Abstract
The rising global demand for food, including potatoes, necessitates increased crop production. To achieve higher yields, farmers frequently depend on regular applications of nitrogen and phosphate fertilizers. As people seek more environmentally friendly alternatives, biofertilizers are gaining popularity as a potential replacement for synthetic fertilizers. This study aimed to determine how Glomus iranicum affects the growth of potatoes (Solanum tuberosum L.) and the nutritional value of potato tubers when grown alongside broad beans (Vicia faba L.). An experiment was conducted using potatoes tested at five dosage levels of G. iranicum, ranging from 0 to 4 g, to see its impact on the plants and soil. Inoculation with G. iranicum produced variable results in associated potato and bean crops, with significant effects on some variables. In particular, inoculation with 3 g of G. iranicum produced an increase in plant height (24%), leaf dry weight (90%), and tuber dry weight (57%) of potatoes. Similarly, 4 g of G. iranicum produced an increase in the foliar fresh weight (115%), root length (124%), root fresh weight (159%), and root dry weight (243%) of broad beans compared to no inoculation. These findings suggest that G. iranicum could be a helpful biological tool in Andean crops to improve the productivity of potatoes associated with broad beans. This could potentially reduce the need for chemical fertilizers in these crops.
1. Introduction
The escalating global demand for food is driving the need for higher crop yields, often achieved through unsustainable agricultural practices, such as the overuse of fertilizers, herbicides, and pesticides [1,2,3]. These practices can significantly alter the soil’s physical, chemical, and biological properties [4,5,6]. Herbicide use can diminish the microbial community, including key organisms like ammonium-oxidizing bacteria and archaea [7].
Potato production often relies on continuous applications of nitrogen and phosphate fertilizers to maintain high yields, a practice ingrained with the green revolution model [8]. However, this practice of fertilizer use can negatively affect biodiversity, nutrient balance, soil compaction, and erosion [4]. Additionally, it can alter the soil’s chemical properties, including cation exchange capacity, organic carbon content, and mineral levels [9]. Low levels of organic carbon in soil can decrease crop productivity [7].
Intercropping is a common practice that offers multiple benefits, such as nitrogen fixation [10], increased nutrient availability [11], improvement in agroecosystem services [12], and improved productivity and sustainability in smallholder farming systems [13]. In the high Andes, staple crops including potato (S. tuberosum), wheat (Triticum aestivum), maize (Zea mays), and broad bean (V. faba) are normally produced using monocultures or alternating crops [14].
Legumes offer a sustainable solution to nitrogen deficiencies in soil by forming symbiotic relationships with nitrogen-fixing bacteria (rhizobia). The development of nodules on legume roots is a direct indicator of their nitrogen-fixing ability [15,16,17]. Beyond nitrogen fixation, legumes also promote sustainable agriculture by efficiently utilizing land resources, increasing soil biodiversity, and improving nutrient cycling [18]. Moreover, legume residues decompose more rapidly than cereal residues, contributing significantly to soil organic carbon levels [19].
Arbuscular mycorrhizal fungi (AMF) establish symbiosis with most terrestrial plants [10,11], promoting their growth and the uptake of nutrients such as phosphorus and nitrogen [20,21,22]. They improve the root network through the extraradical mycelium [23], release organic acids and phosphatases that solubilize essential nutrients [5], and stimulate the production of phytohormones such as auxins, cytokinins, and gibberellic acid [24]. In addition, they increase tolerance to biotic and abiotic stresses [25,26,27] and contribute to soil aggregation by forming micro- and macroaggregates through their extraradical hyphae [28,29].
The search for diversified agricultural systems that optimize soil resource use and minimize dependency on external inputs is increasingly important [30,31]. In the Andes, rotational and intercropping systems have been widely adopted due to their ability to enhance sustainability and productivity [14,32,33].
One promising approach is combining potato as the main crop with broad beans, leveraging the latter’s capacity for nitrogen fixation. This synergy has potential benefits that remain underexplored in practice [34]. Furthermore, introducing inoculations of arbuscular mycorrhizal fungi could amplify these advantages by promoting nutrient uptake and enhancing soil health [35,36,37].
This combination not only improves soil fertility but also offers an ecological advantage by creating a more balanced and sustainable cropping system. Such benefits are particularly significant in areas with less fertile soil, where maintaining long-term productivity is a critical challenge for farmers [38,39].
This study aimed to explore novel combinations of the main crop, potato, in association with a leguminous plant, such as the fava bean, inoculated with the commercial biostimulant MycoUp with the fungus G. iranicum to maximize crop growth performance. Specifically, the objectives were to (1) assess the effect of the mycorrhizal fungus on potato yield, (2) evaluate the influence of the fungus on broad bean growth, and (3) examine the content of phosphorus (P) and potassium (K) in tubers and their availability in the soil.
Importance and Applications of Biofertilizers in Agricultural Crops
Biofertilizers have emerged as a promising solution to the challenges of sustainable agriculture, especially in the context of a growing world population and limitations on the availability of agricultural land. These products, which use beneficial microorganisms, offer an ecological alternative to synthetic fertilizers, which have been shown to have adverse effects on the environment and human health [40,41,42]. These biofertilizers can be grouped according to their functions and mode of action, the most commonly used being nitrogen fixers, potassium solubilizers, phosphorus solubilizers, and rhizobacteria that promote plant growth [43]. In our case, we will focus on (i) nitrogen fixers, (ii) phosphorus solubilizers, and (iii) potassium solubilizers.
As for (i) nitrogen fixers, the most commonly used inoculants are Azotobacter, Rhizobium, Azospirillum, and Azolla. These inoculants help in nitrogen fixation by converting atmospheric nitrogen into a form usable by the plant [44]. A global meta-analysis study showed that biofertilizers increase the average yield of plants (16.2 ± 1.0%) compared to non-inoculated plants, as well as efficiency in the use of nitrogen and phosphorus, where legumes showed a superior trend compared to other plants [45]. In terms of productivity, the use of Cyanobacteria and Azolla complements the nitrogen requirements of rice plants, replacing between 30 and 50% of synthetic nitrogen [46]. In another more extensive trial, the effects of biofertilizer produced from strains of Bradyrhizobium spp. and Streptomyces spp. were tested on legume, cereal and vegetable crops, where a significant increase was found in plant growth, nodulation, nitrogen fixation, and the absorption of nitrogen, phosphorus, and potassium in soy and beans [47].
Similarly, (ii) phosphorus solubilizers contribute to disease resistance and help plants to be more resistant [48,49]; phosphorus can be solubilized by many microorganisms, such as bacteria, some fungi, and actinomycetes [50]. The most commonly used inoculates are mainly from the genera Pseudomonas, Bacillus, Rhodococcus, Chryseobacterium, Phyllobacterium, and Xanthomona [51,52]. As an example of applications, we can mention the exploration of the role of Bacillus spp. in improving the availability of phosphorus and promoting the growth of sugarcane plants in Thailand [53], as well as the improvement of soil fertility, nutrient absorption, and the yield of Sesamum indicum through the application of purple bacteria biofertilizers compared to the application of synthetic fertilizers [54]. Dual applications of biofertilizers with phosphorus promote the absorption of nutrients by chickpeas and the accumulation of nutrients in the soil after harvest, which vary according to the doses applied [55].
Potassium is one of the most important nutrients for plant growth [56]. As for (iii) potassium solubilizers, the literature reports that Aspergillus, Bacillus, Clostridium, Burkholderia, and Pseudomonas, among others, are the main ones that transform potassium into a form accessible to plants [57]. Various studies in different parts of the world report the beneficial effects of potassium-solubilizing bacteria on the growth of various crops. For example, grapes and cotton plants showed a significant increase in plant height, leaf length/width, fruit size, and the number of fruits per plant compared to controls [58]. In wheat, bacterial inoculation significantly increased the nitrogen, phosphorus, and potassium content [59] to levels very similar to those found in Sorghum vulgare, where an increase in biomass and nutrient acquisition was found with co-inoculation applications of waste mica with B. mucilaginosus and A. chroococcum [60]; trials with Bacillus inoculations in nutrient-limited soils planted with cucumber and pepper showed a significant improvement in the availability of phosphorus and potassium in the soil and the absorption of N, P, and K by shoots and roots [61].
2. Materials and Methods
2.1. Description of Site, Plants, and Fungi
The experiment was carried out at the National Institute of Agricultural Innovation (INIA), Santa Ana Agricultural Experimental Station, located in the district of Tambo, Province of Huancayo, Department of Junin, which is geographically located at 12°00′40″ S, longitude 75°13′17″ W, at 3295 m s.a.l. The average temperature inside the greenhouse was 18.6 °C, with a relative humidity of 46% measured with a Boeco® thermohygrometer (BOECO, Hamburg, Germany).
Physicochemical analyses of soil samples were performed for the following parameters: electrical conductivity (EC), pH, organic matter (OM, %), phosphorus content (P, mg/kg), potassium content (K, mg/kg), and texture (% sand, clay and silt). The EC was assessed according to ISO 11265 [62]. The pH was determined using method 9045D [63]. OM was determined by the Walkley–Black method [64]. Phosphorus content was evaluated by the method of Bray and Kurtz [65]. Potassium content was determined by method NOM-021-RECNAT-2000 [66]. Soil texture was determined using the Bouyoucos method [67]. These analyses were carried out at the Soil Laboratory of the Santa Ana Experimental Station of the National Institute for Agrarian Innovation (INIA).
A moderately acid pH was found, which could influence the assimilation of some elements. Normal electrical conductivity was found, without restriction on the crops. We used medium organic matter, suitable for agricultural soils. The available phosphorus was high and accessible to plants. Available potassium was low, and total nitrogen was very low. The soil was loam-textured and suitable for agricultural cultivation due to its balance between water retention, drainage, aeration, and fertility. For more details, see Table 1.

Table 1.
Basic physiochemical properties of cultivated soil.
Results were issued by the Soil, Water and Foliar Laboratory of the National Institute for Agrarian Innovation (INIA), where EC = electrical conductivity, OM = medium organic matter (OM), P = phosphorus, and K = potassium.
The biological material used in this research was the potato (S. tuberosum) “revolution” variety and the broad bean (V. faba) of the “yellow” variety. The seeds came from the previous harvests of the INIA stations; seeds were selected to be as uniform as possible in size and free of wounds and rotting.
The commercial biostimulant MycoUp® (Symborg, Murcia, Spain) was used, composed exclusively of the arbuscular mycorrhizal fungus G. iranicum var. Tenuihypharum. According to the product indications, the substrate was a clay mineral, with a concentration of 120 propagules per gram of substrate.
2.2. Experimental Design
The trial was conducted from September to December 2023 under greenhouse conditions, using a completely randomized design (CRD). It included five levels of commercial biostimulant MycoUp and four replicates: T1 (0 g); T2 (1 g); T3 (2 g), T4 (3 g) and T5 (4 g). The experimental unit was composed of 5 kg of sieved soil, one potato seed associated with two broad bean seeds, and various doses of the biostimulant MycoUp, according to the treatments (Figure 1). Inoculation was performed at the beginning of the trial, and irrigation was carried out three times a week with 250 mL of tap water per experimental unit.

Figure 1.
Processes carried out in the greenhouse and in the laboratory. In (a), the development of potato and bean plants in the greenhouse is shown; (b) shows the evaluation of the height of the bean plant; (c) shows the obtainment of the number of tubers and length of the potato root and beans; and (d) shows the cleaning and preparation of the bean root for drying in the oven.
2.3. Measurement of Plant Growth Parameters
In December 2024, the harvest was carried out according to the physiological maturity of the potato (100 days after planting), contrasting with the wilting of leaves, adopted from [68]. The height of the fresh vegetative parts was measured with a 50 cm metal ruler, and their fresh weight was obtained on an analytical balance with a precision of 0.01 g; in the case of the broad bean, the number of flower buds was counted. Root length, root fresh weight, and the number of tubers in the potato were obtained from the terrestrial part.
The vegetative parts, roots, and tubers were dried at 65 °C for 24 h to determine the dry matter content. Phosphorus (P) and potassium (K) contents in potato tubers were determined in the soil, water and foliar laboratory (LABSAF) of INIA Santa Ana, and the available phosphorus and potassium in the post-harvest soil were also analyzed.
2.4. Statistical Analysis
To determine if the data followed a normal distribution, the Shapiro–Wilk test was used (p-value > 0.05 indicated a normal distribution). Levene’s test was employed to check for equal variances (p-value > 0.05 indicated equal variances). To compare the means between groups, a one-way analysis of variance (ANOVA) was conducted, followed by Tukey’s post-hoc test (statistical significance was defined as a p-value < 0.05). All statistical analyses, including descriptive statistics, the ANOVA, and post-hoc tests, were performed using the R programming language (version 4.3.3 for Windows) with publicly available libraries and custom-written scripts. In addition, a principal component analysis (PCA) was performed in order to see which variables were most associated with each type of treatment.
3. Results
3.1. Plant Growth Parameters
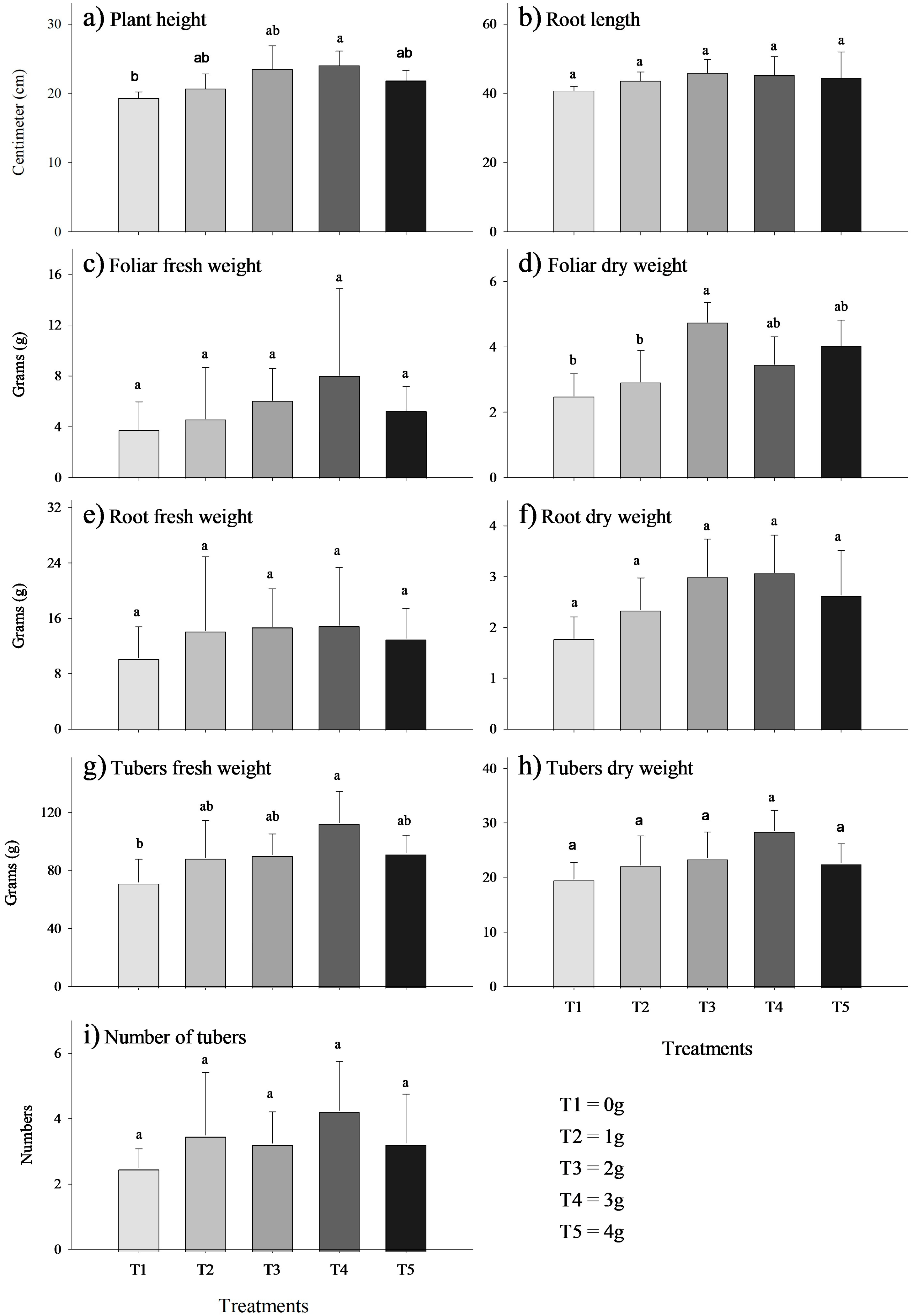
The MycoUp biostimulant demonstrated a dose-dependent increase in efficacy, with higher doses (2 g and 3 g) showing a more pronounced effect compared to the control (0 g). In some instances, the increase became statistically significant (p-value < 0.05) at moderate doses of 2 g and 3 g of MycoUp biostimulant. Significant increases occurred in plant height (24%, Figure 2a), foliar dry weight (90% g, Figure 2d), and tuber fresh weight (57%, Figure 2g), while no significant differences were found in root length (Figure 2b), foliar fresh weight (Figure 2c), root fresh weight (Figure 2e), root dry weight (Figure 2f), tuber dry weight (Figure 2h), and the number of tubers (Figure 2i).
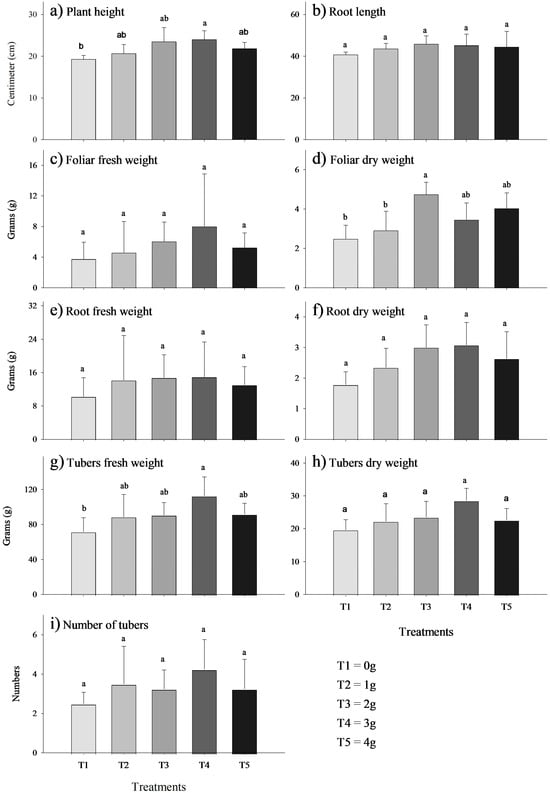
Figure 2.
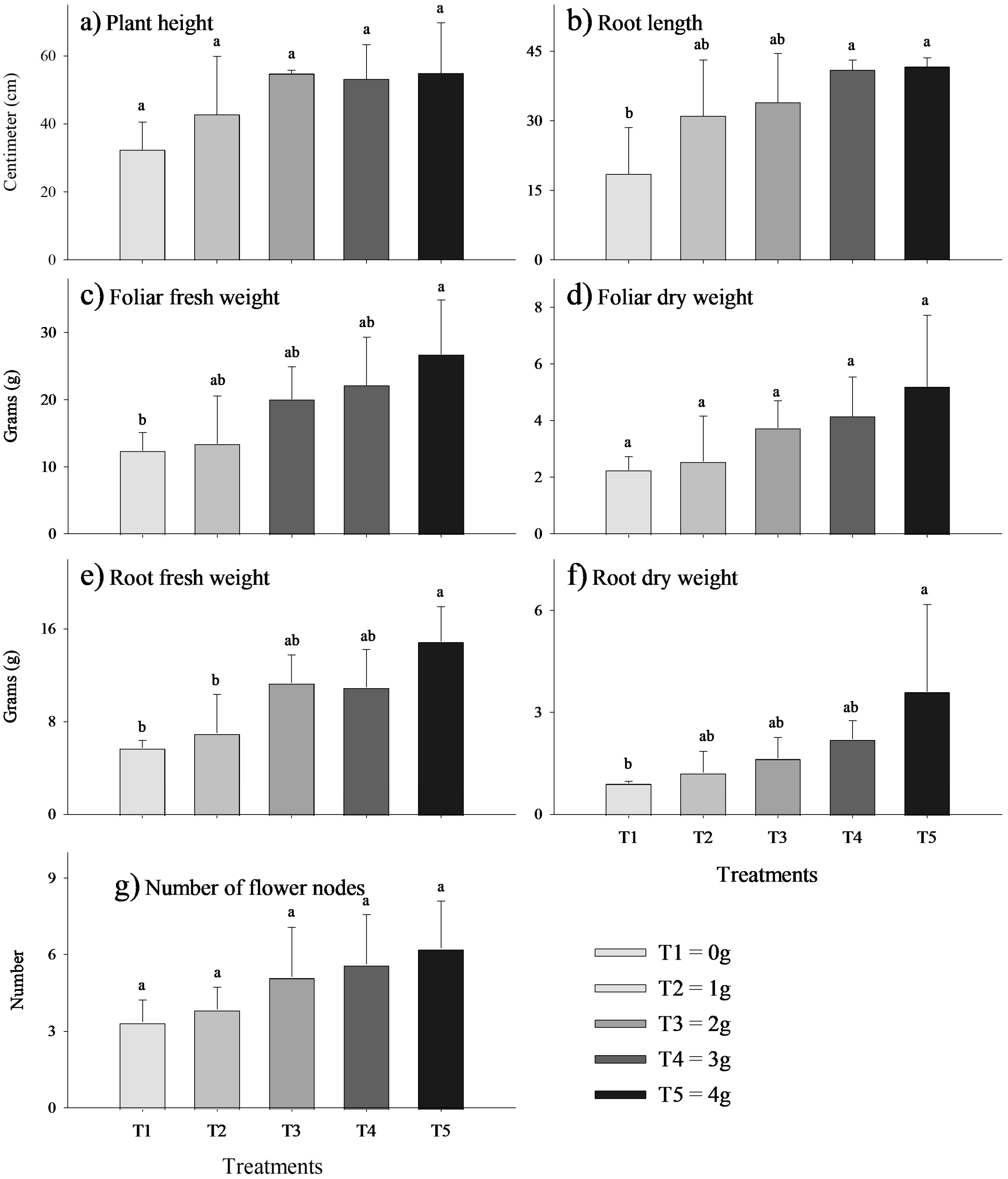
Effect of the biostimulant MycoUp mycorrhization on S. tuberosum plant characteristics: plant height (a), root length (b), foliar fresh weight (c), foliar dry weight (d), root fresh weight (e), root dry weight (f), tuber fresh weight (g), and dry weight (h) of potato, and number of tubers for treatment (i). Values with the same letter represent means that did not differ significantly (95%), according to Tukey’s HSD test. Error bars represent the standard deviation.The growth of V. faba plants treated with the biostimulant MycoUp exhibited significant increases in several parameters, including root length (cm), fresh leaf weight (g), fresh root weight (g), and dry root weight (g), as illustrated in Figure 3. In particular, significant increases (p-value < 0.05) were found in root length (Figure 3b) with a concentration of 3 g (120%) and 4 g (124%), and similarly in foliar fresh weight (Figure 3c), (115%), root fresh weight (Figure 3e) (159%), and root dry weight (Figure 3f) inoculated with 4 g (243%).
3.2. Tuber and Soil Nutrient Contents
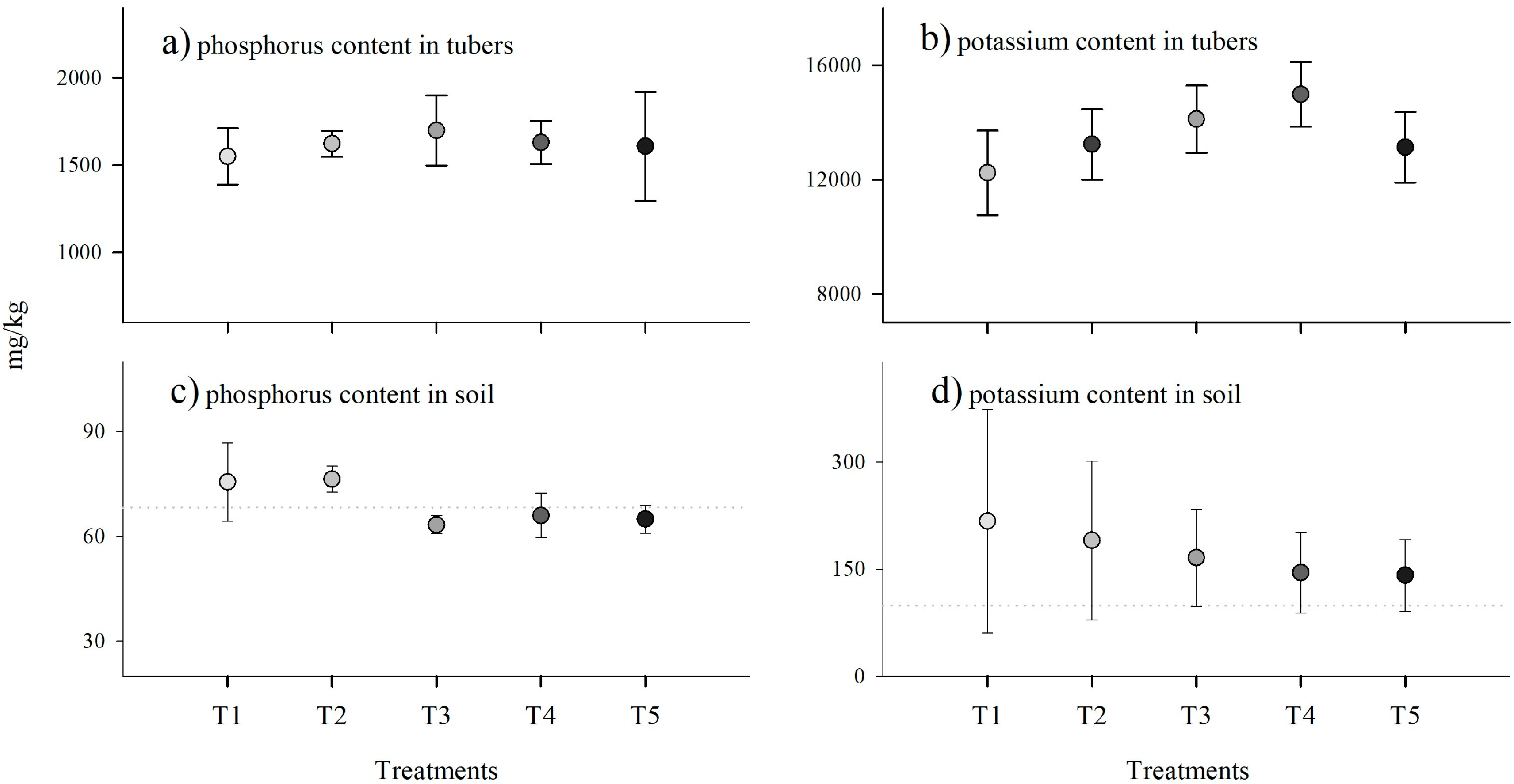
The analysis of phosphorus and potassium content in the tubers showed that there was no significant statistical difference with increasing inoculant concentration (Figure 4). Figure 4a shows that T3 (2 g) presented higher phosphorus content in the tubers (9%), while the smallest value was observed at T1 (0 g). Regarding potassium content (Figure 4b), an increase was observed as the dose increased from 0 g to 3 g (22%).
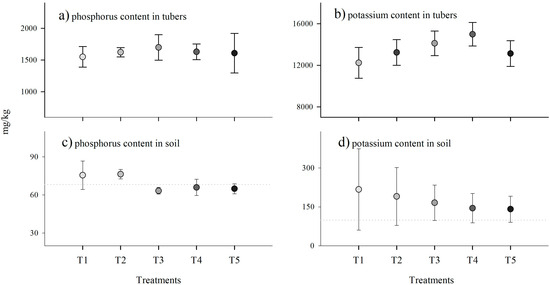
Figure 4.
Averages with their respective standard deviations for each treatment: (a) phosphorus content and (b) potassium content in tubers, (c) phosphorus content, and (d) potassium content in soils at the end of harvest. Gray dotted lines show the soil’s initial phosphorus and potassium levels. Darker shades of gray represent higher levels of G. iranicum.
Soil phosphorus content at the end of harvest remained within the initial range of 68.3 mg/kg. In general, there was a decrease in soil phosphorus content as the dose of biostimulant increased (Figure 4c); the highest value was found at T1 (75.5 mg/kg) and the minimum in T3 (63.31 mg/kg). The potassium content in the soil at the end of the harvests was higher than the initial content of 98.8 mg/kg, but the potassium content decreased as the dose of biostimulant increased (Figure 4d). For example, T1 presented a value of 216.8 mg/kg, T3 165.69 mg/kg, and T5 141.14 mg/kg. In addition, the variability followed the same pattern. For example, with T1, a minimum value of 94.97 mg/kg and a maximum value of 393 mg/kg were observed. In contrast, with T5, the variability was lower, with a minimum value of 106.93 mg/kg and a maximum value of 198.62 mg/kg (Figure 4c).
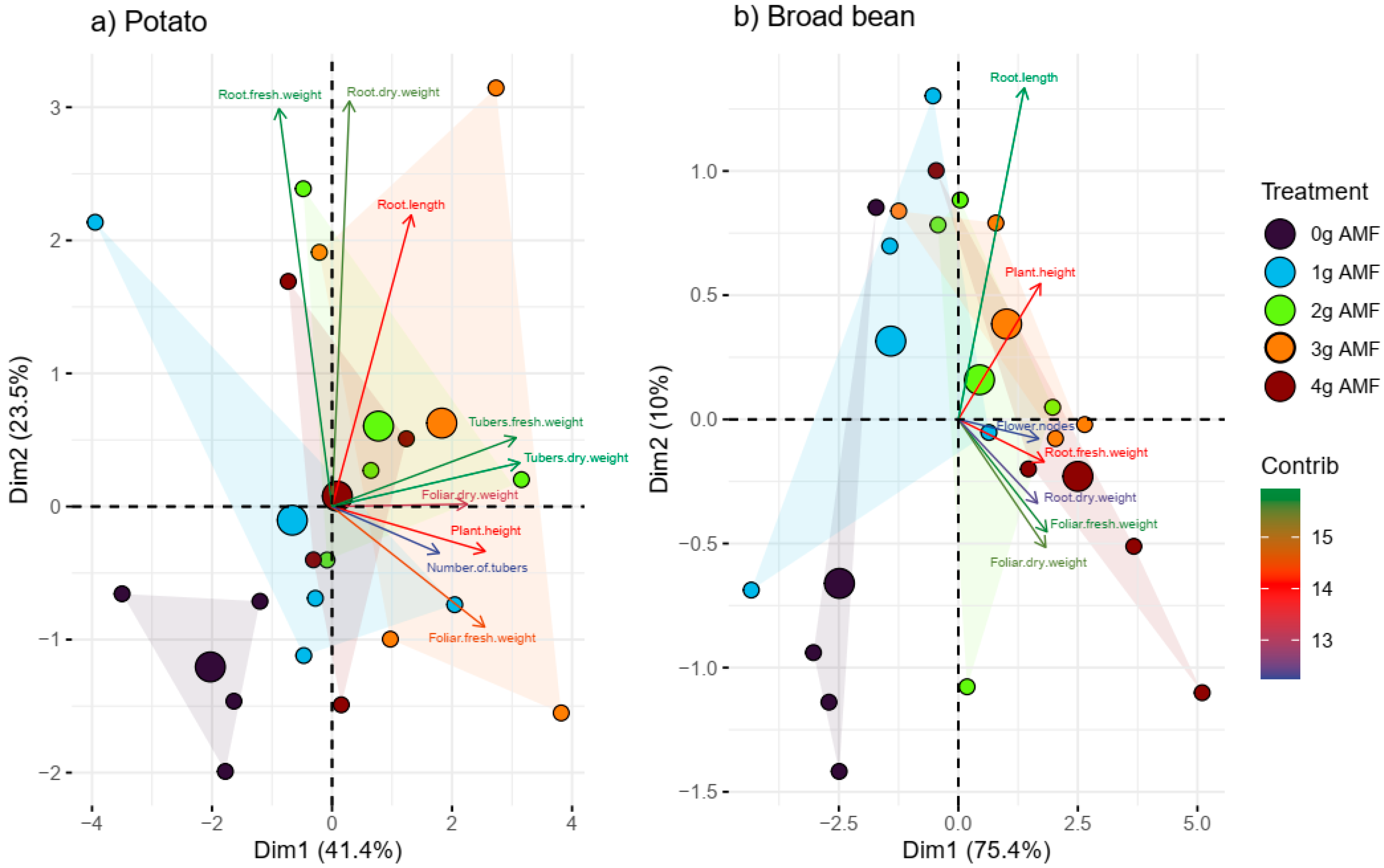
In PCA (Figure 5a), the first component (Dim1) explains 41.4% of the total variability in the potato data. This component is mainly related to the fresh and dry weight of tubers and roots. The second component (Dim2) explains 23.5% of the total variability and seems to be more associated with root length and dry leaf weight of the potato crop. Together (Dim1 and Dim2), they explain 64.9% of the total variability, which means that they capture most of the relevant information of the study.
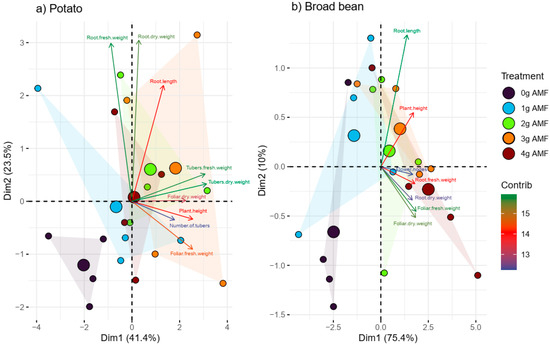
Figure 5.
Principal component analysis (a) using potato plants and (b) broad beans with different doses of biostimulant. The parameters evaluated are shown in the figures.
The first component (Dim1) shows clusters of tuber fresh/dry weight and root fresh/dry weight associated with positive values, suggesting that intermediate doses of the inoculant may be related to increased tuber and root development. On the contrary, Dim2 indicates that root length is favored by moderate inoculations.
In PCA (Figure 5b), the first component (Dim1) explains 75.4% of the total variability. This component is mainly related to the fresh and dry weight of roots and foliage, as well as the number of flowers. The second component (Dim2) explains 10% of the total variability and seems to be more associated with root length and plant height. Together (Dim1 and Dim2), they explain 85.4% of the total variability, and this means that they capture the relevant information in the dataset well.
In the first component (Dim1), there are groupings of the fresh/dry weight of tubers and fresh/dry weight of roots associated with positive values of the first component, which suggests that high doses of the inoculant (3 and 4 g) may be related to greater plant development. In contrast, (Dim2) indicates that root length is favored by moderate inoculations (1 g).
4. Discussion
The analyses reveals that the biostimulant (containing G. iranicum) applied to intercropped potato and bean crops yielded inconsistent results across the evaluated variables. While significant responses were observed in specific cases and at certain doses, such as plant weight, foliar dry weight, root dry weight, and root fresh weight, the overall effects varied (Figure 2 and Figure 3). This is attributed to arbuscular mycorrhizal fungi and their symbiotic ability to provide essential nutrients to host plants, mainly phosphorus and nitrogen, which favor their growth and development [69,70]. In addition, crops associated with leguminous plants favor the growth and development of the main crop [71].
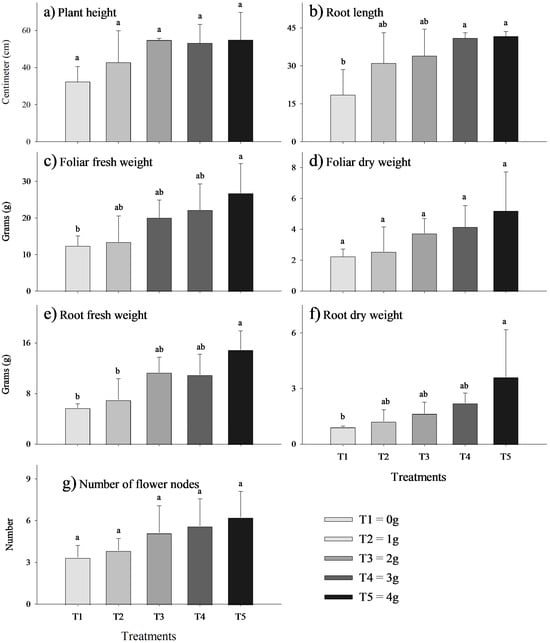
Figure 3.
Effect of biostimulant on V. faba plant characteristics: plant height (a), root length (b), foliar fresh weight (c), foliar dry weight (d), root fresh weight (e), root dry weight (f), and number of flower nodes (g). Values with the same letter represent means that did not differ significantly (p-value < 0.05) according to Tukey’s HSD test. Error bars represent the standard deviation.
The beneficial effects on plant growth and nutrient absorption are driven by various biological and biochemical mechanisms, such as (i) symbiotic associations of G. iranicum with plant roots, expanding the effective root network through a structure called the extraradical mycelium, which markedly improves the efficiency in the acquisition of these nutrients [72], and (b) AMF, such as G. iranicum, which are able to release organic acids and phosphatases that solubilize phosphorus and other essential elements present in unavailable forms in the soil that help to transform insoluble phosphorus into forms accessible to plants [5].
Crop association coupled with inoculation results in significant increases in plant height, foliar dry mass, and tuber fresh weight compared to non-inoculated plants (Figure 2). The increase in plant size by 24% compared with the non-inoculated plants was similar to previous studies with different inoculants such as Rhizophagus intraradices and Funneliformis mosseae [10,73,74,75], which demonstrates that inoculants of AMF benefit plant growth. As for the increase in foliar dry mass (Figure 2d), it was similar to that reported in other types of agricultural crops such as corn, rice, lettuce, grapes, and potatoes [21,72,76,77,78]. This effect is attributed to the fact that AMF produce several plant growth regulators, such as auxins, cytokinins, and gibberellins, in the roots of host plants [69,78,79]. These hormones are related to plant growth and development, promoting cell elongation and the development of new roots, which optimize water and nutrient absorption capacity and consequently improve overall plant productivity [24].
For the root parameters evaluated, an increase in root length, root fresh weight, and root dry weight was observed when the AMF dose was increased from 0 g to 2 or 3 g, although none of these changes were statistically significant (Figure 2b,e,f). These results contrast with previous studies that reported significant increases in root length in Populus species and Vitis vinifera cultivars [80,81]. Similarly, other studies have noted an increase in the number of lateral roots in Acacia auriculiformis and Acer saccharum [82,83].
Regarding tuber numbers, a proportional increase was observed with increasing doses of AMF up to 3 g (Figure 2i); however, these changes were not statistically significant. This increase can be attributed to the ability of mycorrhizae to infect potato roots, enhancing plant development and resulting in a higher number of tubers per pot [84], as reported by [85], with a mean between 5 and 11.33 tubers per pot. This figure exceeds the number reported in our study, which averaged 4.55 tubers. Arbuscular mycorrhizal fungi (AMF) release hormones that promote vegetative and root growth, leading to increased cell division, tissue expansion, and ultimately a higher number of tubers in potato crops [84]. The dose of 3 g AMF seemed to be more effective than 4 g AMF, possibly because it facilitated a balanced colonization of the roots without overloading the plants or creating excessive competition among the fungi; therefore, we note that the efficiency of the symbiosis and the increases in the analyzed potato variables did not always correlate with the amount applied.
Figure 3 illustrates the positive effect of G. iranicum inoculations on several evaluated parameters of the broad bean. We draw particular attention to root size and weight, as these variables more effectively demonstrate the effect of the inoculations. In general, a positive effect was observed as the concentrations increased from 0 to 3 and 4 g of AMF. Significant effects were observed on root length (Figure 3b), foliar fresh weight (Figure 3c), and root dry and fresh weight (Figure 3e,f). Our trials showed better results than that of Pereira et al., (2019) [25] when applying dual inoculations with bacteria and fungi. In contrast, they are similar to previous studies where foliar and root biomass increased with inoculation of rhizobia, AMF, and co-inoculates [15,16,20]. In terms of biomass, the positive effect of inoculations on the biomass of the associated crop is similar to that reported in other crops such as bell pepper and maize [86,87,88]. Increases in dry matter can be attributed to increases in nodulation and phosphorus fixation, as well as improved nutrient acquisition [20]. Certain arbuscular mycorrhizal fungi, such as G. iranicum, have been shown to enhance nutrient uptake and improve tolerance to salt stress [89,90]. This may account for the observed increase in fresh leaf weight without a corresponding rise in dry weight in faba bean plants.
Figure 4a,b, shows an increase in phosphorus (9%) and potassium (22%) content in tubers, but this was not significant compared to non-inoculated tubers. The efficacy of inoculants can be assessed by the increase in root colonization and nutrient uptake upon their application [91], on which many studies focus. For example, significant increases in the P (3%) and K (77%) were associated with the colonization of native arbuscular mycorrhizae [78]. We also found an increase of 0.35% in P with the inoculation of Rhizophagus irregularis [92].
The PCA analysis performed on the yield parameters of potato and broad bean plants summarizes the different responses to inoculations with G. inaricum. The first two components of the PCA explained 64% of the total variance observed for potatoes and 75% of that observed in broad beans (Figure 5a,b), similar to the variances reported for Nicotiana tabacum, Vicia faba, and Cucumis melo [25,69,93]. These percentages exceeded 25% of the total variance, according to [94], which is sufficient for a good interpretation. PCA analysis showed that the most relevant variables in differentiating treatments are the fresh and dry weight of tubers and roots, as well as plant height and number of tubers. Likewise, the PCA analysis for the beans showed that the most important variables in the differentiation of treatments are the fresh and dry weight of the root and the foliage and that these are related to high doses of inoculant.
The presence of fava beans (Vicia faba) in the experimental system could have influenced the results observed due to their capacity to fix atmospheric nitrogen through symbiosis with bacteria of the genus Rhizobium. This process improves the availability of nitrogen in the soil, indirectly benefitting potatoes (S. tuberosum), which could explain part of the increase in the growth and accumulation of nutrients in the tubers [95]. In addition, V. faba can modify the soil microbiome, creating a more favorable environment for the activity of arbuscular mycorrhizal fungi (G. iranicum), which amplifies the positive effects in the mixed cropping system [96]. These interactions reflect real agricultural conditions, where associated crops tend to benefit each other through the exchange of resources and the improvement of soil properties [97]. However, although these interactions are complex, studying them is essential for understanding and optimizing sustainable agricultural systems, particularly in regions such as the Andes, where intercropping is a common practice to maximize the productivity and resilience of agroecosystems [98].
5. Conclusions
We conclude that the arbuscular mycorrhizal fungus G. iranicum can be used to improve the production of potatoes associated with broad beans. A variable increase was observed in the evaluated parameters, though the trends were not consistent across all cases. Significant effects were detected at intermediate doses in potatoes and at high doses in broad beans. Using the commercial inoculum at doses of 2 g and 3 g of AMF, we observed an increase in plant height, dry biomass, and tuber weight. On the other hand, doses of 3 g and 4 g of AMF produced an increase in root length, fresh foliar weight, and the fresh and dry root weight of the broad beans. These results show the first signs of the potential of this inoculum in Andean crops and as a biological tool with which to improve the growth and yield of potato and fava bean production and suggest the need for further experiments in other Andean crops to reduce the need for chemical fertilizers.
Author Contributions
D.L.C.-P. and P.V.-M. designed the methodology; D.L.C.-P. provided and validated the data, S.P. and P.V.-M. performed the data processing; E.J.R.-R. and S.P. analyzed the data; R.S.-A. managed funding acquisition, provisioning, and characterization of microorganism strains, E.J.R.-R., S.P., P.V.-M. and D.L.C.-P. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the INIA project “Mejoramiento de los servicios de investigación y transferencia tecnológica en el manejo y recuperación de suelos agrícolas degradados y aguas para riego en la pequeña y mediana agricultura en los departamentos de Lima, Áncash, San Martín, Cajamarca, Lambayeque, Junín, Ayacucho, Arequipa, Puno y Ucayali” (CUI 2487112), of the Ministry of Agrarian Development and Irrigation (MIDAGRI) of the Peruvian Government.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Our data will be shared on request to the authors.
Acknowledgments
In collaboration with LABSAF teams, who provided the essential infrastructure and equipment, soil laboratory analysis was conducted. We extend our sincere gratitude to Elder Ofelia Pinillos Monge, head of the Plant Protection and Biocontrol Laboratory of the Santa Ana Agriculture Center, for the invaluable use of their facilities in preparing microbial inoculants.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Kumar, R.; Bhardwaj, A.; Singh, L.P.; Singh, G. Quantifying Ecological Impacts: A Comparative Life Cycle Assessment of Conventional and Organic Potato Cultivation. Ecol. Modell. 2023, 486, 110510. [Google Scholar] [CrossRef]
- Mrabet, R. Sustainable Agriculture for Food and Nutritional Security. In Sustainable Agriculture and the Environment; Press: Cambridge, MA, USA, 2023; pp. 25–90. ISBN 9780323905008. [Google Scholar]
- Odewale, S.A.; Odekanle, E.L.; Fakinle, B.S. Global Environmental Sustainability and Agrochemical Use. In One Health Implications of Agrochemicals and Their Sustainable Alternatives; Springer Nature: Singapore, 2023; pp. 735–764. ISBN 978-981-99-3439-3. [Google Scholar]
- Haverkort, A.J.; de Ruijter, F.J.; van Evert, F.K.; Conijn, J.G.; Rutgers, B. Worldwide Sustainability Hotspots in Potato Cultivation. 1. Identification and Mapping. Potato Res. 2013, 56, 343–353. [Google Scholar] [CrossRef]
- Meena, K.K.; Kumar, P.; Sorty, A.M.; Bitla, U.; Pathak, H. Ecology of Arbuscular Mycorrhizae and Influence on Drought Tolerance in Crop Plants. In Microbial BioTechnology for Sustainable Agriculture Volume 1; Springer: Singapore, 2022; pp. 261–285. ISBN 978-981-16-4843-4. [Google Scholar]
- Arévalo-Gardini, E.; Canto, M.; Alegre, J.; Loli, O.; Julca, A.; Baligar, V. Changes in Soil Physical and Chemical Properties in Long Term Improved Natural and Traditional Agroforestry Management Systems of Cacao Genotypes in Peruvian Amazon. PLoS ONE 2015, 10, e0132147. [Google Scholar] [CrossRef] [PubMed]
- Crouzet, O.; Poly, F.; Bonnemoy, F.; Bru, D.; Batisson, I.; Bohatier, J.; Philippot, L.; Mallet, C. Functional and Structural Responses of Soil N-Cycling Microbial Communities to the Herbicide Mesotrione: A Dose-Effect Microcosm Approach. Environ. Sci. Pollut. Res. Int. 2016, 23, 4207–4217. [Google Scholar] [CrossRef]
- Pan, Z.; Fan, D.; Jiang, R.; Abbasi, N.; Song, D.; Zou, G.; Wei, D.; He, P.; He, W. Improving Potato Productivity and Mitigating Nitrogen Losses Using Enhanced-Efficiency Fertilizers: A Global Meta-Analysis. Agric. Ecosyst. Environ. 2023, 348, 108416. [Google Scholar] [CrossRef]
- Ma, H.; Xie, C.; Zheng, S.; Li, P.; Cheema, H.N.; Gong, J.; Xiang, Z.; Liu, J.; Qin, J. Potato Tillage Method Is Associated with Soil Microbial Communities, Soil Chemical Properties, and Potato Yield. J. Microbiol. 2022, 60, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Hijri, M. Analysis of a Large Dataset of Mycorrhiza Inoculation Field Trials on Potato Shows Highly Significant Increases in Yield. Mycorrhiza 2016, 26, 209–214. [Google Scholar] [CrossRef]
- Betencourt, E.; Duputel, M.; Colomb, B.; Desclaux, D.; Hinsinger, P. Intercropping Promotes the Ability of Durum Wheat and Chickpea to Increase Rhizosphere Phosphorus Availability in a Low P Soil. Soil Biol. Biochem. 2012, 46, 181–190. [Google Scholar] [CrossRef]
- Cong, W.F.; Hoffland, E.; Li, L.; Six, J.; Sun, J.H.; Bao, X.G.; Zhang, F.S.; Van Der Werf, W. Intercropping Enhances Soil Carbon and Nitrogen. Glob. Change Biol. 2015, 21, 1715–1726. [Google Scholar] [CrossRef]
- Chai, Q.; Nemecek, T.; Liang, C.; Zhao, C.; Yu, A.; Coulter, J.A.; Wang, Y.; Hu, F.; Wang, L.; Siddique, K.H.M.; et al. Integrated Farming with Intercropping Increases Food Production While Reducing Environmental Footprint. Proc. Natl. Acad. Sci. USA 2021, 118, e2106382118. [Google Scholar] [CrossRef]
- Servicio Nacional de Sanidad Agraria Guía para la Implementación de Buenas Prácticas Agrícolas (BPA). Para El Cultivo de Papa; SENASA: Lima, Peru, 2020. [Google Scholar]
- Dubova, L.; Šenberga, A.; Alsiņa, I. The Effect of Double Inoculation on the Broad Beans (Vicia faba L.) Yield Quality. Res. Rural Dev. 2015, 1, 34–39. [Google Scholar]
- Youseif, S.H.; Fayrouz, H.A.E.M.; Saleh, S.A. Improvement of Faba Bean Yield Using Rhizobium/Agrobacterium Inoculant in Low-Fertility Sandy Soil. Agronomy 2017, 7, 2. [Google Scholar] [CrossRef]
- Yin, J.; Sui, Z.; Li, Y.; Yang, H.; Yuan, L.; Huang, J. A New Function of White-Rot Fungi Ceriporia Lacerata HG2011: Improvement of Biological Nitrogen Fixation of Broad Bean (Vicia faba). Microbiol. Res. 2022, 256, 126939. [Google Scholar] [CrossRef]
- Chen, X.; Chen, J.; Cao, J. Intercropping Increases Soil N-Targeting Enzyme Activities: A Meta-Analysis. Rhizosphere 2023, 26, 100686. [Google Scholar] [CrossRef]
- Virk, A.L.; Lin, B.J.; Kan, Z.R.; Qi, J.Y.; Dang, Y.P.; Lal, R.; Zhao, X.; Zhang, H.L. Simultaneous Effects of Legume Cultivation on Carbon and Nitrogen Accumulation in Soil. Adv. Agron. 2022, 171, 75–110. [Google Scholar] [CrossRef]
- Abd-Alla, M.H.; El-Enany, A.W.E.; Nafady, N.A.; Khalaf, D.M.; Morsy, F.M. Synergistic Interaction of Rhizobium Leguminosarum Bv. Viciae and Arbuscular Mycorrhizal Fungi as a Plant Growth Promoting Biofertilizers for Faba Bean (Vicia faba L.) in Alkaline Soil. Microbiol. Res. 2014, 169, 49–58. [Google Scholar] [CrossRef]
- Nicolás, E.; Maestre-Valero, J.F.; Alarcón, J.J.; Pedrero, F.; Vicente-Sánchez, J.; Bernabé, A.; Gómez-Montiel, J.; Hernández, J.A.; Fernández, F. Effectiveness and Persistence of Arbuscular Mycorrhizal Fungi on the Physiology, Nutrient Uptake and Yield of Crimson Seedless Grapevine. J. Agric. Sci. 2015, 153, 1084–1096. [Google Scholar] [CrossRef]
- Smith, S.E.; Smith, F.A. Fresh Perspectives on the Roles of Arbuscular Mycorrhizal Fungi in Plant Nutrition and Growth. Mycologia 2012, 104, 1–13. [Google Scholar] [CrossRef]
- Parniske, M. Arbuscular Mycorrhiza: The Mother of Plant Root Endosymbioses. Nat. Rev. Microbiol. 2008, 6, 763–775. [Google Scholar] [CrossRef]
- Gómez-Bellot, M.J.; Ortuño, M.F.; Álvarez, S.; Sánchez-Blanco, M.J. Influence of Mycorrhizal or Microbial Complex Inoculation on Laurustinus Plants Irrigated with Reclaimed Water. J. Hortic. Sci. Biotechnol. 2020, 95, 661–672. [Google Scholar] [CrossRef]
- Pereira, S.; Mucha, Â.; Gonçalves, B.; Bacelar, E.; Látr, A.; Ferreira, H.; Oliveira, I.; Rosa, E.; Marques, G. Improvement of Some Growth and Yield Parameters of Faba Bean (Vicia faba) by Inoculation with Rhizobium Laguerreae and Arbuscular Mycorrhizal Fungi. Crop Pasture Sci. 2019, 70, 595–605. [Google Scholar] [CrossRef]
- Baum, C.; El-Tohamy, W.; Gruda, N. Increasing the Productivity and Product Quality of Vegetable Crops Using Arbuscular Mycorrhizal Fungi: A Review. Sci. Hortic. 2015, 187, 131–141. [Google Scholar] [CrossRef]
- Cabral, C.; Ravnskov, S.; Tringovska, I.; Wollenweber, B. Arbuscular Mycorrhizal Fungi Modify Nutrient Allocation and Composition in Wheat (Triticum aestivum L.) Subjected to Heat-Stress. Plant Soil 2016, 408, 385–399. [Google Scholar] [CrossRef]
- Jiménez-Martínez, A.; del, C. Gutiérrez-Castorena, M.; Montaño, N.M.; Gutiérrez-Castorena, E.V.; Alarcón, A.; Gavito, M.E. Micromorphology and Thematic Micro-Mapping Reveal Differences in the Soil Structuring Traits of Three Arbuscular Mycorrhizal Fungi. Pedobiologia 2024, 104, 150953. [Google Scholar] [CrossRef]
- Lehmann, A.; Zheng, W.; Rillig, M.C. Soil Biota Contributions to Soil Aggregation. Nat. Ecol. Evol. 2017, 1, 1828–1835. [Google Scholar] [CrossRef]
- Shah, K.K.; Modi, B.; Pandey, H.P.; Subedi, A.; Aryal, G.; Pandey, M.; Shrestha, J. Diversified Crop Rotation: An Approach for Sustainable Agriculture Production. Adv. Agric. 2021, 2021, 8924087. [Google Scholar] [CrossRef]
- Viguier, L.; Cavan, N.; Bockstaller, C.; Cadoux, S.; Corre-Hellou, G.; Dubois, S.; Duval, R.; Keichinger, O.; Toqué, C.; Toupet de Cordoue, A.L.; et al. Combining Diversification Practices to Enhance the Sustainability of Conventional Cropping Systems. Eur. J. Agron. 2021, 127, 126279. [Google Scholar] [CrossRef]
- Castillo, C.; Solano, J.; Collinao, M.; Catalán, R.; Campos, P.; Aguilera, P.; Sieverding, E.; Borie, F. Intercropping Wheat with Ancestral Non-Mycorrhizal Crops in a Volcanic Soil at Early Growth Stage. Chil. J. Agric. Res. 2022, 82, 663–672. [Google Scholar] [CrossRef]
- Meza, K.; Vanek, S.J.; Sueldo, Y.; Olivera, E.; Ccanto, R.; Scurrah, M.; Fonte, S.J. Grass–Legume Mixtures Show Potential to Increase Above-and Belowground Biomass Production for Andean Forage-Based Fallows. Agronomy 2022, 12, 142. [Google Scholar] [CrossRef]
- Li, J.; Lei, Y.; Wen, Y.; Zhu, J.; Di, X.; Zeng, Y.; Han, X.; Que, Z.; Mediatrice, H.; Rensing, C.; et al. Short-Term Effects of Cenchrus Fungigraminus/Potato or Broad Bean Interplanting on Rhizosphere Soil Fertility, Microbial Diversity, and Greenhouse Gas Sequestration in Southeast China. Microorganisms 2024, 12, 1665. [Google Scholar] [CrossRef]
- Taurinanda, A.P.; Banjarnahor, D.R.V. Mycorrhiza Diversity in Some Intercropping Systems of Potato (Solanum tuberosum L.) and Faba Bean (Vicia faba L.). J. Tek. Pertan. Lampung 2023, 12, 495. [Google Scholar] [CrossRef]
- Al-Zubaidi, A.H.A. Biofertilizer Impact on the Productivity of Broad Bean (Vicia faba L.). SABRAO J. Breed. Genet. 2024, 56, 1705–1711. [Google Scholar] [CrossRef]
- Carrara, J.E.; Reddivari, L.; Lehotay, S.J.; Zinati, G.; Heller, W.P. Arbuscular Mycorrhizal Fungi Increase the Yield and Nutritional Quality of Yellow and Purple Fleshed Potatoes (Solanum tuberosum). Am. J. Potato Res. 2023, 100, 210–220. [Google Scholar] [CrossRef]
- Kuila, D.; Ghosh, S. Arbuscular Mycorrhizae and Mycorrhizae Helper Organism---A Synergistic Movement Towards Soil and Crop Sustainability. In Arbuscular Mycorrhizal Fungi in Sustainable Agriculture: Nutrient and Crop Management; Parihar, M., Rakshit, A., Adholeya, A., Chen, Y., Eds.; Springer Nature: Singapore, 2024; pp. 429–451. ISBN 978-981-97-0300-5. [Google Scholar]
- Sharma, U.C.; Datta, M.; Sharma, V. Soil Microbes and Biofertilizers. In Soils in the Hindu Kush Himalayas: Management for Agricultural Land Use; Springer International Publishing: Cham, Switzerland, 2022; pp. 117–144. ISBN 978-3-031-11458-8. [Google Scholar]
- Mahanty, T.; Bhattacharjee, S.; Goswami, M.; Bhattacharyya, P.; Das, B.; Ghosh, A.; Tribedi, P. Biofertilizers: A Potential Approach for Sustainable Agriculture Development. Environ. Sci. Pollut. Res. 2017, 24, 3315–3335. [Google Scholar] [CrossRef] [PubMed]
- Mitter, E.K.; Tosi, M.; Obregón, D.; Dunfield, K.E.; Germida, J.J. Rethinking Crop Nutrition in Times of Modern Microbiology: Innovative Biofertilizer Technologies. Front. Sustain. Food Syst. 2021, 5, 1–23. [Google Scholar] [CrossRef]
- Nosheen, S.; Ajmal, I.; Song, Y. Microbes as Biofertilizers, a Potential Approach for Sustainable Crop Production. Sustainability 2021, 13, 1868. [Google Scholar] [CrossRef]
- Mahdi, S.S.; Hassan, G.I.; Samoon, S.A.; Rather, H.A.; Dar, S.A.; Zehra, B. Bio-Fertilizers In Organic Agriculture. J. Phytol. 2010, 2, 42–54. [Google Scholar]
- Bhat, T.A.; Ahmad, L.; Ganai, M.A.; Shams-Ul-Haq; Khan, O.A. Nitrogen Fixing Biofertilizers; Mechanism and Growth Promotion: A Review. J. Pure Appl. Microbiol. 2015, 9, 1675–1690. [Google Scholar]
- Schütz, L.; Gattinger, A.; Meier, M.; Müller, A.; Boller, T.; Mäder, P.; Mathimaran, N. Improving Crop Yield and Nutrient Use Efficiency via Biofertilization—A Global Meta-Analysis. Front. Plant Sci. 2018, 8. [Google Scholar] [CrossRef]
- Choudhury, A.T.M.A.; Kennedy, I.R. Prospects and Potentials for Systems of Biological Nitrogen Fixation in Sustainable Rice Production. Biol. Fertil. Soils 2004, 39, 219–227. [Google Scholar] [CrossRef]
- Susniak, K.; Krysa, M.; Kidaj, D.; Szymanska-Chargot, M.; Komaniecka, I.; Zamlynska, K.; Choma, A.; Wielbo, J.; Ilag, L.L.; Sroka-Bartnicka, A. Multimodal Spectroscopic Imaging of Pea Root Nodules to Assess the Nitrogen Fixation in the Presence of Biofertilizer Based on Nod-Factors. Int. J. Mol. Sci. 2021, 22, 12991. [Google Scholar] [CrossRef] [PubMed]
- Bocchi, S.; Malgioglio, A. Azolla-Anabaena as a Biofertilizer for Rice Paddy Fields in the Po Valley, a Temperate Rice Area in Northern Italy. Int. J. Agron. 2010, 2010, 152158. [Google Scholar] [CrossRef]
- Setiawati, M.R.; Damayani, M.; Herdiyantoro, D.; Suryatmana, P.; Anggraini, D.; Khumairah, F.H. The Application Dosage of Azolla Pinnata in Fresh and Powder Form as Organic Fertilizer on Soil Chemical Properties, Growth and Yield of Rice Plant. In Proceedings of the AIP Conference Proceedings, Jatinangor, Indonesia, 8–9 August 2017; Volume 1927. [Google Scholar]
- Baset Mia, M.A.; Shamsuddin, Z.H. Rhizobium as a Crop Enhancer and Biofertilizer for Increased Cereal Production. Afr. J. Biotechnol. 2010, 9, 6001–6009. [Google Scholar]
- Revillas, J.J.; Rodelas, B.; Pozo, C.; Martínez-Toledo, M.V.; González López, J. Production of Amino Acids by Azotobacter Vinelandii and Azotobacter Chroococcum with Phenolic Compounds as Sole Carbon Source under Diazotrophic and Adiazotrophic Conditions. Amino Acids 2005, 28, 421–425. [Google Scholar] [CrossRef] [PubMed]
- Kizilkaya, R. Nitrogen Fixation Capacity of Azotobacter spp. Strains Isolated from Soils in Different Ecosystems and Relationship between Them and the Microbiological Properties of Soils. J. Environ. Biol. 2009, 30, 73–82. [Google Scholar]
- Chungopast, S.; Thongjoo, C.; Islam, A.K.M.M.; Yeasmin, S. Efficiency of Phosphate-Solubilizing Bacteria to Address Phosphorus Fixation in Takhli Soil Series: A Case of Sugarcane Cultivation, Thailand. Plant Soil 2021, 460, 347–357. [Google Scholar] [CrossRef]
- Nguyen Quoc, K.; Le Vinh, T.; Le Thanh, Q.; Tran Ngoc, H.; Do Thi, X.; Huynh Huu, D.; Ly Ngoc Thanh, X.; Le Thi My, T. Effects of Biofertilizer Supplementation, Rhodopseudomonas Spp., on Nitrogen and Phosphorus Uptakes, Growth, and Yield of Sesame (Sesamum indicum L.) on Salt-Affected Soil. J. Plant Nutr. 2024, 47, 1–17. [Google Scholar] [CrossRef]
- Singh, Z.; Singh, G.; Aggarwal, N.; Virk, H.K.; Gupta, R.K. Applications of Biofertilizers and Phosphorus Influence Nutrient Uptake and Nutrient Use Efficiency in Chickpea (Cicer arietinum L.). Commun. Soil Sci. Plant Anal. 2024, 55, 1093–1104. [Google Scholar] [CrossRef]
- Seenivasagan, R.; Babalola, O.O. Utilization of Microbial Consortia as Biofertilizers and Biopesticides for the Production of Feasible Agricultural Product. Biology 2021, 10, 1111. [Google Scholar] [CrossRef]
- Anand, K.; Kumari, B.; Mallick, M.A. Phosphate Solubilizing Microbes: An Effective and Alternative Approach as Biofertilizers. Int. J. Pharm. Pharm. Sci. 2016, 8, 37–40. [Google Scholar]
- Din, M.; Nelofer, R.; Salman, M.; Abdullah; Khan, F.H.; Khan, A.; Ahmad, M.; Jalil, F.; Din, J.U.; Khan, M. Production of Nitrogen Fixing Azotobacter (SR-4) and Phosphorus Solubilizing Aspergillus Niger and Their Evaluation on Lagenaria Siceraria and Abelmoschus Esculentus. Biotechnol. Rep. 2019, 22, e00323. [Google Scholar] [CrossRef] [PubMed]
- Sheng, X.F.; He, L.Y. Solubilization of Potassium-Bearing Minerals by a Wild-Type Strain of Bacillus Edaphicus and Its Mutants and Increased Potassium Uptake by Wheat. Can. J. Microbiol. 2006, 52, 66–72. [Google Scholar] [CrossRef]
- Basak, B.B.; Biswas, D.R. Co-Inoculation of Potassium Solubilizing and Nitrogen Fixing Bacteria on Solubilization of Waste Mica and Their Effect on Growth Promotion and Nutrient Acquisition by a Forage Crop. Biol. Fertil. Soils 2010, 46, 641–648. [Google Scholar] [CrossRef]
- Han, H.S.; Supanjani; Lee, K.D. Effect of Co-Inoculation with Phosphate and Potassium. Plant Soil Environ. 2006, 52, 130–136. [Google Scholar] [CrossRef]
- ISO 11265:1994; Soil Quality-Determination of the Specific Electrical Conductivity. ISO: Geneva, Switzerland, 1996.
- USEPA. Method 9045D Soil and Waste PH; USEPA: Washington, DC, USA, 2004; Volume 8, pp. 104–110. [Google Scholar]
- Walkley, A.; Black, I. An Examination of the Degtjareff Method and a Proposed Modification of the Chromic Matter and a Proposed Modification of the Chromic Acid Titration Method. Soil Sci. 1934, 34, 29–38. [Google Scholar] [CrossRef]
- Bray, R.H.; Kurtz, L.T. Determination of Total Organic and Available Forms of Phosphorus in Soils. Soil Sci. 1945, 59, 39–45. [Google Scholar] [CrossRef]
- Secretaría de Medio Ambiente y Recursos Naturales (SEMARNAT). Norma Oficial Mexicana NOM-021-RECNAT-2000. 2002. Available online: http://www.ordenjuridico.gob.mx/Documentos/Federal/wo69255.pdf (accessed on 10 December 2024).
- Bouyoucos, G.J. Directions for Making Mechanical Analysis of Soils by the Hydrometer Method. Soil Sci. 1936, 42, 225–230. [Google Scholar] [CrossRef]
- Tahir, M.; Ahmad, I.; Shahid, M.; Shah, G.M.; Farooq, A.B.U.; Akram, M.; Tabassum, S.A.; Naeem, M.A.; Khalid, U.; Ahmad, S.; et al. Regulation of Antioxidant Production, Ion Uptake and Productivity in Potato (Solanum tuberosum L.) Plant Inoculated with Growth Promoting Salt Tolerant Bacillus Strains. Ecotoxicol. Environ. Saf. 2019, 178, 33–42. [Google Scholar] [CrossRef]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of Arbuscular Mycorrhizal Fungi in Plant Growth Regulation: Implications in Abiotic Stress Tolerance. Front. Plant Sci. 2019, 10, 466052. [Google Scholar] [CrossRef]
- Begum, N.; Wang, L.; Ahmad, H.; Akhtar, K.; Roy, R.; Khan, M.I.; Zhao, T. Co-Inoculation of Arbuscular Mycorrhizal Fungi and the Plant Growth-Promoting Rhizobacteria Improve Growth and Photosynthesis in Tobacco Under Drought Stress by Up-Regulating Antioxidant and Mineral Nutrition Metabolism. Microb. Ecol. 2022, 83, 971–988. [Google Scholar] [CrossRef]
- Karagatzides, J.D.; Wilton, M.J.; Tsuji, L.J.S. Soil Nutrient Supply in Cultivated Bush Bean–Potato Intercropping Grown in Subarctic Soil Managed with Agroforestry. Sustainability 2021, 13, 8185. [Google Scholar] [CrossRef]
- Fernández Martín, F.; Molina, J.J.; Nicolás Nicolás, E.; Alarcón, J.J.; Kirchmair, M.; García, F.J.; Bernabe, A.J.; Bernal, C. Application of Arbuscular Mycorrhizae Glomus Iranicum Var. Tenuihypharum Var. Nova in Intensive Agriculture: A Study Case. J. Agric. Sci. Technol. B 2017, 7, 221–247. [Google Scholar] [CrossRef]
- Duffy, E.M.; Cassells, A.C. The Effect of Inoculation of Potato (Solanum tuberosum L.) Microplants with Arbuscular Mycorrhizal Fungi on Tuber Yield and Tuber Size Distribution. Appl. Soil Ecol. 2000, 15, 137–144. [Google Scholar] [CrossRef]
- Lone, R.; Shuab, R.; Sharma, V.; Kumar, V.; Mir, R.; Koul, K.K. Effect of Arbuscular Mycorrhizal Fungi on Growth and Development of Potato (Solanum tuberosum) Plant. Asian J. Crop Sci. 2015, 7, 233–243. [Google Scholar] [CrossRef]
- Davies, F.T.; Calderón, C.M.; Huaman, Z.; Gómez, R. Influence of a Flavonoid (Formononetin) on Mycorrhizal Activity and Potato Crop Productivity in the Highlands of Peru. Sci. Hortic. 2005, 106, 318–329. [Google Scholar] [CrossRef]
- Cozzolino, V.; Di Meo, V.; Piccolo, A. Impact of Arbuscular Mycorrhizal Fungi Applications on Maize Production and Soil Phosphorus Availability. J. Geochem. Explor. 2013, 129, 40–44. [Google Scholar] [CrossRef]
- Vicente-Sánchez, J.; Nicolás, E.; Pedrero, F.; Alarcón, J.J.; Maestre-Valero, J.F.; Fernández, F. Arbuscular Mycorrhizal Symbiosis Alleviates Detrimental Effects of Saline Reclaimed Water in Lettuce Plants. Mycorrhiza 2014, 24, 339–348. [Google Scholar] [CrossRef]
- Chafai, W.; Bouchentouf, H.; Khalid, A. Comparative Impact of Arbuscular Mycorrhizal Fungi from Moroccan Soils and Commercial Inoculum on Potato Yield and Nutrient Composition. Aust. J. Crop Sci. 2024, 18, 160–166. [Google Scholar] [CrossRef]
- Salim, H.A.; Ali, A.F.; Alsaady, M.H.M.; Saleh, U.N.; Jassim, N.H.; Hamad, A.R.; Attia, J.A.; Darwish, J.J.; Hassan, A.F. Effect of Plant Growth Promoting Rhizobacteria (PGPR) on Growth of Cauliflower (Brassica oleracea L. Var. botrytis). Plant Arch. 2020, 20, 782–786. [Google Scholar]
- Schellenbaum, L.; Berta, G.; Ravolanirina, F.; Tisserant, B.; Gianinazzi, S.; Fitter, A.H. Influence of Endomycorrhizal Infection on Root Morphology in a Micropropagated Woody Plant Species (Vitis vinifera L.). Ann. Bot. 1991, 68, 135–141. [Google Scholar] [CrossRef]
- Hooker, J.E.; Munro, M.; Atkinson, D. Vesicular-Arbuscular Mycorrhizal Fungi Induced Alteration in Poplar Root System Morphology. Plant Soil 1992, 145, 207–214. [Google Scholar] [CrossRef]
- Giri, B.; Kapoor, R.; Mukerji, K.G. Influence of Arbuscular Mycorrhizal Fungi and Salinity on Growth, Biomass, and Mineral Nutrition of Acacia Auriculiformis. Biol. Fertil. Soils 2003, 38, 170–175. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; DeForest, J.L.; Burton, A.J.; Allen, M.F.; Ruess, R.W.; Hendrick, R.L. Fine Root Architecture of Nine North American Trees. Ecol. Monogr. 2002, 72, 293. [Google Scholar] [CrossRef]
- Nurbaity, A.; Hamdani, J.S.; Rahayu, R.P. Effect of Arbuscular Mycorrhizal Fungi and Different Composition of Growing Medium on Growth and Production of Potato Seed Cultivars Medians in Inceptisols Jatinangor. IOP Conf. Ser. Earth Environ. Sci. 2019, 393, 012052. [Google Scholar] [CrossRef]
- Naji, N.S.; Dhiyab, N.S.; Abed, R.M. Efficiency of Some Local Isolates of Arbuscular Mycorrhizae in the Growth and Productivity of Potatoes (Solanum tuberosum L.) in Plastic Pots. Plant Sci. Today 2024, 11, 274–280. [Google Scholar] [CrossRef]
- Kaya, C.; Ashraf, M.; Sonmez, O.; Aydemir, S.; Tuna, A.L.; Cullu, M.A. The Influence of Arbuscular Mycorrhizal Colonisation on Key Growth Parameters and Fruit Yield of Pepper Plants Grown at High Salinity. Sci. Hortic. 2009, 121, 1–6. [Google Scholar] [CrossRef]
- Sheng, M.; Tang, M.; Chen, H.; Yang, B.; Zhang, F.; Huang, Y. Influence of Arbuscular Mycorrhizae on the Root System of Maize Plants under Salt Stress. Can. J. Microbiol. 2009, 55, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Abiala, M.A.; Popoola, O.O.; Olawuyi, O.J.; Oyelude, J.O.; Akanmu, A.O.; Killani, A.S.; Osonubi, O.; Odebode, A.C. Harnessing the Potentials of Vesicular Arbuscular Mycorrhizal (VAM) Fungi to Plant Growth—A Review. Int. J. Pure Appl. Sci. Technol. 2013, 14, 61–79. [Google Scholar]
- Balliu, A.; Sallaku, G.; Rewald, B. AMF Inoculation Enhances Growth and Improves the Nutrient Uptake Rates of Transplanted, Salt-Stressed Tomato Seedlings. Sustainability 2015, 7, 15967–15981. [Google Scholar] [CrossRef]
- Allito, B.B.; Ewusi-Mensah, N.; Logah, V.; Hunegnaw, D.K. Legume-Rhizobium Specificity Effect on Nodulation, Biomass Production and Partitioning of Faba Bean (Vicia faba L.). Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Elliott, A.J.; Daniell, T.J.; Cameron, D.D.; Field, K.J. A Commercial Arbuscular Mycorrhizal Inoculum Increases Root Colonization across Wheat Cultivars but Does Not Increase Assimilation of Mycorrhiza-Acquired Nutrients. Plants People Planet 2021, 3, 588–599. [Google Scholar] [CrossRef]
- Ghobadi, M.; Dehnavi, M.M.; Yadavi, A.R.; Parvizi, K.; Zafari, D. Reduced P Fertilization Improves Fe and Zn Uptake in Potato When Inoculated with AMF in P, Fe and Zn Deficient Soil. Rhizosphere 2020, 15, 100239. [Google Scholar] [CrossRef]
- Miceli, A.; Vetrano, F.; Torta, L.; Esposito, A.; Moncada, A. Effect of Mycorrhizal Inoculation on Melon Plants under Deficit Irrigation Regimes. Agronomy 2023, 13, 440. [Google Scholar] [CrossRef]
- Mohammadi, S.A.; Prasanna, B.M. Analysis of Genetic Diversity in Crop Plants—Salient Statistical Tools and Considerations. Crop Sci. 2003, 43, 1235–1248. [Google Scholar] [CrossRef]
- Hanoon, M.B.; Haran, M.S.; Sahi, M.K. Effect of Rhizobium Inoculation and Different Levels of Organic and Nitrogen Fertilizers on Growth and Production of Broad Bean (Vicia faba L.) and Nitrogen Readiness in Soil. Int. J. Agric. Stat. Sci. 2020, 16, 229–236. [Google Scholar]
- Wang, X.; Feng, H.; Wang, Y.; Wang, M.; Xie, X.; Chang, H.; Wang, L.; Qu, J.; Sun, K.; He, W.; et al. Mycorrhizal Symbiosis Modulates the Rhizosphere Microbiota to Promote Rhizobia–Legume Symbiosis. Mol. Plant 2021, 14, 503–516. [Google Scholar] [CrossRef]
- He, J.; Zhang, L.; Van Dingenen, J.; Desmet, S.; Goormachtig, S.; Calonne-Salmon, M.; Declerck, S. Arbuscular Mycorrhizal Hyphae Facilitate Rhizobia Dispersal and Nodulation in Legumes. ISME J. 2024, 18. [Google Scholar] [CrossRef]
- Mateo, N.; Tapia, M. High Mountain Environment and Farming Systems in the Andean Region of Latin America. In Proceedings of the International Workshop on Mountain Crops and Genetic Resources, Katmandu, Nepal, 16–19 February 1987; p. 51. [Google Scholar]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).