
Hydrogen Peroxide Measurement Can Be Used to Monitor Plant Oxidative Stress Rapidly Using Modified Ferrous Oxidation Xylenol Orange and Titanium Sulfate Assay Correlation

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Leaves Collection
2.2. Preparation of Frozen and Nonfrozen Samples
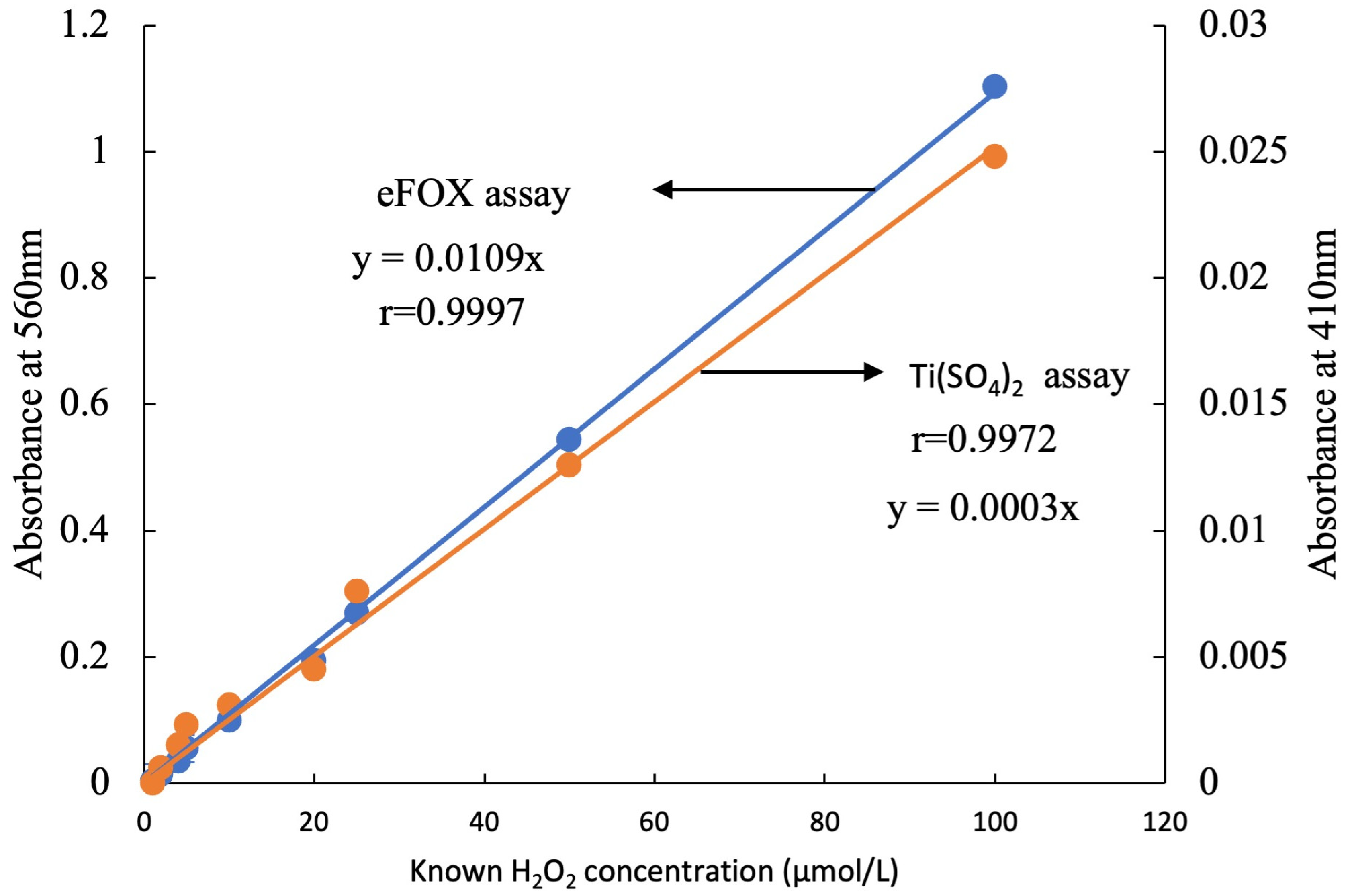
2.3. Preparations of the Standard Curve with the eFOX and Ti(SO4)2
2.4. Quantification of H2O2 Content with eFOX Assay
2.5. Determination of H2O2 Content with Ti(SO4)2 Assay
2.6. Statistical Analyses
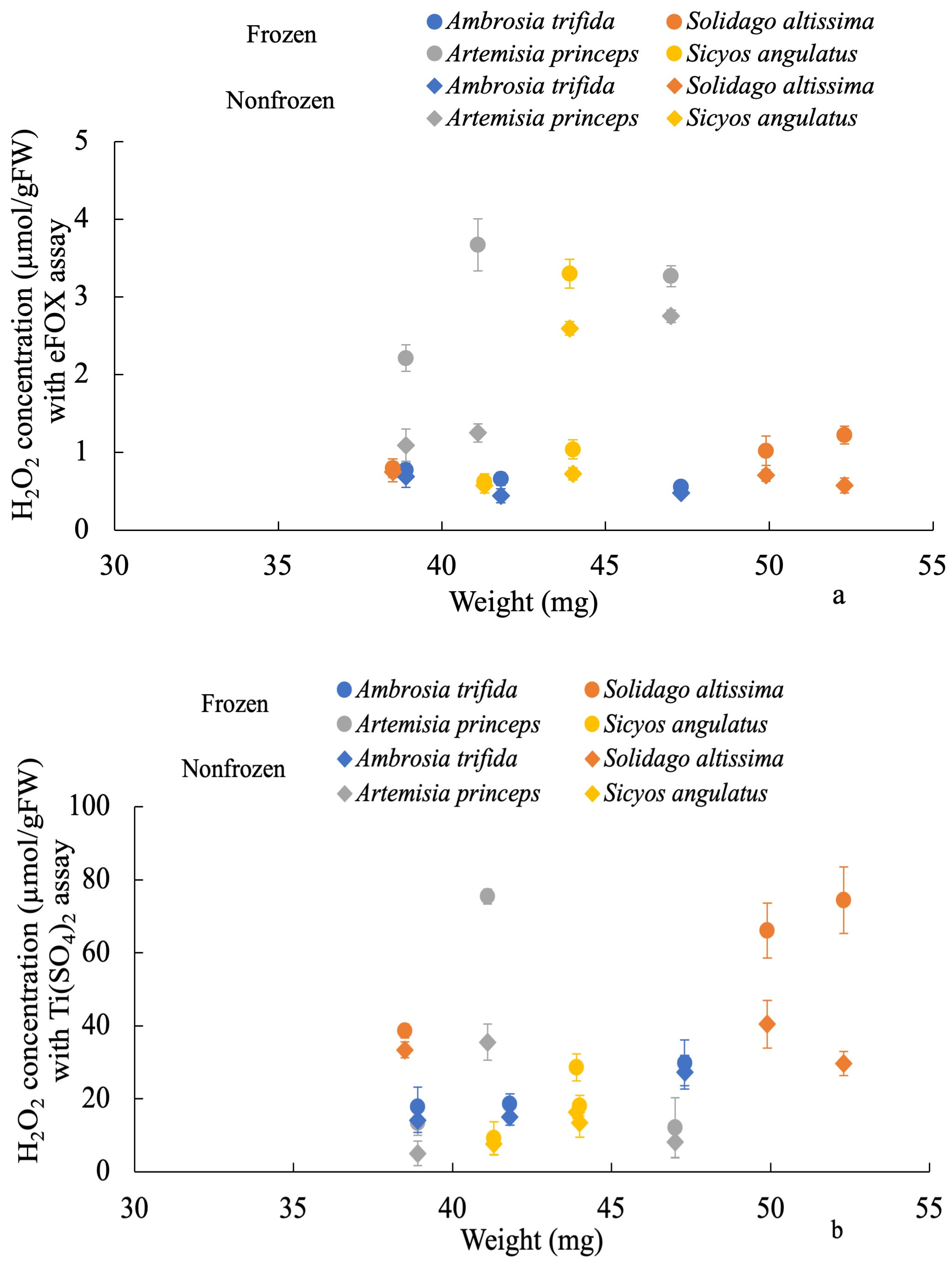
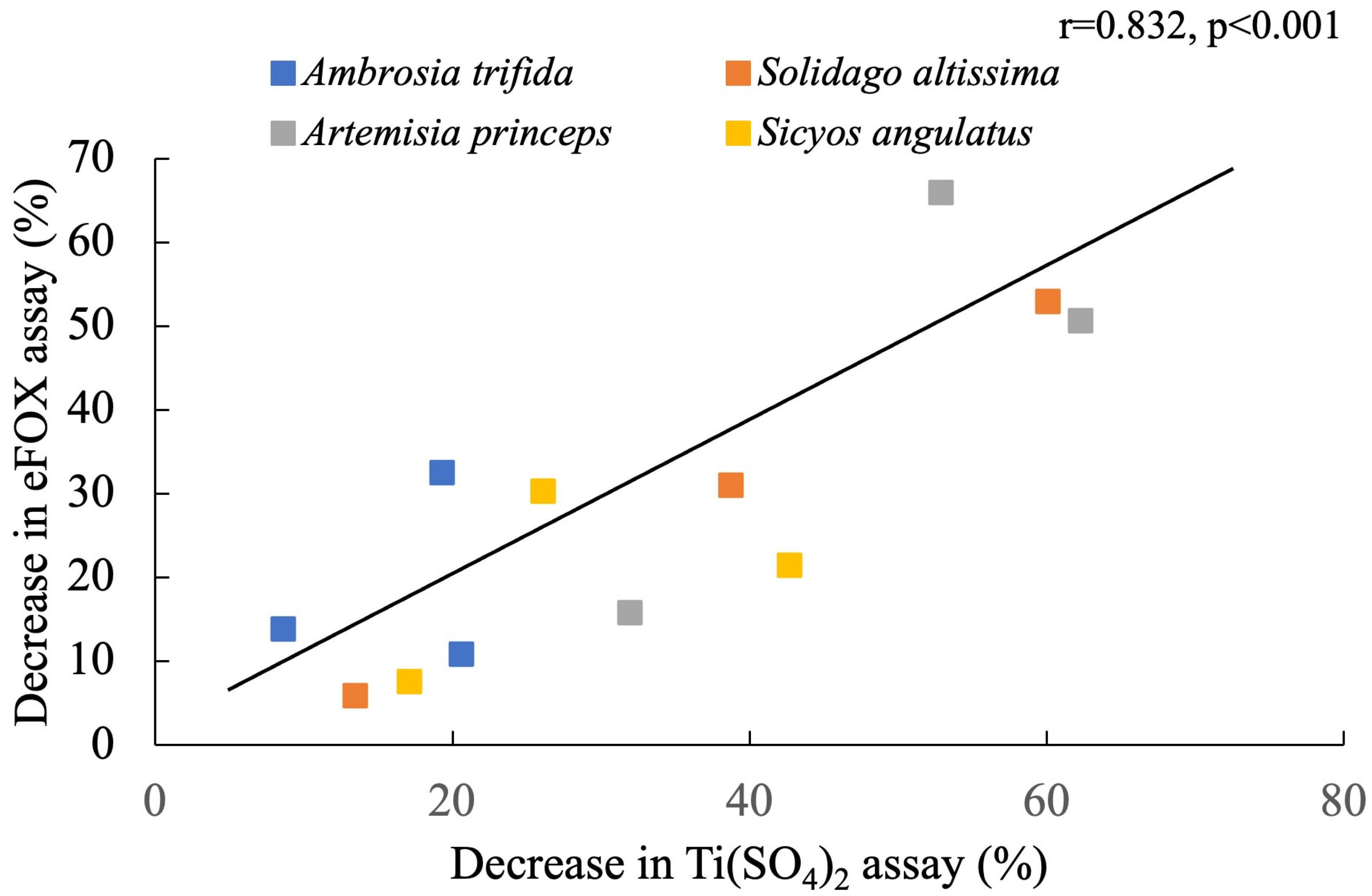
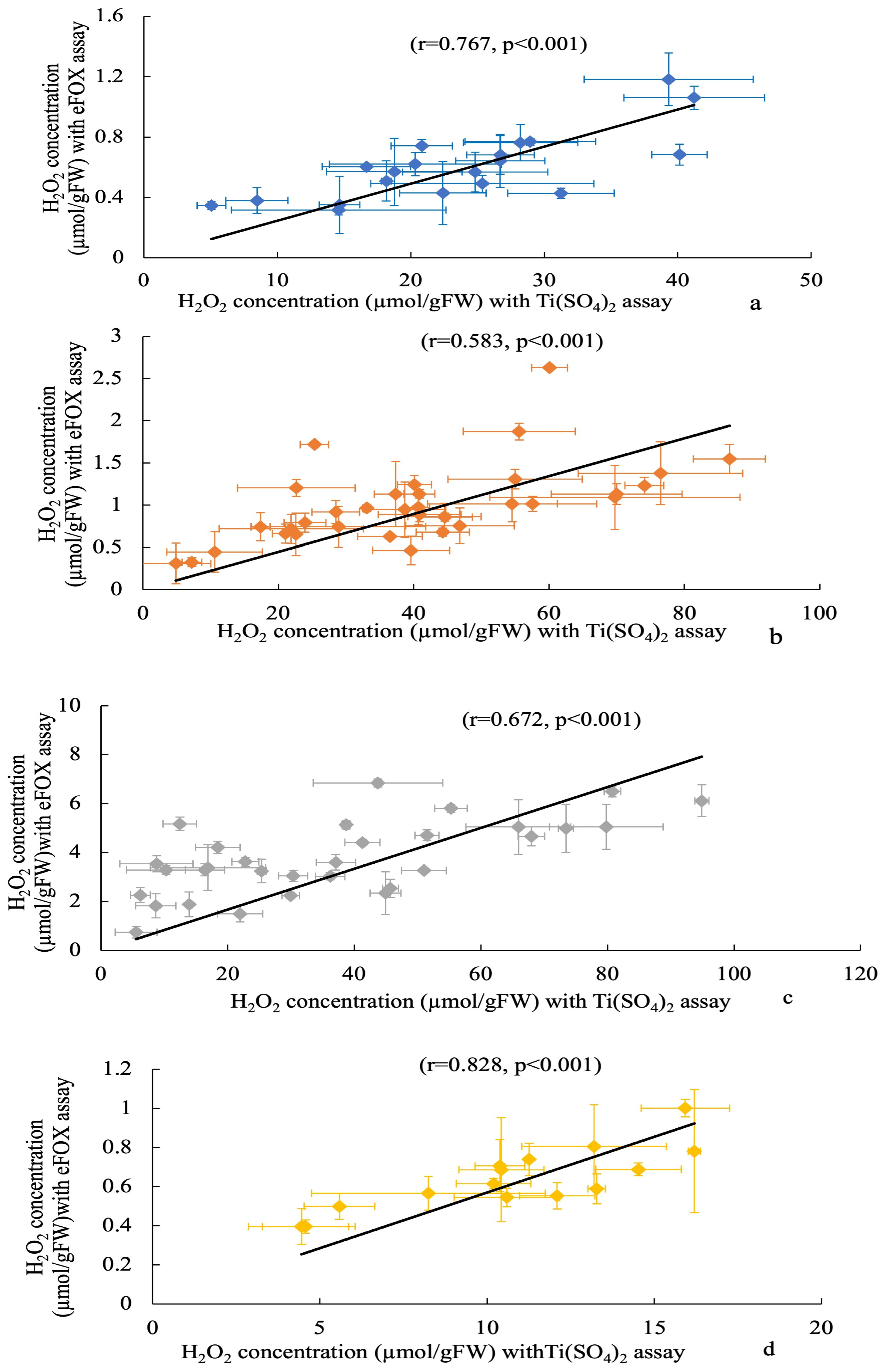
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nallaperuma, B.; Asaeda, T. Long-term changes in riparian forest cover under a dam-induced flow scheme: The accompanying a numerical modelling perspective. J. Ecohydraulics 2019, 4, 106–112. [Google Scholar] [CrossRef]
- Corbacho, C.; Sánchez, J.M.; Costillo, E. Patterns of structural complexity and human disturbance of riparian vegetation in agricultural landscapes of a Mediterranean area. Agric. Ecosyst. Environ. 2003, 95, 495–507. [Google Scholar] [CrossRef]
- Richardson, J.S.; Taylor, E.B.; Schluter, D.; Pearson, M.; Hatfield, T.; Lapointe, N.W.; Cooke, S.J.; Imhof, J.G.; Boisclair, D.; Casselman, J.M.; et al. Do riparian zones qualify as critical habitat for endangered freshwater fishes? Can. J. Fish. Aquat. Sci. 2010, 67, 1197–1204. [Google Scholar] [CrossRef]
- Collier, P.; Gunning, J.W. Why Has Africa Grown Slowly? J. Econ. Perspect. 1999, 13, 3–22. [Google Scholar] [CrossRef]
- Yarrow, M.; Marín, V.H.; Finlayson, M.; Tironi, A.; Delgado, L.E.; Fischer, F. The ecology of Egeria densa Planchón (Liliopsida: Alismatales): A wetland ecosystem engineer? Rev. Chil. De Hist. Nat. 2009, 82, 299–313. [Google Scholar] [CrossRef]
- Asada, K. The Water-Water Cycle in Chloroplasts: Scavenging of Active Oxygens and Dissipation of Excess Photons. Annu. Rev. Biol. 1999, 50, 601–639. [Google Scholar] [CrossRef]
- Neill, S.; Desikan, R.; Hancock, J. Hydrogen peroxide signalling. Curr. Opin. Plant Biol. 2002, 5, 388–395. [Google Scholar] [CrossRef]
- Orozco-Cárdenas, M.L.; Narváez-Vásquez, J.; Ryan, C.A. Hydrogen Peroxide Acts as a Second Messenger for the Induction of Defense Genes in Tomato Plants in Response to Wounding, Systemin, and Methyl Jasmonate. Plant Cell 2001, 13, 179–191. [Google Scholar] [CrossRef]
- Wohlgemuth, H.; Mittelstrass, K.; Kschieschan, S.; Bender, J.; Weigel, H.-J.; Overmyer, K.; Kangasjärvi, J.; Sandermann, H.; Langebartels, C. Activation of an oxidative burst is a general feature of sensitive plants exposed to the air pollutant ozone. Plant Cell Environ. 2002, 25, 717–726. [Google Scholar] [CrossRef]
- Spiro, M.D.; Ridley, B.L.; Eberhard, S.; Kates, K.A.; Mathieu, Y.; O’Neill, M.A.; Mohnen, D.; Guern, J.; Darvill, A.; Albersheim, P. Biological Activity of Reducing-End-Derivatized Oligogalacturonides in Tobacco Tissue Cultures1. Plant Physiol. 1998, 116, 1289–1298. [Google Scholar] [CrossRef]
- Bellincampi, D.; Dipierro, N.; Salvi, G.; Cervone, F.; De Lorenzo, G. Extracellular H2O2 Induced by Oligogalacturonides Is Not Involved in the Inhibition of the Auxin-Regulated rolB Gene Expression in Tobacco Leaf Explants. Plant Physiol. 2000, 122, 1379–1386. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Silva, H.; Klessig, D.F. Active oxygen species in the induction of plant systemic acquired resistance induced by salicylic acid. Science 1993, 262, 1883–1886. [Google Scholar] [CrossRef] [PubMed]
- Gechev, T.S.; Hille, J. Hydrogen peroxide as a signal controlling plant programmed cell death. J. Cell Biol. 2005, 168, 17–20. [Google Scholar] [CrossRef] [PubMed]
- Asaeda, T.; Rahman, M.; Abeynayaka, H.D.L. Hydrogen peroxide can be a plausible biomarker in cyanobacterial bloom treatment. Sci. Rep. 2022, 12, 12. [Google Scholar] [CrossRef]
- Asaeda, T.; Jayasanka, S.M.D.H.; Xia, L.-P.; Barnuevo, A. Application of Hydrogen Peroxide as an Environmental Stress Indicator for Vegetation Management. Engineering 2018, 4, 610–616. [Google Scholar] [CrossRef]
- Asaeda, T.; Rahman, M.; Liping, X.; Schoelynck, J. Hydrogen Peroxide Variation Patterns as Abiotic Stress Responses of Egeria densa. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef]
- Cho, U.-H.; Seo, N.-H. Oxidative stress in Arabidopsis thaliana exposed to cadmium is due to hydrogen peroxide accumulation. Plant Sci. 2005, 168, 113–120. [Google Scholar] [CrossRef]
- Işeri, Ö.D.; Körpe, D.A.; Sahin, F.I.; Haberal, M. Hydrogen peroxide pretreatment of roots enhanced oxidative stress response of tomato under cold stress. Acta Physiol. Plant. 2013, 35, 1905–1913. [Google Scholar] [CrossRef]
- Chaliha, M.; Sultanbawa, Y. Terminalia ferdinandiana, a traditional medicinal plant of Australia, alleviates hydrogen peroxide induced oxidative stress and inflammation, in vitro. J. Complement. Integr. Med. 2019, 17. [Google Scholar] [CrossRef]
- Smirnoff, N.; Arnaud, D. Hydrogen peroxide metabolism and functions in plants. New Phytol. 2018, 221, 1197–1214. [Google Scholar] [CrossRef]
- Satterfield, C.N.; Bonnell, A.H. Interferences in Titanium Sulfate Method for Hydrogen Peroxide. Anal. Chem. 1955, 27, 1174–1175. [Google Scholar] [CrossRef]
- Zhou, B.; Wang, J.; Guo, Z.; Tan, H.; Zhu, X. A simple colorimetric method for determination of hydrogen peroxide in plant tissues. Plant Growth Regul. 2006, 49, 113–118. [Google Scholar] [CrossRef]
- Barnuevo, A.; Asaeda, T. Integrating the ecophysiology and biochemical stress indicators into the paradigm of mangrove ecology and a rehabilitation blueprint. PLoS ONE 2018, 13, e0202227. [Google Scholar] [CrossRef] [PubMed]
- Asaeda, T.; Senavirathna, M.D.H.J.; Krishna, L.V.; Yoshida, N. Impact of regulated water levels on willows (Salix subfragilis) at a flood-control dam, and the use of hydrogen peroxide as an indicator of environmental stress. Ecol. Eng. 2018, 127, 96–102. [Google Scholar] [CrossRef]
- Brennan, T.; Frenkel, C. Involvement of Hydrogen Peroxide in the Regulation of Senescence in Pear. Plant Physiol. 1977, 59, 411–416. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Queval, G.; Hager, J.; Gakière, B.; Noctor, G. Why are literature data for H2O2 contents so variable? A discussion of potential difficulties in the quantitative assay of leaf extracts. J. Exp. Bot. 2008, 59, 135–146. [Google Scholar] [CrossRef]
- Okazaki, Y.; Rao, S.; Asao, S.; Tateishi, T.; Katsuda, S.-I.; Furuki, Y. Effects of Ti, Al and V Concentrations on Cell Viability. Mater. Trans. JIM 1998, 39, 1053–1062. [Google Scholar] [CrossRef]
- Vilaplana, R.; Valentines, M.C.; Toivonen, P.; Larrigaudière, C. Antioxidant Potential and Peroxidative State of ‘Golden Smoothee’ Apples Treated with 1-Methylcyclopropene. J. Am. Soc. Hortic. Sci. 2006, 131, 104–109. [Google Scholar] [CrossRef]
- Lentheric, I.; Pinto, E.; Graell, J.; Larrigaudière, C. Effects of CO2 pretreatment on oxidative metabolism and core- browning incidence in controlled atmosphere stored pears. J. Hortic. Sci. Biotechnol. 2003, 78, 177–181. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M. Free Radicals in Biology and Medicine, 5th ed.; Oxford University Press: Oxford, UK, 2015. [Google Scholar]
- Demmano, G.; Selegny, E.; Vincent, J.-C. Experimental Procedure for a Hydrogen Peroxide Assay Based on the Peroxidase-Oxidase Reaction. JBIC J. Biol. Inorg. Chem. 1996, 238, 785–789. [Google Scholar] [CrossRef]
- Orozco-Cárdenas, M.L.; Ryan, C.A. Nitric Oxide Negatively Modulates Wound Signaling in Tomato Plants. Plant Physiol. 2002, 130, 487–493. [Google Scholar] [CrossRef]
- Thordal-Christensen, H.; Zhang, Z.; Wei, Y.; Collinge, D.B. Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley—Powdery mildew interaction. Plant J. 1997, 11, 1187–1194. [Google Scholar] [CrossRef]
- Yao, N.; Tada, Y.; Sakamoto, M.; Nakayashiki, H.; Park, P.; Tosa, Y.; Mayama, S. Mitochondrial oxidative burst involved in apoptotic response in oats. Plant J. 2002, 30, 567–579. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, A.; Creissen, G.; Kular, B.; Firmin, J.; Robinson, S.; Verhoeyen, M.; Mullineaux, P. Changes in oxidative processes and components of the antioxidant system during tomato fruit ripening. Planta 2002, 214, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.R.; Lee, K. Determination of environmental H2O2 for extended periods by chemiluminescence with real-time inhibition of iron interferences. Microchem. J. 2019, 147, 1021–1027. [Google Scholar] [CrossRef]
- Halliwell, B.; Whiteman, M. Measuring reactive species and oxidative damage in vivo and in cell culture: How should you do it and what do the results mean? Br. J. Pharmacol. 2004, 142, 231–255. [Google Scholar] [CrossRef]
- Patterson, B.D.; MacRae, E.A.; Ferguson, I.B. Estimation of hydrogen peroxide in plant extracts using titanium(IV). Anal. Biochem. 1984, 139, 487–492. [Google Scholar] [CrossRef]
- Li, J.; Lu, B.; Xu, L.L. An improved method for the determination of hydrogen peroxide in leaves. Progr. Biochem. Biophys. 2000, 27, 548–551. [Google Scholar]
- Gay, C.; Collins, J.; Gebicki, J.M. Hydroperoxide Assay with the Ferric–Xylenol Orange Complex. Anal. Biochem. 1999, 273, 149–155. [Google Scholar] [CrossRef]
- Veljovic-Jovanovic, S.; Noctor, G.; Foyer, C.H. Are leaf hydrogen peroxide concentrations commonly overestimated? The potential influence of artefactual interference by tissue phenolics and ascorbate. Plant Physiol. Biochem. 2002, 40, 501–507. [Google Scholar] [CrossRef]
- Grime, J.P.; Mackey, J.M.L. The role of plasticity in resource capture by plants. Evol. Ecol. 2002, 16, 299–307. [Google Scholar] [CrossRef]
- Cheeseman, J.M. Hydrogen peroxide concentrations in leaves under natural conditions. J. Exp. Bot. 2006, 57, 2435–2444. [Google Scholar] [CrossRef] [PubMed]
- Allen, D.J.; Ort, D.R. Impacts of chilling temperatures on photosynthesis in warm-climate plants. Trends Plant Sci. 2001, 6, 36–42. [Google Scholar] [CrossRef]
- Peng, C.; Crawshaw, J.P.; Maitland, G.C.; Trusler, J.M.; Vega-Maza, D. The pH of CO2-saturated water at temperatures between 308K and 423K at pressures up to 15 MPa. J. Supercrit. Fluids 2013, 82, 129–137. [Google Scholar] [CrossRef]
- Peng, P.; Hu, A.; Gerlich, A.P.; Zou, G.; Liu, L.; Zhou, Y.N. Joining of Silver Nanomaterials at Low Temperatures: Processes, Properties, and Applications. ACS Appl. Mater. Interfaces 2015, 7, 12597–12618. [Google Scholar] [CrossRef]
- Fei, H.; Dong, J.; Arellano-Jiménez, M.J.; Ye, G.; Kim, N.D.; Samuel, E.L.; Peng, Z.; Zhu, Z.; Qin, F.; Bao, J.; et al. Atomic cobalt on nitrogen-doped graphene for hydrogen generation. Nat. Commun. 2015, 6, 8668. [Google Scholar] [CrossRef]
- Lovelock, C.E.; Krauss, K.W.; Osland, M.J.; Reef, R.; Ball, M.C. The physiology of mangrove trees with changing climate. Trop. Tree Physiol. Adapt. Responses A Chang. Environ. 2016, 6, 149–179. [Google Scholar]
- Dautania, G.K.; Singh, G.P. Role of Light and Dark Cycle and Different Temperatures in the Regulation of Growth and Protein Expression in Oscillatoria agardhii Strain. Braz. Arch. Biol. Technol. 2014, 57, 933–940. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Bouché, N.; Fait, A.; Bouchez, D.; Møller, S.G.; Fromm, H. Mitochondrial succinic-semialdehyde dehydrogenase of the γ-aminobutyrate shunt is required to restrict levels of reactive oxygen intermediates in plants. Proc. Natl. Acad. Sci. USA 2003, 100, 6843–6848. [Google Scholar] [CrossRef] [PubMed]
- Karpinski, S.; Escobar, C.; Karpinska, B.; Creissen, G.; Mullineaux, P.M. Photosynthetic electron transport regu- lates the ex-pression of cytosolic ascorbate peroxidase genes in Arabidopsis during excess light stress. Plant Cell 1997, 9, 627–640. [Google Scholar] [PubMed]
- Karpinski, S.; Reynolds, H.; Karpinska, B.; Wingsle, G.; Creissen, G.; Mullineaux, P. Systemic signaling and acclimation in response to excess excitation energy in Arabidopsis. Science 1999, 284, 654–657. [Google Scholar] [CrossRef]
- Chaparzadeh, N.; D’Amico, M.L.; Khavari-Nejad, R.-A.; Izzo, R.; Navari-Izzo, F. Antioxidative responses of Calendula officinalis under salinity conditions. Plant Physiol. Biochem. 2004, 42, 695–701. [Google Scholar] [CrossRef]
- Tewari, R.K.; Kumar, P.; Tewari, N.; Srivastava, S.; Sharma, P.N. Macronutrient deficiencies and differential antioxidant responses—Influence on the activity and expression of superoxide dismutase in maize. Plant Sci. 2004, 166, 687–694. [Google Scholar] [CrossRef]
- Mur, L.A.J.; Kenton, P.; Draper, J. In planta measurements of oxidative bursts elicited by avirulent and virulent bacterial pathogens suggests that H2O2 is insufficient to elicit cell death in tobacco. Plant Cell Environ. 2005, 28, 548–561. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Ruelland, E.; Vaultier, M.N.; Curie, M.; Galile, R.; Seine, I. Cold signalling and cold acclimation in Plants. Adv. Bot. Res. 2009, 49, 35–150. [Google Scholar] [CrossRef]
- Miura, K.; Tada, Y. Regulation of water, salinity, and cold stress responses by salicylic acid. Front. Plant Sci. 2014, 5, 4. [Google Scholar] [CrossRef]
- Gu, L.; Hanson, P.J.; Mac Post, W.; Kaiser, D.P.; Yang, B.; Nemani, R.; Pallardy, S.G.; Meyers, T. The 2007 Eastern US Spring Freeze: Increased Cold Damage in a Warming World? Bioscience 2008, 58, 253–262. [Google Scholar] [CrossRef]
- Sevillano, L.; Sanchez-Ballesta, M.T.; Romojaro, F.; Flores, F.B. Physiological, hormonal and molecular mechanisms regulating chilling injury in horticultural species. Postharvest technologies applied to reduce its impact. J. Sci. Food Agric. 2009, 89, 555–573. [Google Scholar] [CrossRef]
- Luengwilai, K.; Beckles, D.M.; Saltveit, M.E. Chilling-injury of harvested tomato (Solanum lycopersicum L.) cv. Micro-Tom fruit is reduced by temperature pre-treatments. Postharvest Biol. Technol. 2012, 63, 123–128. [Google Scholar] [CrossRef]
- Lukatkin, A.S. Initiation and Development of Chilling Injury in Leaves of Chilling-Sensitive Plants. Russ. J. Plant Physiol. 2005, 52, 542–546. [Google Scholar] [CrossRef]
- Upadhyaya, H.; Khan, M.H.; Panda, S.K. Hydrogen peroxide induces oxidative stress in detached leaves of Oryza sativa L. Gen. Appl. Plant Physiol. 2007, 33, 83–95. [Google Scholar]
- Liu, Z.-J.; Guo, Y.-K.; Bai, J.-G. Exogenous Hydrogen Peroxide Changes Antioxidant Enzyme Activity and Protects Ultrastructure in Leaves of Two Cucumber Ecotypes Under Osmotic Stress. J. Plant Growth Regul. 2010, 29, 171–183. [Google Scholar] [CrossRef]
- Goud, P.B.; Kachole, M.S. Effect of exogenous hydrogen peroxide on peroxidase and polyphenol oxidase activities in Ca-januscajan (L.) Millsp.detached leaves. Int. J. Curr. Res. Rev. 2011, 3, 61–65. [Google Scholar]
- Kacperska, A. Metabolic Consequences of Low Temperature Stress in Chilling-Insensitive Plants. In Low Temperature Stress Physiology in Crops; CRC Press: Boca Raton, FL, USA, 2018; pp. 27–40. [Google Scholar] [CrossRef]
- Sharon, O.; Kahn, V. Browning Potential, PPO, Catalase and Acid Phosphatase Activities during Ripening of Non-chilled and Chilled Avocado. J. Sci. Food Agric. 1979, 30, 634–638. [Google Scholar] [CrossRef]
- Yang, T.; Poovaiah, B.W. Hydrogen peroxide homeostasis: Activation of plant catalase by calcium/calmodulin. Proc. Natl. Acad. Sci. USA 2002, 99, 4097–4102. [Google Scholar] [CrossRef]
- Rao, P.S.; Kalva, S.; Yerramilli, A.; Mamidi, S. Free Radicals and Tissue Damage: Role of Antioxidants. Free. Radic. Antioxid. 2011, 1, 2–7. [Google Scholar] [CrossRef]
- MacRae, E.A.; Ferguson, I.B. Changes in catalase activity and hydrogen peroxide concentration in plants in response to low temperature. Physiol. Plant. 1985, 65, 51–56. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| eFOX Assay | Ti(SO4)2 Assay | |||
|---|---|---|---|---|
| Species Name | Frozen | Nonfrozen | Frozen | Nonfrozen |
| Ambrosia trifida | r = −0.670, p = 532 | r = −0.977, p = 0.137 | r = 0.958, p = 186 | r = 0.957, p = 0.187 |
| Solidago altissima | r = −0.812, p = 0.397 | r = 0.946, p = 0.21 | r = 0.30, p = 0.806 | r = 0.302, p = 0.804 |
| Artemisia princeps | r = 0.984, p = 0.115 | r = 0.496, p = 0.67 | r = −0.162, p = 0.896 | r = −0.273, p = 0.824 |
| Sicyos angulatus | r = 0.528, p = 0.646 | r = 0.593, p = 0.596 | r = 0.931, p = 0.239 | r = 0.824, p = 0.383 |
| Species Name | Maximum Reduction Rate (%) in eFOX Assay | Maximum Reduction Rate (%) in Ti(SO4)2 Assay |
|---|---|---|
| Ambrosia trifida | 32.47 | 20.62 |
| Solidago altissima | 52.91 | 60.10 |
| Artemisia princeps | 65.94 | 52.90 |
| Sicyos angulatus | 30.28 | 26.10 |
| Species name | H2O2 Concentration (µmol/gFW) | Technique | Notes | Reference |
|---|---|---|---|---|
| Litchi chinensis Sonn. | 0.64 | Glucose peroxidase | Fully expanded leaves | [20] |
| Spinacia olearacea | 0.54 | Titanium (IV)-PAR complex. | Fully expanded leaves | [39] |
| Glycine max L. | 3.47 | Ferrous oxidation/xylenol orange | Fully expanded leaves | [43] |
| Calendula officinalis | 6 | Glycolate oxidase | Young leaves | [56] |
| Arabidopsis thaliana | 5 | Luminol chemiluminescence | Young leaves | [54,55] |
| Zea mays L. | 75 | Ti(SO4)2 assay | Fully expanded leaves | [57] |
| Nicotinia tabacum | 5 | Titanium-peroxide complex assay | Fully expanded leaves | [58] |
| Rice (Oryza sativa L.) | 0.26 | Titanium (IV) sulphate- TiCl4 complex | Young leaves | [40] |
| Wheat (Triticum aestivum L.) | 0.58 | Titanium (IV) sulphate- TiCl4 complex | Old leaves | [40] |
| Banana (Musa acuminata Colla) | 0.72 | Glucose peroxidase | Fully expanded leaves | [20] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, M.; Asaeda, T.; Fukahori, K.; Imamura, F.; Nohara, A.; Matsubayashi, M. Hydrogen Peroxide Measurement Can Be Used to Monitor Plant Oxidative Stress Rapidly Using Modified Ferrous Oxidation Xylenol Orange and Titanium Sulfate Assay Correlation. Int. J. Plant Biol. 2023, 14, 546-557. https://doi.org/10.3390/ijpb14030043
Rahman M, Asaeda T, Fukahori K, Imamura F, Nohara A, Matsubayashi M. Hydrogen Peroxide Measurement Can Be Used to Monitor Plant Oxidative Stress Rapidly Using Modified Ferrous Oxidation Xylenol Orange and Titanium Sulfate Assay Correlation. International Journal of Plant Biology. 2023; 14(3):546-557. https://doi.org/10.3390/ijpb14030043
Chicago/Turabian StyleRahman, Mizanur, Takashi Asaeda, Kiyotaka Fukahori, Fumiko Imamura, Akio Nohara, and Masaomi Matsubayashi. 2023. "Hydrogen Peroxide Measurement Can Be Used to Monitor Plant Oxidative Stress Rapidly Using Modified Ferrous Oxidation Xylenol Orange and Titanium Sulfate Assay Correlation" International Journal of Plant Biology 14, no. 3: 546-557. https://doi.org/10.3390/ijpb14030043
APA StyleRahman, M., Asaeda, T., Fukahori, K., Imamura, F., Nohara, A., & Matsubayashi, M. (2023). Hydrogen Peroxide Measurement Can Be Used to Monitor Plant Oxidative Stress Rapidly Using Modified Ferrous Oxidation Xylenol Orange and Titanium Sulfate Assay Correlation. International Journal of Plant Biology, 14(3), 546-557. https://doi.org/10.3390/ijpb14030043