1. Introduction
Helichrysum arenarium (L.) Moench is a perennial herb that belongs to the genus Helichrysum within the Asteraceae family. This species is characterized by its woody and robust rhizome. Its medicinal significance is notable, attributed to the presence of tannins and volatile oils throughout all parts of the plant, which are known to stimulate bile secretion as well as exhibit anti-inflammatory and antioxidant properties. This herb thrives at altitudes ranging from 900 m to 2400 m on earthy hillocks, semi-sandy dunes, slopes of wet saline soils, gravelly terrains, sand dunes, and grasslands, and beneath pine forests [
1,
2]. The distribution of this species extends across Russia, Europe, Mongolia, and the Xinjiang Province in China. Within Xinjiang Province specifically, it can primarily be found in Altay City, Habahe County, Fuyun County, and Qinghe County. The Altay region is characterized by a continental north-temperate cold climate zone featuring dry and hot summers alongside cold winters. It experiences low precipitation levels coupled with high evaporation rates; there are significant temperature fluctuations between day and night, along with abundant sunlight [
3]. The sampling site itself exhibits sandy soil conditions combined with a dry climate marked by considerable diurnal temperature variations and ample sunlight. The primary chemical constituents of
H. arenarium (L.) Moench are predominantly flavonoids and polyphenolic compounds, with tannins and volatile oils present throughout the entire herb. Furthermore, the flower sap demonstrates significant effects in promoting bile secretion [
4,
5]. This plant exhibits a wide array of medicinal properties, including choleretic, hepatoprotective, and antiviral activities. Additionally, its flowers reveal lipid-lowering effects, with identified components such as lcariside F2, icari-side D1, phenethanol-β-vicianoside, benzyl alcohol-β-vicianoside, benzoic acid-β-D-gentiobioside, and phenethanol-β-D-gentiobioside [
6,
7,
8].
The rhizosphere is a multifunctional and dynamic zone in the soil, located approximately 5 mm from the root system, where essential nutrients for plant growth can be sourced from the nutrient sphere [
9]. This area serves as a crucial habitat for microorganisms, which play an integral role in the interactions within the plant rhizosphere. Root exudates are produced by various parts of the root system during plant growth. These exudates consist of a range of organic compounds, protons, and inorganic ions that are secreted or released into the surrounding environment [
10]. They collectively influence plant growth and development directly, as well as indirectly by affecting the surrounding ecosystem [
11]. Research indicates that higher local plant diversity correlates with greater variability in root exudates, which in turn enhances the diversity of soil microbial communities [
12]. Plant metabolites play a crucial role in shaping these microbial communities. Studies have demonstrated that root secretions can impact both the activity and species composition of rhizosphere microorganisms [
13]; they stimulate microbial growth, enhance their metabolic activity, accelerate nutrient cycling within the rhizosphere, and improve soil nutrient utilization. Through root secretions, plants regulate rhizosphere microflora to promote their own growth effectively [
14].
The root system of this plant enhances soil microorganisms through the secretion of various compounds. Rhizosphere microorganisms play a crucial role in regulating plant growth and development by engaging in metabolic activities. Bacteria and fungi are the primary types of rhizosphere microorganisms, with beneficial species promoting plant growth by facilitating soil nutrient cycling, secreting phytohormones, and inhibiting pathogenic bacteria. Conversely, harmful microorganisms can infect plants, leading to stunted growth or even death. Plants depend on the rhizosphere microbial community for their functions by acquiring nutrients from these organisms and under the influence of their activities and composition. The diversity of rhizosphere microbes is essential for maintaining stable soil ecosystems [
15,
16,
17]. Numerous studies have investigated the rhizosphere microbial communities associated with various plants worldwide, resulting in the isolation of functional strains that are widely utilized in biopromotion studies or antagonism research. However, no study has specifically addressed the rhizospheric microorganisms linked to
H. arenarium (L.) Moench. This unique plant, found in Xinjiang, possesses significant medicinal value. Moreover, demand for its output is increasing due to a growing market interest in Kozimuke granules—a traditional Kazakh medicine that utilizes it as a raw material. To establish a foundation for the large-scale cultivation of this plant, we conducted screenings for rhizosphere growth-promoting bacteria.
The objective of this study was to investigate the diversity of rhizosphere microorganisms associated with H. arenarium (L.) Moench through high-throughput sequencing techniques. This research lays a foundation for understanding the microbial species present in the rhizosphere, with a particular focus on rhizosphere probiotics and their role in regulating soil fertility and enhancing plant health.
2. Materials and Methods
2.1. Sample Collection
Soil samples were collected from Haba River County, located in the Altay Region of the Xinjiang Uygur Autonomous Region, at an elevation of 737.4 m in late July 2023 (48°18′ N, 86°78′ E) (
Figure 1). The samples were obtained utilizing a five-point sampling method. A sample square measuring 20 cm × 20 cm was collected using the five-point sampling method. From the five samples, three exhibiting optimal growth were randomly selected, and soil was collected from the root system at a depth of 0 to 5 mm. A total of 15 samples were collected.
Sample Pre-Treatment
The collected soil samples were meticulously homogenized, passed through a 2 mm sieve, divided into three aliquots, appropriately labeled, and stored at −80 °C and 4 °C in a refrigerator. For the soil collected by the 5-point sampling method, the soil of each point was fully mixed and 3 parallel samples were selected for high-throughput sequencing.
2.2. Cuiture Medium
NA medium: peptone, beef paste powder, NaCl, and Agarose. Inorganic phosphorus culture medium: glucose, (NH4)2SO4, yeast extract powder, NaCl, KCl, MgSO4, FeSO4, MnSO4, GaPO4, and agarose. Mongina medium: glucose, (NH4)2SO4, yeast extract powder, NaCl, KCl, MgSO4, FeSO4, MnSO4, GaCO4, and agarose. Ashby’s Medium: mannitol, CaCl2·2H2O, K2HPO4, MgSO4·7H2O, MoO3, FeCl3, and agar. CAS detection medium: piperazine diethanolsulfonic acid, agarose, peptone, MgSO4·7H2O, CaCl2, and agar. Silicate bacteria medium: agarose, (NH4)2SO4, yeast powder, MgSO4, Na2HPO4, FeSO4, MnSO4, potassium feldspar, and agar.
2.3. Quantification of Soil Nutrient Indicators
The organic matter content was assessed through potassium dichromate oxidation in conjunction with thermal oxidation. The total nitrogen content was determined by utilizing the Kjeldahl method, while hydrolyzable nitrogen content was evaluated using the alkaline diffusion technique. Effective phosphorus content was quantified via colorimetric analysis employing an ultraviolet spectrophotometer, and readily available potassium content was measured using flame photometry [
18].
2.4. Analysis of Microbial Amplicons in the Rhizosphere Using High-Throughput Sequencing
The extraction of genomic DNA from soil samples was performed by utilizing the TianGen magnetic bead-based soil genomic DNA extraction kit. This process was followed by an assessment of purity and concentration through 1% agarose gel electrophoresis. Subsequently, PCR amplification was conducted to obtain bacterial 16S rRNA sequences as well as fungal ITS sequences. Sequencing libraries were prepared using the NEB Next® Ultra™ II FS DNA PCR-free Library Prep Kit in accordance with the manufacturer’s guidelines, which included the addition of indexes. Following this, library quantification was carried out using Qubit and real-time PCR methods, while size distribution analysis was performed with a bioanalyzer. After quantification, the libraries were pooled together and subjected to sequencing on the Illumina platform, taking into account both effective library concentration and data requirements.
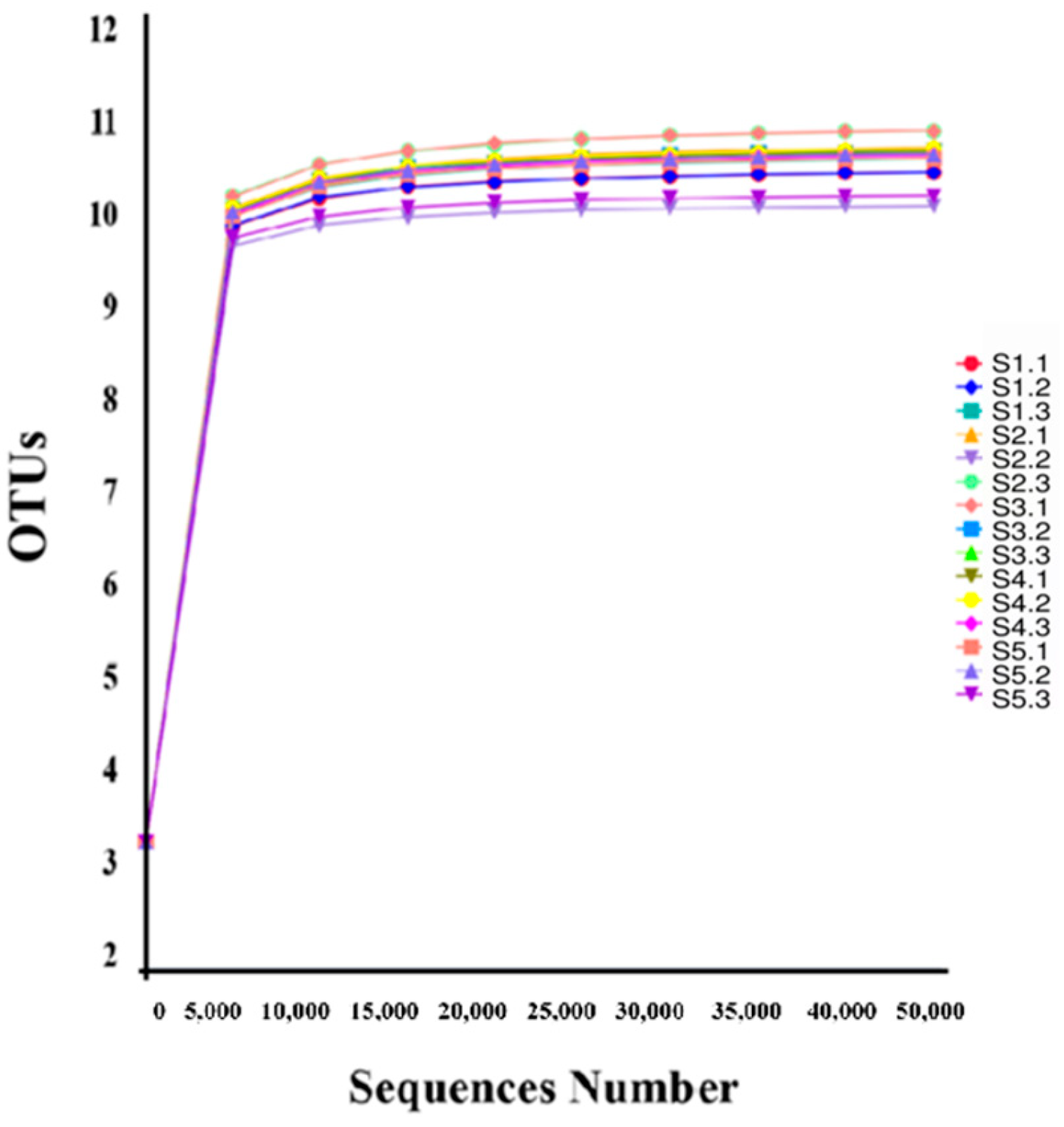
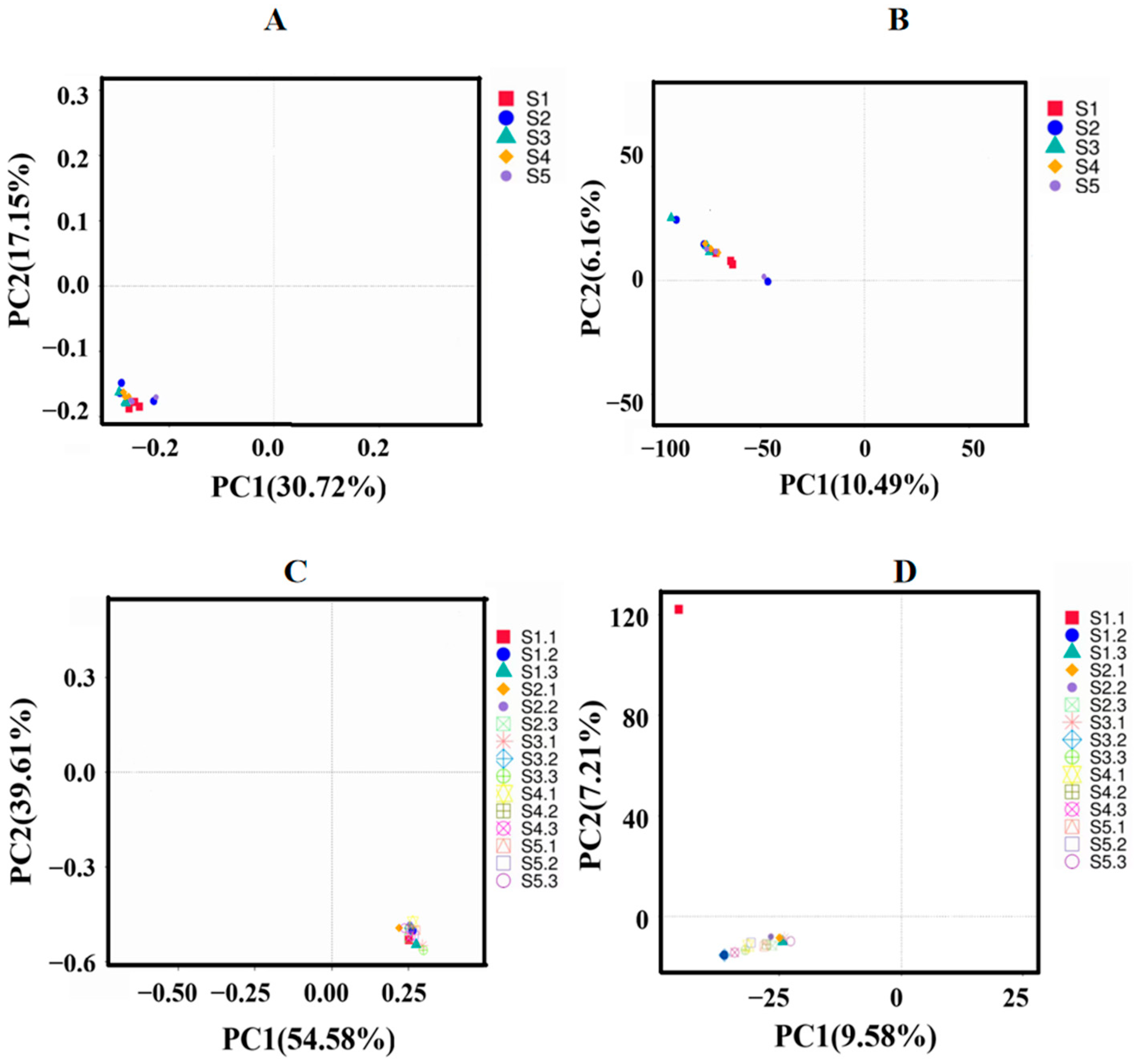
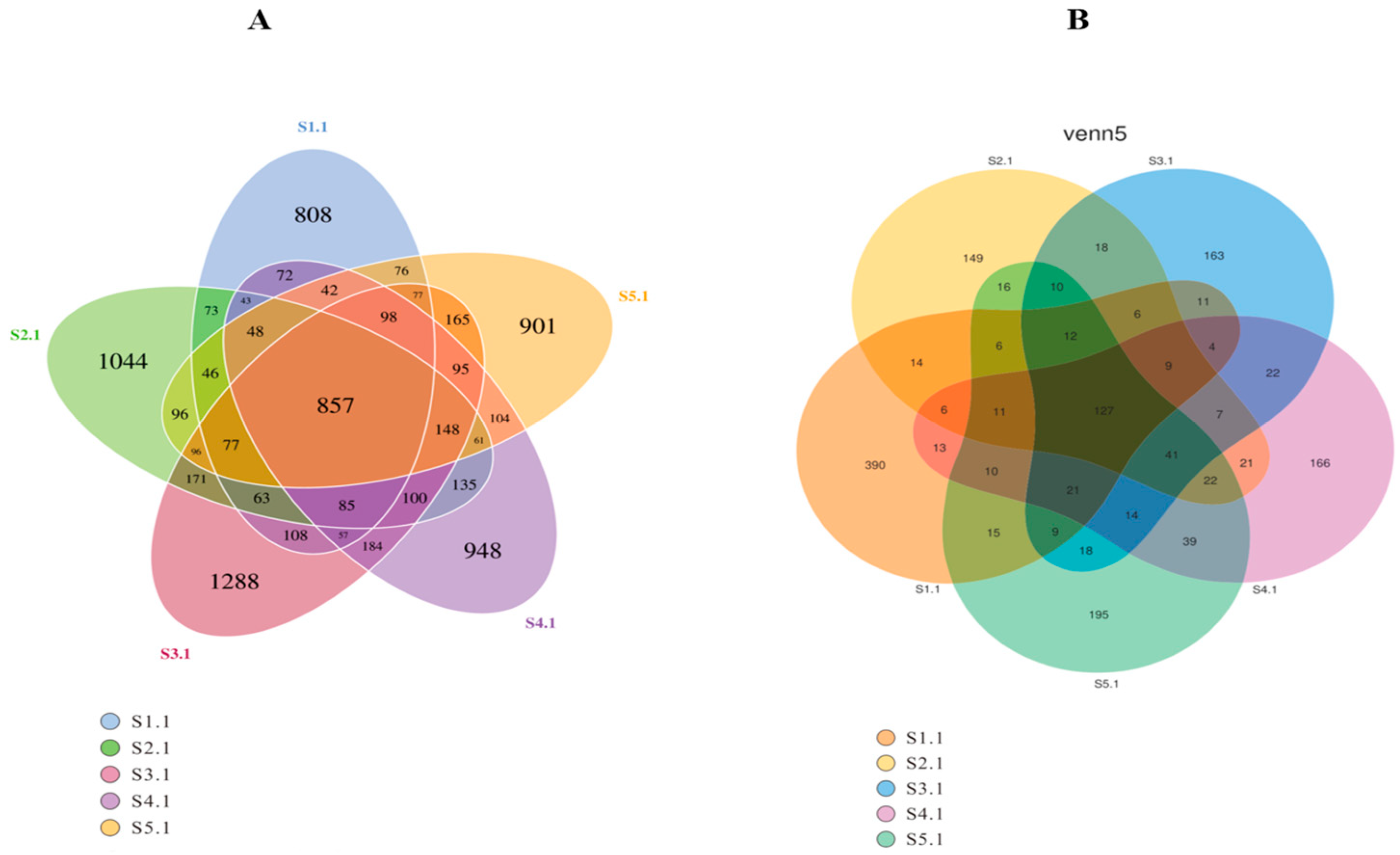
Mark the species annotation. A distribution histogram of relative abundance was generated in Perl using the SVG function. To visually represent variations in abundance and taxonomic clustering, a heatmap was created in R utilizing the pheatmap function, which incorporated abundance data for the top 35 taxa at each taxonomic rank from every sample. Commonalities and unique characteristics among different samples or groups were illustrated through Venn and flower diagrams, produced in R with the VennDiagram function and in Perl with the SVG function, respectively [
19].
2.5. Screening of Rhizosphere Growth-Promoting Bacteria in H. arenarium (L.) Moench
2.5.1. Isolation and Purification of Culturable Bacteria from the Rhizosphere of H. arenarium (L.) Moench
Each rhizosphere soil sample weighed 1 g and was mixed with 9 ml of sterile water using a vortex mixer for a duration of 30 min. The resulting mixture underwent gradient dilutions (10
−2, 10
−3, 10
−4, 10
−5, and 10
−6) employing the dilution coating method. A suspension of 100 μL was plated onto Nutrient Agar (NA) and a Gao I medium [
20], followed by incubation at 28 °C for seven days. Single colonies were subsequently selected from these cultures for purification and enumeration before being stored in a refrigerator maintained at an temperature of four degrees Celsius.
2.5.2. Analysis of Strains’ Phosphorus-Solubility Capacity
The strains obtained were inoculated into both an inorganic phosphorus medium and an organophosphorus medium. Each pure culture strain was introduced into three different media to observe the formation of phosphorus solubilization zones at a temperature of 28 °C over a period of seven days. Strains that exhibited significant phosphate-solubilizing zones were selected based on the measurements taken for both the diameter of the phosphorus solubilization zone (D) and the colony diameter (d). The ratio between these two measurements determined their effectiveness in terms of phosphorus solubilization capacity [
21].
2.5.3. Screening of Nitrogen-Fixing Bacteria
The isolated strains were inoculated onto Ashby medium using sterile toothpicks and subsequently incubated at a temperature of 28 °C for seven days. Bacterial colonies that exhibited viscous translucent growth with white, brown, or dark brown pigmentation indicated their nitrogen-fixing capability, which was assessed based on the measurement of their growth diameters [
22].
2.5.4. Screening of Strains with Iron Chelating Ability
The isolated strains were streaked onto a CAS detection medium, and each pure culture strain was incubated at 28 °C for 7 days with three replicates. Following this incubation period, the nitrogen fixation zone was examined, and strains that exhibited a pronounced orange chelating ring were selected. The iron-chelating capacity of the strains was assessed based on the diameter of the orange chelate ring [
23].
2.5.5. Revealing the Potential of Potassium Solubilizing Bacteria Through Screening
The strains were subsequently inoculated onto a silicate bacterial medium using the dot method and incubated at 28 °C for a duration of 7 days. The yellow halo surrounding each colony was monitored at regular intervals of 24 h. Upon completion of the 7-day incubation period, the potassium solubility of the strain was preliminarily assessed based on the size of the halo [
24].
2.6. Identification of Growth-Promoting Bacteria Using Molecular Biology Techniques
The selected growth-promoting bacteria underwent DNA extraction and amplification of their 16S rRNA region, the bacterial universal primers was as shown in
Table 1, the PCR system was as shown in
Table 2, and with PCR conditions (
Table 3). Strain identification and related primer sequences. The resulting amplification products were analyzed through electrophoresis and subsequently sent for sequencing to the Shanghai Shenggong Sequencing Company. Upon the completion of the sequencing process, alignment analysis was conducted on the NCBI website (
https://www.ncbi.nlm.nih.gov/, accessed on 13 August 2024). The NCBI website (
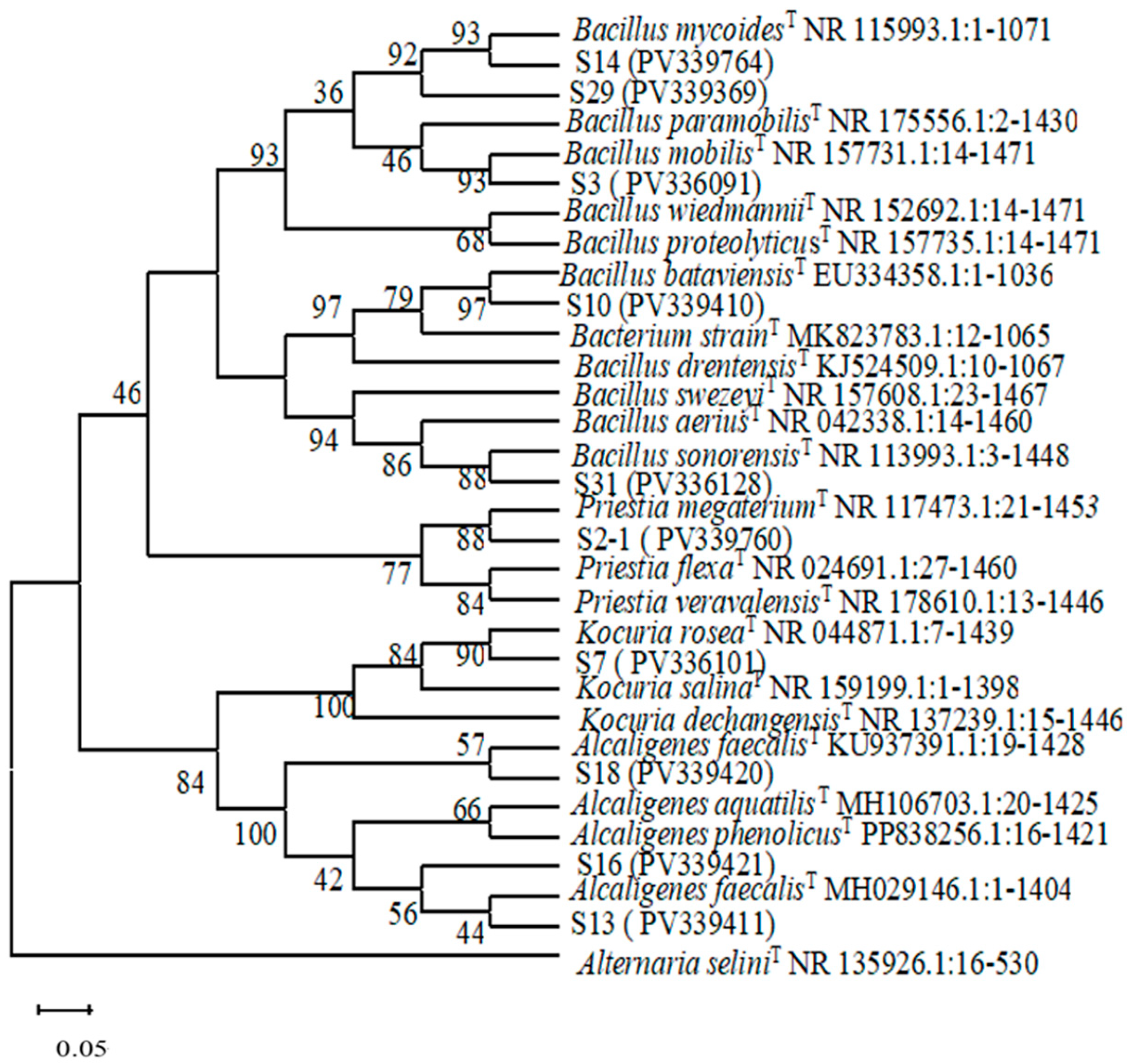
https://www.ncbi.nlm.nih.gov/, accessed on 22 March 2025) was utilized to submit the obtained DNA sequences to the GenBank database, where they were compared with known sequences. Sequences exhibiting high homology were selected for further analysis. A phylogenetic tree was constructed using MEGA 4.1 software, and we employed the Complete Deletion mode of the neighbor-joining method to build the tree.
4. Discussion
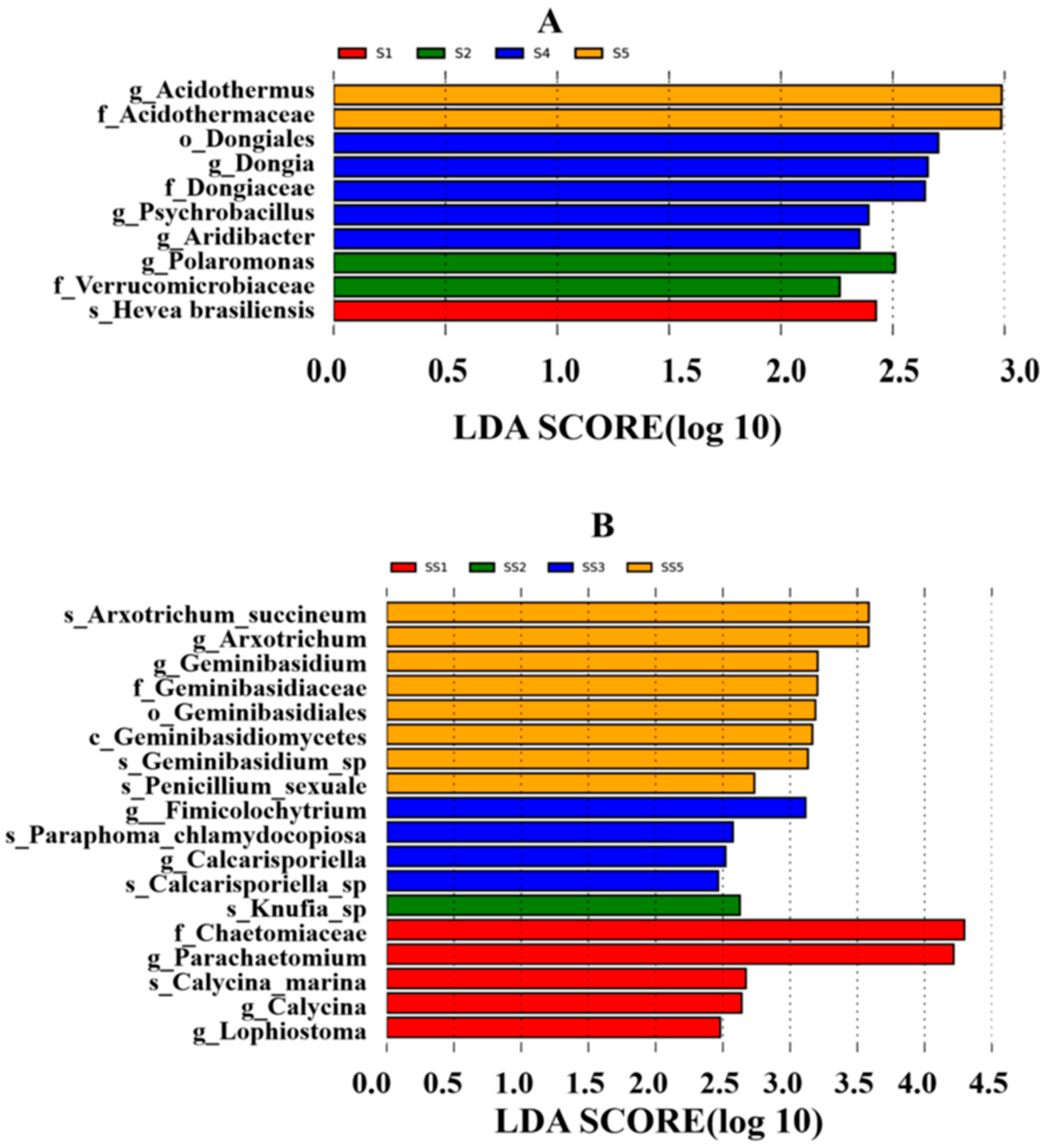
The microbial community structure in the rhizosphere soil plays a pivotal role in nutrient acquisition by
H. arenarium (L.) Moench. Consequently, we undertook a comprehensive study and analysis of the microbial community structure within the rhizosphere soil, alongside screening for growth-promoting and antagonistic bacteria. Our findings indicated that bacteria exhibited greater species richness, community diversity, and abundance compared to fungi in the rhizosphere soil environment. This observation underscores a predominant bacterial influence on root ecosystems, corroborated by previous studies highlighting bacterial dominance among soil microorganisms [
26,
27]. Furthermore, increased bacterial diversity is indicative of enhanced soil resilience [
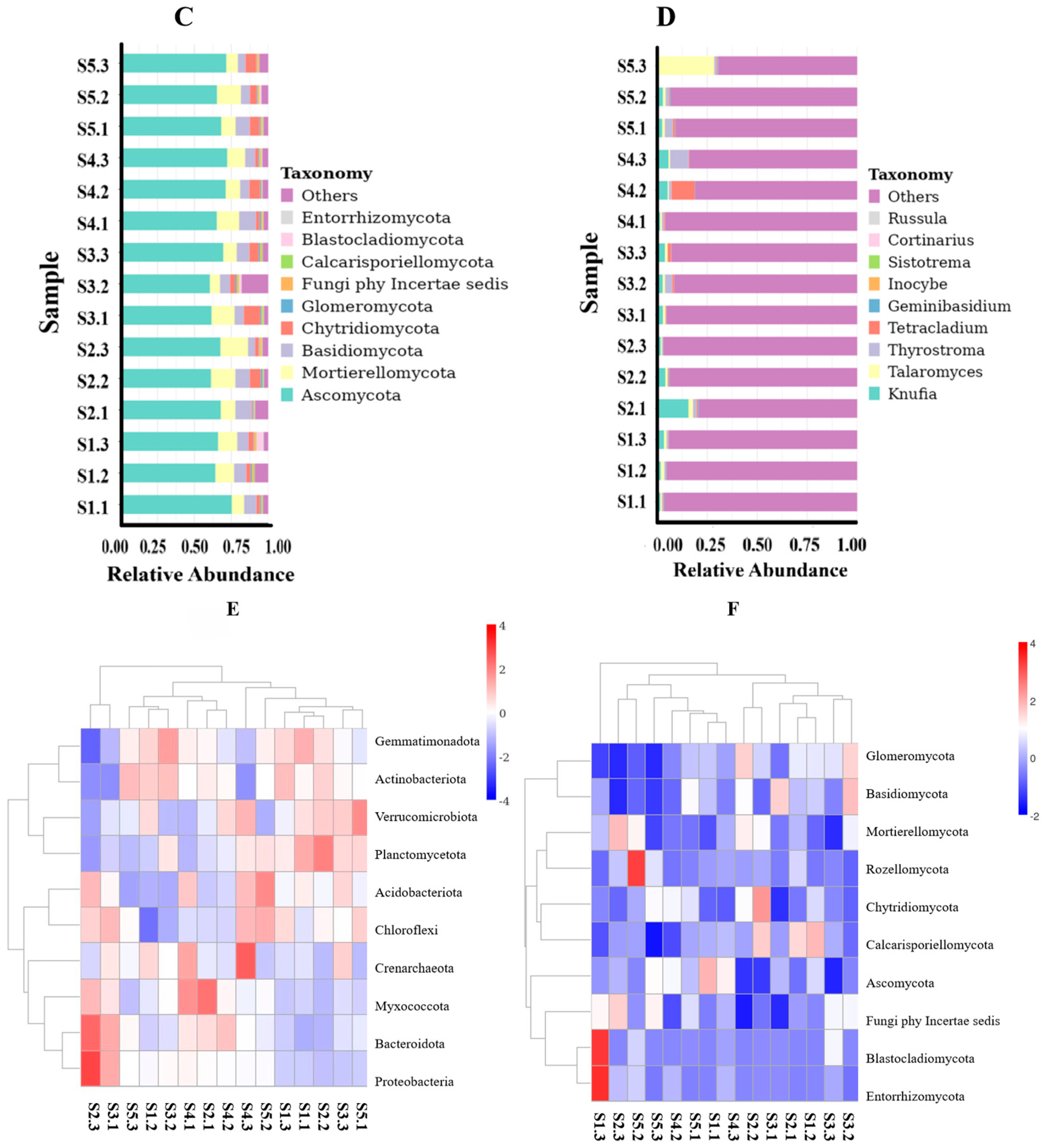
28]. Based on OTU analysis results, eight major bacterial phyla were identified within the rhizosphere microbial community (with relative abundances exceeding 1%), among which Actinobacteriota and Proteobacteria emerged as the dominant phyla. Notably, Proteobacteria accounted for over 40% of all identified phyla and were consistently enriched in the rhizosphere soils of various plants such as alfalfa [
29], strawberry [
30], potato [
31], and asparagus [
32]. This suggests that Proteobacteria possess adaptability to diverse plant-specific rhizospheric environments while serving as key constituents of these communities. However, it is important to note that different plant species exhibit distinct characteristics regarding microbial diversity within their respective rhizosphere soils. In our study area—which encompasses mounds, semi-dunes, and slopes with wet saline or gravelly soils at altitudes ranging from 900 m to 2400 m in pine forests or grasslands—pH levels ranged between 6.68 and 7.06, predominantly representing neutral to slightly acidic conditions.
Based on the unique growth environment, Actinobacteriota, Proteobacteria, Acidobacteriota, Verrucomicrobiota, and Gemmatimonadota are the predominant phyla in
H. arenarium (L.) Moench. However, while soil fungi are less abundant than bacteria, they play a critical role in maintaining ecosystems [
33]. In the rhizosphere microbial community of
H. arenarium (L.) Moench, four major fungal phyla exhibit relative abundances greater than 1%, with Ascomycota and Mortierellomycota being the dominant groups alongside Basidiomycota.
At the genus level, the relative abundance of
Candidatus Udaeobacter, 67-14, RB41,
Gemmatimonas, and MB-A2-108 exceeds 1%. The
Gemmatimonas belong to
Gemmatimonadota; 67-14 and MB-A2-108 belong to
Actinobacteriota. The results are consistent with those of dominant bacteria.
Gemmatimonas is a beneficial bacterium found in the rhizosphere that colonizes a majority of plants within this environment. RB41 was found to be colonized in the rhizosphere of winter wheat [
34].
Candidatus Udaeobacter is a dominant genus of bacteria and a key core group in rhizosphere soil [
32]; 67-14 is a bacterium that predominates in the rhizosphere of olive trees [
35]. MB-A2-108 has established itself in the rhizosphere of wheat [
36] and is also a predominant bacterium within the rhizosphere of Chinese cabbage [
37].
Notably, Ascomycota is the most prevalent phylum, accounting for over 70% of all identified taxa; this finding aligns with previous studies conducted by Li Qingshan et al. [
38]. Ascomycota primarily decomposes recalcitrant organic matter such as plant litter and the lignin present in soil. Ren Neifan et al. isolated a strain of Penicillium cinerea BIBA-G563 from healthy
Ruthenica roots belonging to Ascomycota, demonstrating a control efficacy of 75.4% against root rot [
39]. Given that
H. arenarium (L.) Moench is a root-breeding plant, it provides numerous attachment points for rhizosphere microorganisms, thereby enhancing their abundance and diversity while facilitating the enrichment of dominant flora.
At the genus level, the relative abundance of
Talaromyces,
Knufia,
Tetracladium, and
Thyrostroma is greater than 1%.
Talaromyces,
Knufia, and
Tetracladium are classified within the Ascomycota phylum, while
Thyrostroma is categorized under Basidiomycota. These classifications align with the findings regarding the dominant flora at the phylum level.
Talaromyces was identified in the blood-red mushroom pond [
40]. Xu Yunlong successfully isolated a strain of
Talaromyces purpureogenus, designated XZY3PSF, from corn rhizosphere soil. This strain has demonstrated a significant ability to enhance the primary nutritional indices of crops [
41].
Knufia was the predominant genus of fungi found in both herbaceous litter soil [
42] and Pinus
tabulaeformis sand-fixing forests [
43] within desert regions.
Tetracladium was predominantly found in the roots of P. muris [
44], while
Thyrostroma serves as the dominant fungal species within the stems of
P. pseudoephedra [
45].
Rhizosphere growth-promoting bacteria enhance plant nutrient absorption through various mechanisms, including biological nitrogen fixation, phosphorus solubilization, potassium solubilization, the secretion of plant hormones, and the production of iron carriers. In this study, we screened nine strains of growth-promoting bacteria: five strains belonging to the genus Bacillus within the phylum Firmicutes; three strains from the genus Alcaligenes under Proteobacteria; and one strain from the genus Kocuria in the Actinobacteria phylum. Firmicutes and Proteobacteria were identified as rhizosphere growth-promoting bacteria and dominant taxa in high-throughput sequencing analyses. Additionally, Bacillus emerged as a predominant growth-promoting bacterium within the rhizosphere, aligning with the findings from high-throughput sequencing results.
Previous studies have predominantly identified
Bacillus and
Pseudomonas as the primary growth-promoting bacteria. Among these, strain S16, which belongs to
Alcaligenes, exhibits the highest capacity for phosphorus detoxification and iron carrier production. In contrast, strain S10 from
Bacillus demonstrates superior nitrogen fixation abilities. Nitrogen-fixing bacteria convert atmospheric nitrogen into ammonia, thereby enhancing soil nitrogen content. Meanwhile, phosphorus-solubilizing bacteria transform insoluble phosphorus into forms that are accessible to plants, increasing effective phosphorus levels in the soil. Additionally, bacterial strains that produce iron carriers facilitate iron nutrition in plants through the synthesis and utilization of these carriers [
46,
47,
48]. In this study, the nitrogen-fixing medium was employed to assess the nitrogen-fixing capabilities of various strains. However, the nitrogen-fixing medium alone proved to be somewhat inadequate for a comprehensive evaluation of these abilities. Over 90% of rhizosphere microorganisms remain unculturable using traditional media, leading to the identification of only a limited number of culturable strains (such as
Bacillus and
Pseudomonas). This limitation results in the underestimation of the diversity present among rhizosphere nitrogen-fixing bacteria. Furthermore, certain azotobacter species within the rhizosphere require plant signaling molecules (such as nodulation factors) or collaboration with other microorganisms for their activation. When the strain is cultured in isolation, its nitrogen-fixing genes may become silenced, resulting in the inaccurate assessment of the strain’s functional capabilities. Furthermore, different strains may only exhibit nitrogen fixation abilities under specific plant host or soil conditions. The homogenization condition of the medium fails to adequately reflect niche differentiation, leading to the exclusion of certain functional strains. Furthermore, bacteria within the medium may experience survival stress (such as carbon limitation), which can inhibit nitrogenase expression. In contrast, carbon sources secreted by plant roots in natural environments (such as organic acids) have the potential to enhance nitrogen fixation activity. In the subsequent phase, quantifiable techniques such as the acetylene reduction assay (ARA) can be employed to assess the nitrogenase activity of various strains, thereby facilitating the identification of superior nitrogen-fixing bacteria. This study focused exclusively on isolating and identifying rhizosphere growth-promoting bacteria. It is important to note that the strains isolated through culturable methods only represent a fraction of the culturable component within the rhizosphere microbial community; consequently, there may be an underestimation of the diversity present among actual functional strains. In subsequent phases of this study, optimal growth-promoting bacteria will be selected based on strain combination experiments. This approach aims to establish an experimental foundation for developing and promoting microbial fertilizers specific to
H. arenarium (L.) Moench, while also laying a research groundwork for conserving wild resources of
H. arenarium (L.) Moench, exploring rhizosphere microorganism development and utilization, and providing robust support for future applications of
H. arenarium (L.) Moench in food and medicine.
5. Conclusions
The high-throughput sequencing results identified Actinomycetes, Proteobacteria, and Blastomonas as the predominant bacterial phyla in the rhizosphere soil of H. arenarium (L.) Moench, while Ascomycetes and Mortieromycetes were recognized as the primary fungal phyla. The assessment of efficiency promotion for 43 bacterial species isolated from the rhizosphere soil of H. arenarium (L.) Moench demonstrated that three strains—namely S31, S16, and S29—exhibited phosphorus solubilization capabilities. Notably, strain S16 exhibited the highest inorganic phosphorus solubility index at 1.23. Furthermore, strain S31 displayed organophosphorus solubilization abilities. Additionally, strains S7 and S10 possessed nitrogen fixation capacity; however, none of the isolated strains demonstrated proficiency in dissolving potassium compounds.
A total of ten strains with exceptional iron-bearing capacity were identified, among which strain S16 demonstrated both the highest D/d value and the most robust iron-bearing capability. These ten growth-promoting strains belong to the genera Bacillus, Alcaligenes, and Kocuria. In particular, this is Priestia megaterium for strain S2-1; B. mobilis for strain S3; Kocuria rosea for strain S7; B. bataviensis for strain S10; B. mycoides for strain S14; B. paramobilis for strain S29; B. sonorensis for strain S31; and A. faecalis for strains S18, S13, and S16.
In this study, we focused exclusively on analyzing the rhizosphere community structure of wild H. arenarium (L.) Moench and conducted an initial screening of growth-promoting bacteria. However, there remains significant potential for further research in this area. In subsequent phases, we will quantitatively assess the growth-promoting bacteria for their capabilities in nitrogen fixation, phosphorus solubilization, and indole-3-acetic acid (IAA) production. Additionally, we aim to identify optimal strains of growth-promoting bacteria through experiments designed to evaluate their efficacy in promoting the growth of H. arenarium (L.) Moench. Our study provides a foundational experimental basis for the development and dissemination of microbiological fertilizers derived from H. arenarium (L.) Moench.


 and
and 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}