance of compartment pH. Plant Biol. 2016, 18, 879–882. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Ali, S.; Zandi, P.; Mehmood, A.; Ullah, S.; Ikram, M.; Ismail, M.A.S.; Babar, M.A. Role of sugar, amino acids in improving plant abiotic stress tolerance. Pak. J. Bot. 2020, 52, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Nahar, K.; Hasanuzzaman, M.; Alam, M.M.; Fujita, M. Glutathione-induced drought stress tolerance in mung bean: Coordinated roles of the antioxidant defense and methylglyoxal detoxification systems. AoB Plants 2015, 7, plv069. [Google Scholar] [CrossRef]
- Iqbal, N.; Nazar, R.; Khan, N.A. Osmolytes and Plants Acclimation to Changing Environment: Emerging Omics Technologies; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Duke, J.A. Dr Duke’s Phytochemical and Ethnobotanical Database, Phytochemical Database, USDA-ARSNGRL, Beltsville Agricultural Research Centre, Maryland, USA, 2000. Available online: https://www.vetiver.org/TVN_vetoil03.pdf (accessed on 3 May 2024).
- De Siquiera, J.M.; Bomm, M.D.; Gomes, P.N.F. Activity—Guided isolation of constituents of Unonopsis lindmanii—Annonaceae, based on the brine shrimp lethality bioassay. Química Nova 1998, 21, 557–559. [Google Scholar] [CrossRef]
- Wirasathien, L.; Boonarkart, C.; Pengsuparp, T.; Suttisri, R. Biological activities of alkaloids from Pseuduvaria setosa. Pharm. Biol. 2006, 44, 274–278. [Google Scholar] [CrossRef]
- Hossain, M.S.; Ferdous, A.J.; Hasan, C.M. In vitro antimicrobial activities of alkaloids from the stem bark of Desmos longiflorus (Roxb.). Bangladesh J. Bot. 1993, 22, 37–40. [Google Scholar]
- Rhaman, M.M.; Loppa, S.S.; Sadik, G.; Rashid, H.O.; Islam, R.; Khonkar, P.; Rashid, M.A. Antibacterial and cytotoxic compounds from the bark of Cananga odorata. Fitoterapia 2005, 76, 758–761. [Google Scholar] [CrossRef] [PubMed]
- Waechter, A.I.; Cave´, A.; Hocquemiller, R.; Bories, C.; Muñoz, V.; Fournet, A. Antiprotozoal activity of aporphine alkaloids isolated from Unonopsis buchtienii (Annonaceae). Phytother. Res. 1999, 13, 175–177. [Google Scholar] [CrossRef]
- Hesse, M. Alkaloids: Nature’s Curse or Blessing? Verlag Helvetica Chemica Acta: Zurich, Switzerland; Wiley-VCH: Weinheim, Germany, 2002; p. 414. [Google Scholar]
- Hufford, C.D.; Sharma, A.S.; Oguntimein, B.O. Antibacterial and antifungal activity of liriodenine and related oxoaporphine alkaloids. J. Pharm. Sci. 1980, 69, 1180–1183. [Google Scholar] [CrossRef]
- Woo, S.H.; Sun, N.J.; Cassady, J.M.; Snapka, R.M. Topoisomerase II inhibition by aporphine alkaloids. Biochem. Pharmacol. 1999, 57, 1141–1145. [Google Scholar] [CrossRef]
- De-La-Cruz-Chacón, I.; González-Esquinca, A.R.; Fefer, P.G.; Garcia, L.F. Liriodenine, early antimicrobial defense in Annona diversifolia. Z. Naturforschung C 2011, 66, 377–384. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Water Stress (Days) | RWC (%) | ε (MPa) | π0 (MPa) |
|---|---|---|---|
| 0 | 82.33 ± 13 | 2.79 ± 1.85 | −1.23 ± 0.33 a |
| 15 | 72 ± 19 | 1.29 ± 1.42 | −1.99 ± 0.35 b |
| 25 | 75.16 ± 15 | 3.63 ± 3.13 | −2.54 ± 0.31 c |
| 35 | 72 ± 7.56 | 1.67 ± 0.96 | −2.93 ± 0.27 c |
| p = 0.42 | p = 0.45 | p < 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cisneros-Andrés, A.; Cruz-Ortega, R.; Castro-Moreno, M.; González-Esquinca, A.R. Liriodenine and Its Probable Role as an Osmolyte during Water Stress in Annona lutescens (Annonaceae). Int. J. Plant Biol. 2024, 15, 429-441. https://doi.org/10.3390/ijpb15020033
Cisneros-Andrés A, Cruz-Ortega R, Castro-Moreno M, González-Esquinca AR. Liriodenine and Its Probable Role as an Osmolyte during Water Stress in Annona lutescens (Annonaceae). International Journal of Plant Biology. 2024; 15(2):429-441. https://doi.org/10.3390/ijpb15020033
Chicago/Turabian StyleCisneros-Andrés, Alfredo, Rocío Cruz-Ortega, Marisol Castro-Moreno, and Alma Rosa González-Esquinca. 2024. "Liriodenine and Its Probable Role as an Osmolyte during Water Stress in Annona lutescens (Annonaceae)" International Journal of Plant Biology 15, no. 2: 429-441. https://doi.org/10.3390/ijpb15020033
APA StyleCisneros-Andrés, A., Cruz-Ortega, R., Castro-Moreno, M., & González-Esquinca, A. R. (2024). Liriodenine and Its Probable Role as an Osmolyte during Water Stress in Annona lutescens (Annonaceae). International Journal of Plant Biology, 15(2), 429-441. https://doi.org/10.3390/ijpb15020033