
Evolution and Functional Diversity of GATA Transcription Factors in Filamentous Fungi: Structural Characteristics, Metabolic Regulation and Environmental Response

Abstract
1. Introduction
2. Structures and Types of GATA TFs in Filamentous Fungi
2.1. Structural Characterization of Filamentous Fungi GATA TFs
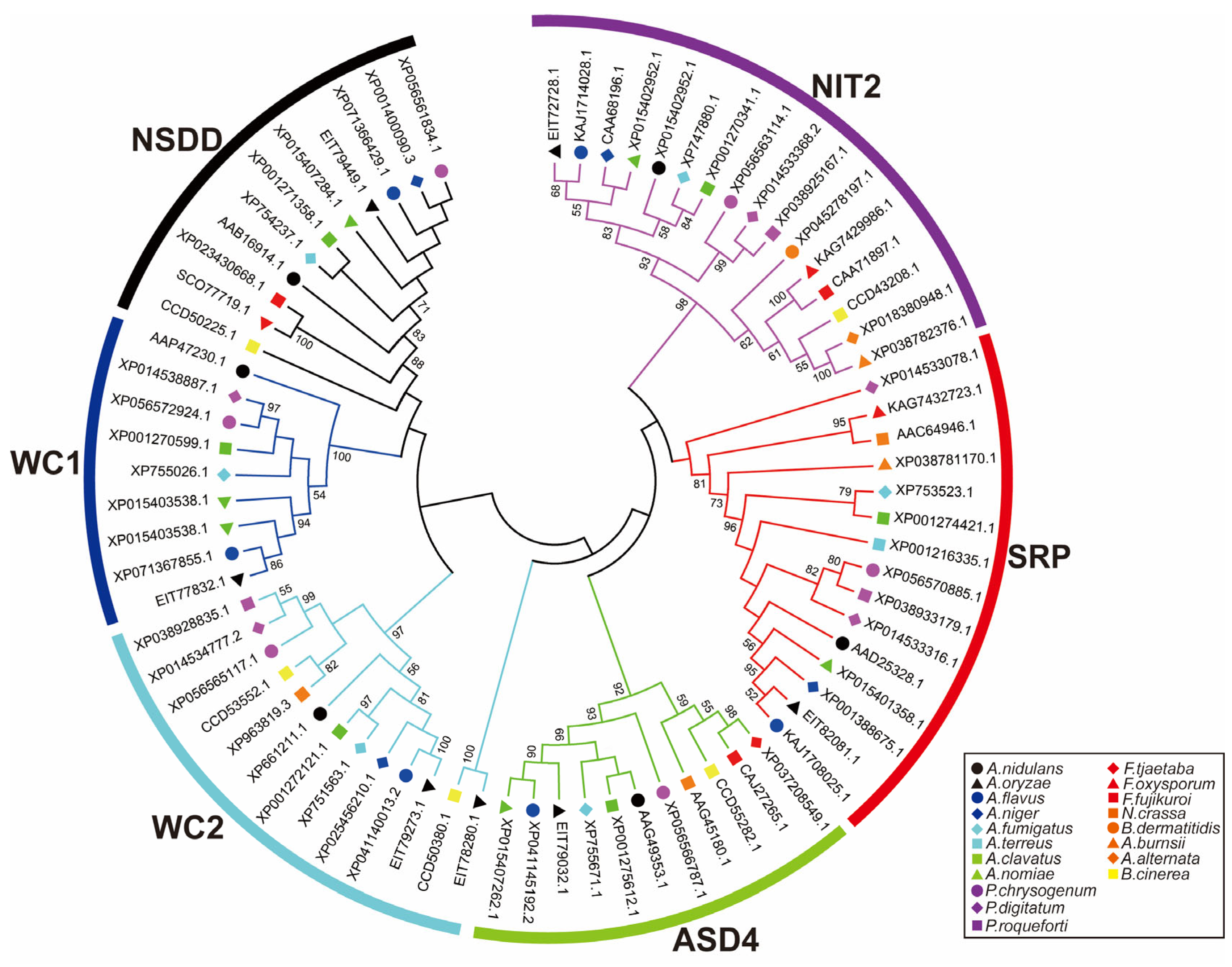
2.2. Types of Filamentous Fungal GATA TFs
3. Biological Function of GATA TFs in Filamentous Fungi
3.1. The Effect of GATA TFs on the Growth and Development of Filamentous Fungi
3.2. Regulatory Roles of GATA TFs in the Secondary Metabolite Biosynthesis of Filamentous Fungi
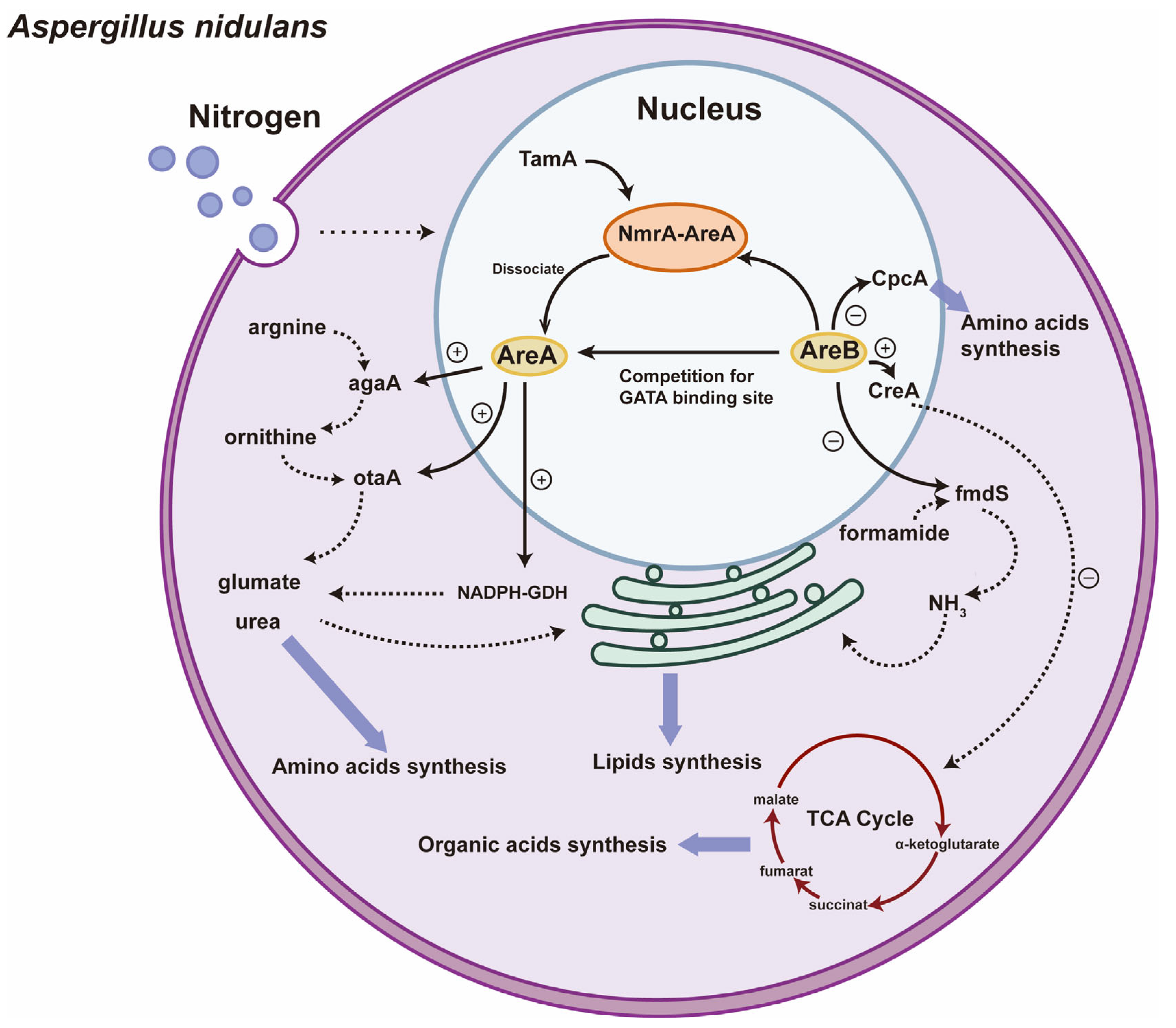
3.3. The Regulatory Roles of GATA TFs in Nitrogen Metabolism and Lipid Synthesis of Filamentous Fungi
3.4. The Roles of GATA TFs in Response to Stress in Filamentous Fungi
4. Conclusions and Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Kerkaert, J.D.; Huberman, L.B. Regulation of nutrient utilization in filamentous fungi. Appl. Microbiol. Biotechnol. 2023, 107, 5873–5898. [Google Scholar] [CrossRef]
- Ghimire, P.S.; Jin, C. Genetics, molecular, and proteomics advances in filamentous fungi. Curr. Microbiol. 2017, 74, 1226–1236. [Google Scholar] [CrossRef]
- Lorenzini, M.; Cappello, M.S.; Logrieco, A.; Zapparoli, G. Polymorphism and phylogenetic species delimitation in filamentous fungi from predominant mycobiota in withered grapes. Int. J. Food Microbiol. 2016, 238, 56–62. [Google Scholar] [CrossRef]
- Orban, A.; Fraatz, M.A.; Rühl, M. Aroma profile analyses of filamentous fungi cultivated on solid substrates. Adv. Biochem. Eng. Biotechnol. 2019, 169, 85–107. [Google Scholar]
- Adav, S.S.; Ravindran, A.; Sze, S.K. Quantitative proteomic study of Aspergillus fumigatus secretome revealed deamidation of secretory enzymes. J. Proteom. 2015, 119, 154–168. [Google Scholar] [CrossRef]
- Adav, S.S.; Li, A.A.; Manavalan, A.; Punt, P.; Sze, S.K. Quantitative iTRAQ secretome analysis of Aspergillus niger reveals novel hydrolytic enzymes. J. Proteome Res. 2010, 9, 3932–3940. [Google Scholar] [CrossRef]
- Kawasaki, L.; Wysong, D.; Diamond, R.; Aguirre, J. Two divergent catalase genes are differentially regulated during Aspergillus nidulans development and oxidative stress. J. Bacteriol. 1997, 179, 3284–3292. [Google Scholar] [CrossRef]
- Appels, F.V.W.; Dijksterhuis, J.; Lukasiewicz, C.E.; Jansen, K.M.; Wösten, H.A.B.; Krijgsheld, P. Hydrophobin gene deletion and environmental growth conditions impact mechanical properties of mycelium by affecting the density of the material. Sci. Rep. 2018, 8, 4703. [Google Scholar] [CrossRef]
- Wösten, H.A.B. Filamentous fungi for the production of enzymes, chemicals, and materials. Curr. Opin. Biotechnol. 2019, 59, 65–70. [Google Scholar] [CrossRef]
- Saykhedkar, S.; Ray, A.; Ayoubi-Canaan, P.; Hartson, S.D.; Prade, R.; Mort, A.J. A time course analysis of the extracellular proteome of Aspergillus nidulans growing on sorghum stover. Biotechnol. Biofuels 2012, 5, 52. [Google Scholar] [CrossRef]
- He, B.; Tu, Y.; Jiang, C.; Zhang, Z.; Li, Y.; Zeng, B. Functional genomics of Aspergillus oryzae: Strategies and progress. Microorganisms 2019, 7, 103. [Google Scholar] [CrossRef]
- Sonne, M.; Jawetz, E. Comparison of the action of ampicillin and benzylpenicillin on enterococci in vitro. Appl. Microbiol. 1968, 16, 645–648. [Google Scholar] [CrossRef]
- Wu, X.L.; Yang, Y.J.; Li, S.X.; Huang, L.; Liang, F. Clinical effects of different penicillin administration regimens in the treatment of group B Streptococcus infection in pregnant women. Drug Eval. 2023, 20, 1542–1545. [Google Scholar]
- Dutta, A.K.; Phull, P.S. Treatment of Helicobacter pylori infection in the presence of penicillin allergy. World J. Gastroenterol. 2021, 27, 7661–7668. [Google Scholar] [CrossRef]
- Van Den Berg, M.; Gidijala, L.; Kiela, J.; Bovenberg, R.; Vander Keli, I. Biosynthesis of active pharmaceuticals: β-lactam biosynthesis in filamentous fungi. Biotechnol. Genet. Eng. Rev. 2010, 27, 1–32. [Google Scholar]
- Liu, L.; Chen, Z.; Liu, W.; Ke, X.; Tian, X.; Chu, J. Cephalosporin C biosynthesis and fermentation in Acremonium chrysogenum. Appl. Microbiol. Biotechnol. 2022, 106, 6413–6426. [Google Scholar] [CrossRef]
- Majeed, S.; Abdullah, M.S.; Dash, G.K.; Ansari, M.T.; Nanda, A. Biochemical synthesis of silver nanoparticles using filamentous fungi Penicillium decumbens (MTCC-2494) and its efficacy against A-549 lung cancer cell line. J. Nanomater. 2016, 2016, 2184134. [Google Scholar]
- Perez-Cuesta, U.; Aparicio-Fernandez, L.; Guruceaga, X.; Martin-Souto, L.; Abad-Diaz-de-Cerio, A.; Antoran, A.; Buldain, I.; Hernando, F.L.; Ramirez-Garcia, A.; Rementeria, A. Melanin and pyomelanin in Aspergillus fumigatus: From its genetics to host interaction. Int. Microbiol. 2020, 23, 55–63. [Google Scholar] [CrossRef]
- Chamilos, G.; Carvalho, A. Aspergillus fumigatus DHN-Melanin. Curr. Top. Microbiol. Immunol. 2020, 425, 17–28. [Google Scholar]
- Pfannmuller, A.; Leufken, J.; Studt, L.; Michielse, C.B.; Sieber, C.M.K.; Guldener, U.; Hawat, S.; Hippler, M.; Fufezan, C.; Tudzynski, B. Comparative transcriptome and proteome analysis reveals a global impact of the nitrogen regulators AreA and AreB on secondary metabolism in Fusarium fujikuroi. PLoS ONE 2017, 12, e0176194. [Google Scholar] [CrossRef]
- Hedden, P.; Sponsel, V. A Century of Gibberellin Research. J. Plant Growth Regul. 2015, 34, 740–760. [Google Scholar] [CrossRef] [PubMed]
- Fasoyin, O.E.; Yang, K.; Qiu, M.; Wang, B.; Wang, S.; Wang, S. Regulation of Morphology, Aflatoxin Production, and Virulence of Aspergillus flavus by the Major Nitrogen Regulatory Gene areA. Toxins 2019, 11, 718. [Google Scholar] [CrossRef]
- Caceres, I.; Khoury, A.A.; Khoury, R.E.; Lorber, S.; Oswald, I.P.; Khoury, A.E.; Atoui, A.; Puel, O.; Bailly, J.D. Aflatoxin Biosynthesis and Genetic Regulation: A Review. Toxins 2020, 12, 150. [Google Scholar] [CrossRef]
- Lin, H.; Cheng, W.; Ding, H.T.; Chen, X.J.; Zhou, Q.F.; Zhao, Y.H. Direct Microbial Conversion of Wheat Straw into Lipid by a Cellulolytic Fungus of Aspergillus oryzae A-4 in Solid-State Fermentation. Bioresour. Technol. 2010, 101, 7556–7562. [Google Scholar] [PubMed]
- Beopoulos, A.; Cescut, J.; Haddouche, R.; Uribelarrea, J.L.; Molina-Jouve, C.; Nicaud, J.M. Yarrowia lipolytica as a Model for Bio-Oil Production. Prog. Lipid Res. 2009, 48, 375–387. [Google Scholar] [CrossRef]
- Fazili, A.B.A.; Shah, A.M.; Zan, X.; Naz, T.; Nosheen, S.; Nazir, Y.; Ullah, S.; Zhang, H.; Song, Y. Mucor circinelloides: A Model Organism for Oleaginous Fungi and Its Potential Applications in Bioactive Lipid Production. Microb. Cell Factories 2022, 21, 29. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, Y.; Zhang, S.; Liu, Q.; Dang, W.; Song, Y. Lipid Accumulation and SNF1 Transcriptional Analysis of Mucor circinelloides on Xylose under Nitrogen Limitation. Antonie Van Leeuwenhoek 2023, 116, 383–391. [Google Scholar] [CrossRef]
- Macios, M.; Caddick, M.X.; Weglenski, P.; Scazzocchio, C.; Dzikowska, A. The GATA Factors AREA and AREB Together with the Co-Repressor NMRA, Negatively Regulate Arginine Catabolism in Aspergillus nidulans in Response to Nitrogen and Carbon Source. Fungal Genet. Biol. 2012, 49, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Christensen, T.; Hynes, M.J.; Davis, M.A. Role of the Regulatory Gene areA of Aspergillus oryzae in Nitrogen Metabolism. Appl. Environ. Microbiol. 1998, 64, 3232–3237. [Google Scholar] [CrossRef]
- Chen, Y.; Cao, Y.; Gai, Y.; Ma, H.; Zhu, Z.; Chung, K.R.; Li, H. Genome-Wide Identification and Functional Characterization of GATA Transcription Factor Gene Family in Alternaria alternata. J. Fungi 2021, 7, 1013. [Google Scholar] [CrossRef]
- Michielse, C.B.; Pfannmüller, A.; Macios, M.; Rengers, P.; Dzikowska, A.; Tudzynski, B. The Interplay between the GATA Transcription Factors AreA, the Global Nitrogen Regulator and AreB in Fusarium fujikuroi. Mol. Microbiol. 2014, 91, 472–493. [Google Scholar] [CrossRef]
- Haas, H.; Zadra, I.; Stöffler, G.; Angermayr, K. The Aspergillus nidulans GATA Factor SREA is Involved in Regulation of Siderophore Biosynthesis and Control of Iron Uptake. J. Biol. Chem. 1999, 274, 4613–4619. [Google Scholar] [CrossRef]
- Oberegger, H.; Schoeser, M.; Zadra, I.; Schrettl, M.; Parson, W.; Haas, H. Regulation of freA, acoA, lysF, and cycA Expression by Iron Availability in Aspergillus nidulans. Appl. Environ. Microbiol. 2002, 68, 5769–5772. [Google Scholar] [CrossRef]
- Franken, A.C.; Lechner, B.E.; Werner, E.R.; Haas, H.; Lokman, B.C.; Ram, A.F.; van den Hondel, C.A.; de Weert, S.; Punt, P.J. Genome Mining and Functional Genomics for Siderophore Production in Aspergillus niger. Brief. Funct. Genom. 2014, 13, 482–492. [Google Scholar] [CrossRef] [PubMed]
- Yap, A.; Volz, R.; Paul, S.; Moye-Rowley, W.S.; Haas, H. Regulation of high-affinity iron acquisition, including acquisition mediated by the iron permease FtrA, is coordinated by AtrR, SrbA, and SreA in Aspergillus fumigatus. mBio 2023, 14, e0075723. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.; Han, K.H.; Yu, J.H. Upstream Regulation of Development and Secondary Metabolism in Aspergillus Species. Cells 2022, 12, 2. [Google Scholar] [CrossRef] [PubMed]
- Purschwitz, J.; Müller, S.; Kastner, C.; Schöser, M.; Haas, H.; Espeso, E.A.; Atoui, A.; Calvo, A.M.; Fischer, R. Functional and Physical Interaction of Blue- and Red-Light Sensors in Aspergillus nidulans. Curr. Biol. 2008, 18, 255–259. [Google Scholar] [CrossRef]
- Jiang, C.; Lv, G.; Ge, J.; He, B.; Zhang, Z.; Hu, Z.; Zeng, B. Genome-Wide Identification of the GATA Transcription Factor Family and Their Expression Patterns under Temperature and Salt Stress in Aspergillus oryzae. AMB Express 2021, 11, 56. [Google Scholar] [CrossRef]
- Fuller, K.K.; Ringelberg, C.S.; Loros, J.J.; Dunlap, J.C. The Fungal Pathogen Aspergillus fumigatus Regulates Growth, Metabolism, and Stress Resistance in Response to Light. mBio 2013, 4, e00142-13. [Google Scholar] [CrossRef]
- Teakle, G.R.; Gilmartin, P.M. Two Forms of Type IV Zinc-Finger Motif and Their Kingdom-Specific Distribution between the Flora, Fauna and Fungi. Trends Biochem. Sci. 1998, 23, 100–102. [Google Scholar] [CrossRef]
- Scazzocchio, C. The fungal GATA factors. Curr. Opin. Microbiol. 2000, 3, 126–131. [Google Scholar] [CrossRef]
- Yu, M.; Yu, J.; Cao, H.; Yong, M.; Liu, Y. Genome-wide identification and analysis of the GATA transcription factor gene family in Ustilaginoidea virens. Genome 2019, 62, 807–816. [Google Scholar] [CrossRef]
- Ronsmans, A.; Wery, M.; Szachnowski, U.; Gautier, C.; Descrimes, M.; Dubois, E.; Morillon, A.; Georis, I. Transcription-dependent spreading of the Dal80 yeast GATA factor across the body of highly expressed genes. PLoS Genet. 2019, 15, e1007999. [Google Scholar] [CrossRef]
- Monahan, B.J.; Askin, M.C.; Hynes, M.J.; Davis, M.A. Differential expression of Aspergillus nidulans ammonium permease genes is regulated by GATA transcription factor AreA. Eukaryot. Cell 2006, 5, 226–237. [Google Scholar] [CrossRef]
- Wu, X.; Zhou, B.; Yin, C.; Guo, Y.; Lin, Y.; Pan, L.; Wang, B. Characterization of natural antisense transcript, sclerotia development and secondary metabolism by strand-specific RNA sequencing of Aspergillus flavus. PLoS ONE 2014, 9, e1007999. [Google Scholar] [CrossRef]
- Conlon, H.; Zadra, I.; Haas, H.; Arst, H.N., Jr.; Jones, M.G.; Caddick, M.X. The Aspergillus nidulans GATA transcription factor gene areB encodes at least three proteins and features three classes of mutation. Mol. Microbiol. 2001, 40, 361–375. [Google Scholar] [CrossRef]
- Haas, H.; Marzluf, G.A. NRE, the major nitrogen regulatory protein of Penicillium chrysogenum, binds specifically to elements in the intergenic promoter regions of nitrate assimilation and penicillin biosynthetic gene clusters. Curr. Genet. 1995, 28, 177–183. [Google Scholar] [CrossRef]
- Haas, H.; Angermayr, K.; Stöffler, G. Molecular analysis of a Penicillium chrysogenum GATA factor encoding gene (sreP) exhibiting significant homology to the Ustilago maydis urbs1 gene. Gene 1997, 184, 33–37. [Google Scholar] [CrossRef]
- Niehaus, E.M.; Schumacher, J.; Burkhardt, I.; Rabe, P.; Spitzer, E.; Münsterkötter, M.; Güldener, U.; Sieber, C.M.K.; Dickschat, J.S.; Tudzynski, B. The GATA-Type Transcription Factor Csm1 Regulates Conidiation and Secondary Metabolism in Fusarium fujikuroi. Front. Microbiol. 2017, 8, 1175. [Google Scholar] [CrossRef]
- López-Berges, M.S.; Schäfer, K.; Hera, C.; Di Pietro, A. Combinatorial function of velvet and AreA in transcriptional regulation of nitrate utilization and secondary metabolism. Fungal Genet. Biol. 2014, 62, 78–84. [Google Scholar] [CrossRef]
- Harrison, K.A.; Marzluf, G.A. Characterization of DNA binding and the cysteine rich region of SRE, a GATA factor in Neurospora crassa involved in siderophore synthesis. Biochemistry 2002, 41, 15288–15295. [Google Scholar] [CrossRef]
- Fu, Y.H.; Marzluf, G.A. Characterization of nit-2, the major nitrogen regulatory gene of Neurospora crassa. Mol. Cell. Biol. 1987, 7, 1691–1696. [Google Scholar] [CrossRef]
- Wong, K.H.; Hynes, M.J.; Todd, R.B.; Davis, M.A. Deletion and Overexpression of the Aspergillus nidulans GATA Factor AreB Reveals Unexpected Pleiotropy. Microbiology 2009, 155, 3868–3880. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.; Haas, H.; Marzluf, G.A. ASD4, a new GATA factor of Neurospora crassa, displays sequence-specific DNA binding and functions in ascus and ascospore development. Biochemistry 2000, 39, 11065–11073. [Google Scholar] [CrossRef] [PubMed]
- Brenna, A.; Talora, C. WC-1 and the proximal GATA sequence mediate a cis-/trans-acting repressive regulation of light-dependent gene transcription in the dark. Int. J. Mol. Sci. 2019, 20, 2854. [Google Scholar] [CrossRef]
- Ren, W.; Qian, C.; Ren, D.; Cai, Y.; Deng, Z.; Zhang, N.; Wang, C.; Wang, Y.; Zhu, P.; Xu, L. The GATA transcription factor BcWCL2 regulates citric acid secretion to maintain redox homeostasis and full virulence in Botrytis cinerea. mBio 2024, 15, e0013324. [Google Scholar] [CrossRef]
- Schumacher, J.; Simon, A.; Cohrs, K.C.; Viaud, M.; Tudzynski, P. The transcription factor BcLTF1 regulates virulence and light responses in the necrotrophic plant pathogen Botrytis cinerea. PLoS Genet. 2014, 10, e1004040. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, G.M.; Sullivan, T.D.; Gallardo, S.S.; Brandhorst, T.T.; Wymelenberg, A.J.V.; Cuomo, C.A.; Suen, G.; Currie, C.R.; Klein, B.S. SREB, a GATA transcription factor that directs disparate fates in Blastomyces dermatitidis including morphogenesis and siderophore biosynthesis. PLoS Pathog. 2010, 6, e1000846. [Google Scholar] [CrossRef]
- Lee, M.K.; Kwon, N.J.; Lee, I.S.; Jung, S.; Kim, S.C.; Yu, J.H. Negative regulation and developmental competence in Aspergillus. Sci. Rep. 2016, 6, 28874. [Google Scholar] [CrossRef]
- Szewczyk, E.; Krappmann, S. Conserved regulators of mating are essential for Aspergillus fumigatus cleistothecium formation. Eukaryot. Cell 2010, 9, 774–783. [Google Scholar] [CrossRef]
- Shen, L.; Gaslonde, T.; Roullier, C.; Wang, H.; Bodin, J.; Porée, F.H.; Ruprich-Robert, G.; Chapeland-Leclerc, F. Functional Characterization of the GATA-Type Transcription Factor PaNsdD in the Filamentous Fungus Podospora anserina and Its Interplay with the Sterigmatocystin Pathway. Appl. Environ. Microbiol. 2022, 88, e0237821. [Google Scholar] [CrossRef] [PubMed]
- He, X.Y. Functional Study of GATA Transcription Factors SsAREA and SsSRE in Sphaerotheca fuliginea. Ph.D. Thesis, Jilin University, Changchun, China, 2021. [Google Scholar]
- Wei, S.; Zhang, Y.; Wu, M.; Lv, Y.; Zhang, S.; Zhai, H.; Hu, Y. Mechanisms of methyl 2-methylbutyrate suppression on Aspergillus flavus growth and aflatoxin B1 biosynthesis. Int. J. Food Microbiol. 2024, 409, 110462. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Pan, Y.; Liu, G. Disruption of the nitrogen regulatory gene AcareA in Acremonium chrysogenum leads to reduction of cephalosporin production and repression of nitrogen metabolism. Fungal Genet. Biol. 2013, 61, 69–79. [Google Scholar] [CrossRef]
- Tudzynski, B.; Homann, V.; Feng, B.; Marzluf, G.A. Isolation, characterization and disruption of the areA nitrogen regulatory gene of Gibberella fujikuroi. Mol. Genet. Genom. 1999, 261, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ke, X.; Jia, R.; Huang, L.; Liu, Z.; Zheng, Y. Gibberellic acid overproduction in Fusarium fujikuroi using regulatory modification and transcription analysis. Appl. Microbiol. Biotechnol. 2023, 107, 3071–3084. [Google Scholar] [CrossRef]
- Wang, L.; Wang, C.; Xu, L.; Wang, M. Regulation of nitrogen utilization and mycotoxin biosynthesis by the GATA transcription factor AaAreA in Alternaria alternata. World J. Microbiol. Biotechnol. 2024, 40, 236. [Google Scholar] [CrossRef]
- Zhu, J.; Sun, Z.; Shi, D.; Song, S.; Lian, L.; Shi, L.; Ren, A.; Yu, H.; Zhao, M. Dual functions of AreA, a GATA transcription factor, on influencing ganoderic acid biosynthesis in Ganoderma lucidum. Environ. Microbiol. 2019, 21, 4166–4179. [Google Scholar] [CrossRef]
- Niehaus, E.M.; von Bargen, K.W.; Espino, J.J.; Pfannmüller, A.; Humpf, H.U.; Tudzynski, B. Characterization of the fusaric acid gene cluster in Fusarium fujikuroi. Appl. Microbiol. Biotechnol. 2014, 98, 1749–1762. [Google Scholar] [CrossRef]
- Schrettl, M.; Kim, H.S.; Eisendle, M.; Kragl, C.; Nierman, W.C.; Heinekamp, T.; Werner, E.R.; Jacobsen, I.; Illmer, P.; Yi, H.; et al. SreA-mediated iron regulation in Aspergillus fumigatus. Mol. Microbiol. 2008, 70, 27–43. [Google Scholar] [CrossRef]
- Yang, Y.; Heidari, F.; Hu, B. Fungi (Mold)-Based Lipid Production. Methods Mol. Biol. 2019, 1995, 51–89. [Google Scholar]
- Hassane, A.M.A.; Eldiehy, K.S.H.; Saha, D.; Mohamed, H.; Mosa, M.A.; Abouelela, M.E.; Abo-Dahab, N.F.; El-Shanawany, A.A. Oleaginous fungi: A promising source of biofuels and nutraceuticals with enhanced lipid production strategies. Arch. Microbiol. 2024, 206, 338. [Google Scholar] [CrossRef] [PubMed]
- Marzluf, G.A. Genetic regulation of nitrogen metabolism in the fungi. Microbiol. Mol. Biol. Rev. 1997, 61, 17–32. [Google Scholar] [PubMed]
- Sun, X.; Chen, Y.Y.; Wu, Y.N.; Chen, B.; Xi, C.H.; Jiang, L.G. Method for Preparing Sphingolipids Using Olive Oil Fermentation. Chinese Patent CN201610087770.3, 11 June 2021. [Google Scholar]
- Dzurendova, S.; Zimmermann, B.; Tafintseva, V.; Kohler, A.; Ekeberg, D.; Shapaval, V. The influence of phosphorus source and the nature of nitrogen substrate on the biomass production and lipid accumulation in oleaginous Mucoromycota fungi. Appl. Microbiol. Biotechnol. 2020, 104, 8065–8076. [Google Scholar] [CrossRef]
- Aguilar, L.R.; Pardo, J.P.; Lomelí, M.M.; Bocardo, O.I.L.; Juárez Oropeza, M.A.; Guerra Sánchez, G. Lipid droplets accumulation and other biochemical changes induced in the fungal pathogen Ustilago maydis under nitrogen-starvation. Arch. Microbiol. 2017, 199, 1195–1209. [Google Scholar] [CrossRef]
- Maltsev, Y.; Kulikovskiy, M.; Maltseva, S. Nitrogen and phosphorus stress as a tool to induce lipid production in microalgae. Microb. Cell Factories 2023, 22, 239. [Google Scholar] [CrossRef] [PubMed]
- Ran, Y.; Xu, H.; Yang, Q.; Xu, Y.; Yang, H.; Qiao, D.; Cao, Y. GATA-type transcriptional factor SpGAT1 interacts with SpMIG1 and promotes lipid accumulation in the oleaginous yeast Saitozyma podzolica zwy-2-3. Biotechnol. Biofuels Bioprod. 2022, 15, 103. [Google Scholar] [CrossRef] [PubMed]
- Pomraning, K.R.; Bredeweg, E.L.; Baker, S.E. Regulation of nitrogen metabolism by GATA zinc finger transcription factors in Yarrowia lipolytica. mSphere 2017, 2, e00038-17. [Google Scholar] [CrossRef]
- Javelle, A.; Morel, M.; Rodríguez-Pastrana, B.R.; Botton, B.; André, B.; Marini, A.M.; Brun, A.; Chalot, M. Molecular characterization, function, and regulation of ammonium transporters (Amt) and ammonium-metabolizing enzymes (GS, NADP-GDH) in the ectomycorrhizal fungus Hebeloma cylindrosporum. Mol. Microbiol. 2003, 47, 411–430. [Google Scholar] [CrossRef]
- Breuninger, M.; Trujillo, C.G.; Serrano, E.; Fischer, R.; Requena, N. Different nitrogen sources modulate activity but not expression of glutamine synthetase in arbuscular mycorrhizal fungi. Fungal Genet. Biol. 2004, 41, 542–552. [Google Scholar] [CrossRef]
- Small, A.J.; Hynes, M.J.; Davis, M.A. The TamA protein fused to a DNA-binding domain can recruit AreA, the major nitrogen regulatory protein, to activate gene expression in Aspergillus nidulans. Genetics 1999, 153, 95–105. [Google Scholar] [CrossRef]
- Khandelwal, R.; Srivastava, P.; Bisaria, V.S. Recent advances in the production of malic acid by native fungi and engineered microbes. World J. Microbiol. Biotechnol. 2023, 39, 217. [Google Scholar] [CrossRef] [PubMed]
- Lamb, H.K.; Ren, J.; Park, A.; Johnson, C.; Leslie, K.; Cocklin, S.; Thompson, P.; Mee, C.; Cooper, A.; Stammers, D.K.; et al. Modulation of the ligand binding properties of the transcription repressor NmrA by GATA-containing DNA and site-directed mutagenesis. Protein Sci. 2004, 13, 3127–3138. [Google Scholar] [CrossRef] [PubMed]
- Chudzicka-Ormaniec, P.; Macios, M.; Koper, M.; Weedall, G.D.; Caddick, M.X.; Weglenski, P.; Dzikowska, A. The role of the GATA transcription factor AreB in regulation of nitrogen and carbon metabolism in Aspergillus nidulans. FEMS Microbiol. Lett. 2019, 366, fnz066. [Google Scholar] [CrossRef] [PubMed]
- Kuruvilla, F.G.; Shamji, A.F.; Schreiber, S.L. Carbon- and nitrogen-quality signaling to translation are mediated by distinct GATA-type transcription factors. Proc. Natl. Acad. Sci. USA 2001, 98, 7283–7288. [Google Scholar] [CrossRef]
- Strauss, J.; Horvath, H.K.; Abdallah, B.M.; Kindermann, J.; Mach, R.L.; Kubicek, C.P. The function of CreA, the carbon catabolite repressor of Aspergillus nidulans, is regulated at the transcriptional and post-transcriptional level. Mol. Microbiol. 1999, 32, 169–178. [Google Scholar] [CrossRef]
- Svetlov, V.V.; Cooper, T.G. The Saccharomyces cerevisiae GATA factors Dal80p and Deh1p can form homo- and heterodimeric complexes. J. Bacteriol. 1998, 180, 5682–5688. [Google Scholar] [CrossRef]
- Dagenais, T.R.; Keller, N.P. Pathogenesis of Aspergillus fumigatus in Invasive Aspergillosis. Clin. Microbiol. Rev. 2009, 22, 447–465. [Google Scholar] [CrossRef]
- Boyce, K.J.; Andrianopoulos, A. Fungal dimorphism: The switch from hyphae to yeast is a specialized morphogenetic adaptation allowing colonization of a host. FEMS Microbiol. Rev. 2015, 39, 797–811. [Google Scholar] [CrossRef]
- Romani, L. Immunity to fungal infections. Nat. Rev. Immunol. 2004, 4, 11–24. [Google Scholar] [CrossRef]
- Marty, A.J.; Broman, A.T.; Zarnowski, R.; Dwyer, T.G.; Bond, L.M.; Lounes-Hadj Sahraoui, A.; Fontaine, J.; Ntambi, J.M.; Keleş, S.; Kendziorski, C.; et al. Fungal Morphology, Iron Homeostasis, and Lipid Metabolism Regulated by a GATA Transcription Factor in Blastomyces dermatitidis. PLoS Pathog. 2015, 11, e1004959. [Google Scholar] [CrossRef]
- Bugeja, H.E.; Hynes, M.J.; Andrianopoulos, A. AreA controls nitrogen source utilization during both growth programs of the dimorphic fungus Penicillium marneffei. Fungal Biol. 2012, 116, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Crespo, J.L.; Daicho, K.; Ushimaru, T.; Hall, M.N. The GATA transcription factors GLN3 and GAT1 link TOR to salt stress in Saccharomyces cerevisiae. J. Biol. Chem. 2001, 276, 34441–34444. [Google Scholar] [CrossRef]
- García-Salcedo, R.; Casamayor, A.; Ruiz, A.; González, A.; Prista, C.; Loureiro-Dias, M.C.; Ramos, J.; Ariño, J. Heterologous expression implicates a GATA factor in regulation of nitrogen metabolic genes and ion homeostasis in the halotolerant yeast Debaryomyces hansenii. Eukaryot. Cell 2006, 5, 1388–1398. [Google Scholar] [CrossRef]
- Schumacher, J. How light affects the life of Botrytis. Fungal Genet. Biol. 2017, 106, 26–41. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, G.; Deng, W.; Li, T. Distribution, evolution and expression of GATA-TFs provide new insights into their functions in light response and fruiting body development of Tolypocladium guangdongense. PeerJ 2020, 8, e9784. [Google Scholar] [CrossRef] [PubMed]
- Flores-Ibarra, A.; Maia, R.N.A.; Olasz, B.; Church, J.R.; Gotthard, G.; Schapiro, I.; Heberle, J.; Nogly, P. Light-Oxygen-Voltage (LOV)-sensing Domains: Activation Mechanism and Optogenetic Stimulation. J. Mol. Biol. 2024, 436, 168356. [Google Scholar] [CrossRef]
- Ballario, P.; Talora, C.; Galli, D.; Linden, H.; Macino, G. Roles in dimerization and blue light photoresponse of the PAS and LOV domains of Neurospora crassa white collar proteins. Mol. Microbiol. 1998, 29, 719–729. [Google Scholar] [CrossRef]
- Chen, C.H.; Ringelberg, C.S.; Gross, R.H.; Dunlap, J.C.; Loros, J.J. Genome-wide analysis of light-inducible responses reveals hierarchical light signalling in Neurospora. EMBO J. 2009, 28, 1029–1042. [Google Scholar] [CrossRef] [PubMed]
- Kritsky, M.S.; Russo, V.E.; Filippovich, S.Y.; Afanasieva, T.P.; Bachurina, G.P. The opposed effect of 5-azacytidine and light on the development of reproductive structures in Neurospora crassa. Photochem. Photobiol. 2002, 75, 79–83. [Google Scholar] [CrossRef]
- Li, X.; Wang, F.; Liu, M.; Dong, C. Hydrophobin CmHYD1 Is Involved in Conidiation, Infection and Primordium Formation, and Regulated by GATA Transcription Factor CmAreA in Edible Fungus, Cordyceps militaris. J. Fungi 2021, 7, 674. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Fungal Genus | Species | Number of GATA TFs |
|---|---|---|
| Aspergillus | Aspergillus nidulans | 6 |
| Aspergillus oryzae | 7 | |
| Aspergillus flavus | 6 | |
| Aspergillus niger | 6 | |
| Aspergillus fumigatu | 6 | |
| Aspergillus terreus | 1 | |
| Aspergillus clavatus | 6 | |
| Aspergillus nomiae | 6 | |
| Penicillium | Penicillium chrysogenum | 10 |
| Penicillium roqueforti | 5 | |
| Penicillium digitatum | 6 | |
| Fusarium | Fusarium tjaetaba | 4 |
| Fusarium oxysporum | 8 | |
| Fusarium fujikuroi | 3 | |
| Fusarium graminearum | 8 | |
| Neurospora | Neurospora crassa | 5 |
| Neurospora discreta | 6 | |
| Neurospora tetrasperma | 6 | |
| Mucor | Mucor mucedo | 2 |
| Mucor velutinosus | 3 | |
| Alternaria | Alternaria alternata | 6 |
| Alternaria burnsii | 2 | |
| Yarrowia | Yarrowia lipolytica | 10 |
| Sclerotinia | Sclerotinia sclerotiorum | 7 |
| Botrytis | Botrytis cinerea | 6 |
| Blastomyces | Blastomyces dermatitidis | 6 |
| Fungi Species | GATA TFs | GATA Zinc Finger | Zinc Finger Type | Number of Zinc Fingers (PF00320) | Reference |
|---|---|---|---|---|---|
| Aspergillus nidulans | AreA | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | [28,32,33,35,37] |
| AreB | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | ||
| SreA | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 2 | ||
| NsdD | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| LreA | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| LreB | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| Aspergillus oryzae | AoAreA | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | [29,38] |
| AoAreB | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | ||
| AoSreA | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 2 | ||
| AoNsdD | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| AoLreA | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| AoLreB | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| AoSnf5 | Cys-X2-Cys-X20-Cys-X2-Cys | - | 1 | ||
| Aspergillus flavus | AreA | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | [22,45] |
| AreB | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | ||
| SreA | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 2 | ||
| NsdD | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| LreA | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| LreB | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| Aspergillus niger | AreA | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | [6,43] |
| AreB | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | ||
| SreA | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 2 | ||
| NsdD | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| LreA | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| LreB | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| Aspergillus fumigatus | AreA | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | [18,19,39] |
| AreB | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | ||
| SreA | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 2 | ||
| NsdD | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| LreA | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| LreB | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| Aspergillus terreus | SreA | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 2 | - |
| Aspergillus clavatus | AreA | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | - |
| AreB | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | ||
| SreA | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 2 | ||
| NsdD | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| LreA | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| LreB | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| Aspergillus nomiae | AreA | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | - |
| AreB | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | ||
| SreA | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 2 | ||
| NsdD | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| LreA | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| LreB | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| Penicillium chrysogenum | AreA | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | [47,48] |
| NreB | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | ||
| SreP | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 2 | ||
| NsdD | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| WC1 | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| WC2 | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| Penicillium roqueforti | AreA | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | - |
| WC2 | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| Penicillium digitatum | AreA | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | - |
| SreP | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 2 | ||
| WC1 | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| WC2 | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| Ams2 | Cys-X2-Cys-X11-Cys-X2-Cys | - | 1 | ||
| Fusarium tjaetaba | ASD4 | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | - |
| Fusarium oxysporum | AreA | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | [50] |
| SreA | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 2 | ||
| NsdD | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| Fusarium fujikuroi | AreA | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | [30,31,55] |
| AreB | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | ||
| Csm1 | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| Neurospora crassa | NIT2 | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | [51,52,54] |
| WC-1 | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| WC-2 | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| ASD4 | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | ||
| SRE | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 2 | ||
| Alternaria alternata | AaAreA | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | [30] |
| AaAreB | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | ||
| AaSreA | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 2 | ||
| AaNsdD | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| AaLreA | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| AaLreB | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| Alternaria burnsii | SreP | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 2 | - |
| Botrytis cinerea | AreA | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | [56,57] |
| AreB | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | ||
| BcLTF1 | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| BcWCL2 | Cys-X2-Cys-X18-Cys-X2-Cys | IVb | 1 | ||
| Snf5 | Cys-X2-Cys-X8-Cys-X2-Cys | - | 1 | ||
| Blastomyces dermatitidis | AreA | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 1 | [58] |
| SreB | Cys-X2-Cys-X17-Cys-X2-Cys | IVa | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, D.; Zhao, R.; Lin, Y.; Jiang, C. Evolution and Functional Diversity of GATA Transcription Factors in Filamentous Fungi: Structural Characteristics, Metabolic Regulation and Environmental Response. Microbiol. Res. 2025, 16, 120. https://doi.org/10.3390/microbiolres16060120
Hu D, Zhao R, Lin Y, Jiang C. Evolution and Functional Diversity of GATA Transcription Factors in Filamentous Fungi: Structural Characteristics, Metabolic Regulation and Environmental Response. Microbiology Research. 2025; 16(6):120. https://doi.org/10.3390/microbiolres16060120
Chicago/Turabian StyleHu, Danni, Ruoyu Zhao, Yingxu Lin, and Chunmiao Jiang. 2025. "Evolution and Functional Diversity of GATA Transcription Factors in Filamentous Fungi: Structural Characteristics, Metabolic Regulation and Environmental Response" Microbiology Research 16, no. 6: 120. https://doi.org/10.3390/microbiolres16060120
APA StyleHu, D., Zhao, R., Lin, Y., & Jiang, C. (2025). Evolution and Functional Diversity of GATA Transcription Factors in Filamentous Fungi: Structural Characteristics, Metabolic Regulation and Environmental Response. Microbiology Research, 16(6), 120. https://doi.org/10.3390/microbiolres16060120