
Biogas Dynamics and Microbial Composition Employing Different Inocula and Substrates in Continuous Stirred-Tank Reactors



Abstract
1. Introduction
2. Materials and Methods
Substrates Collection, Processing and Storage
3. Results and Discussion
3.1. Impact of Substrates Properties on Anaerobic Digestion
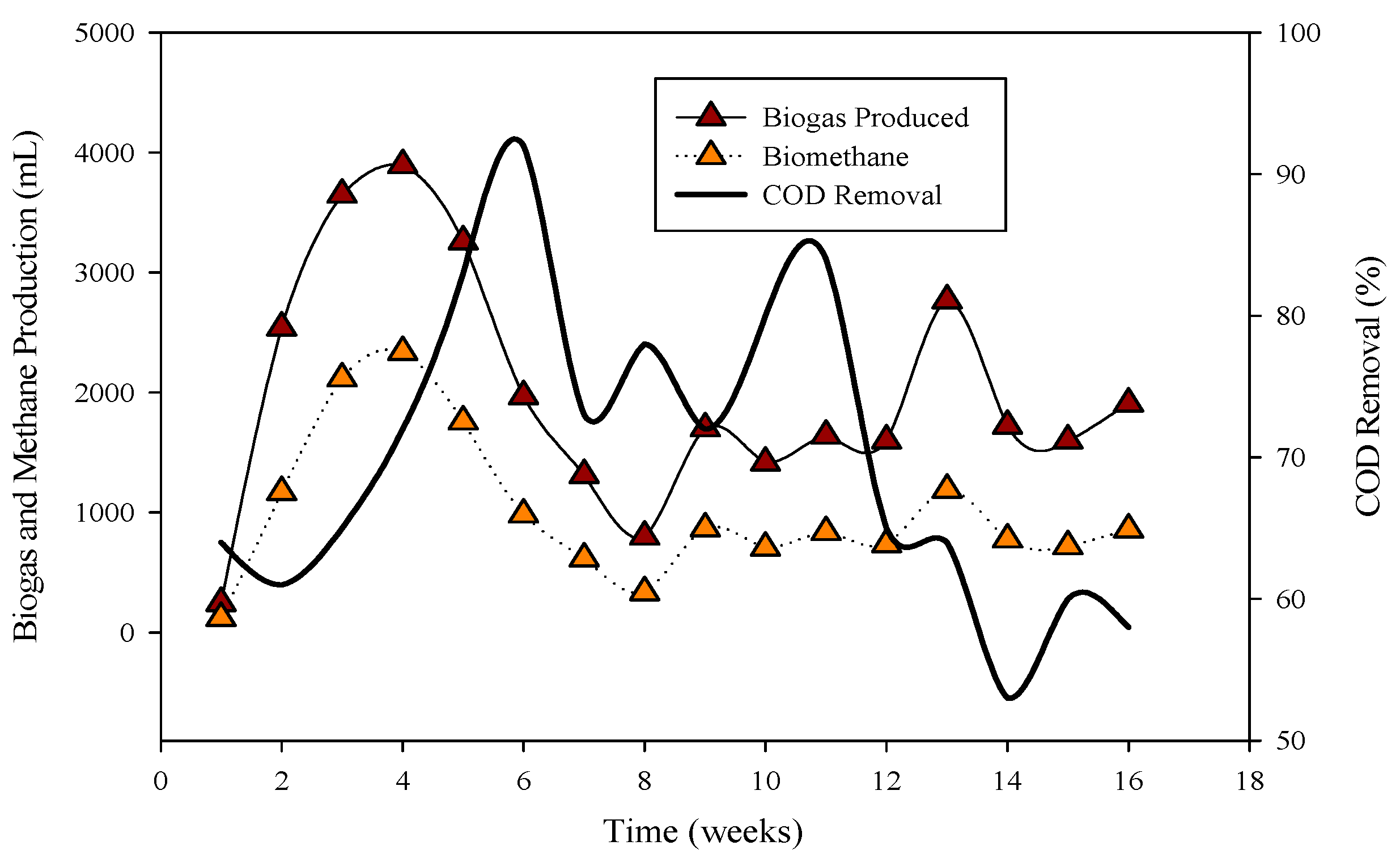
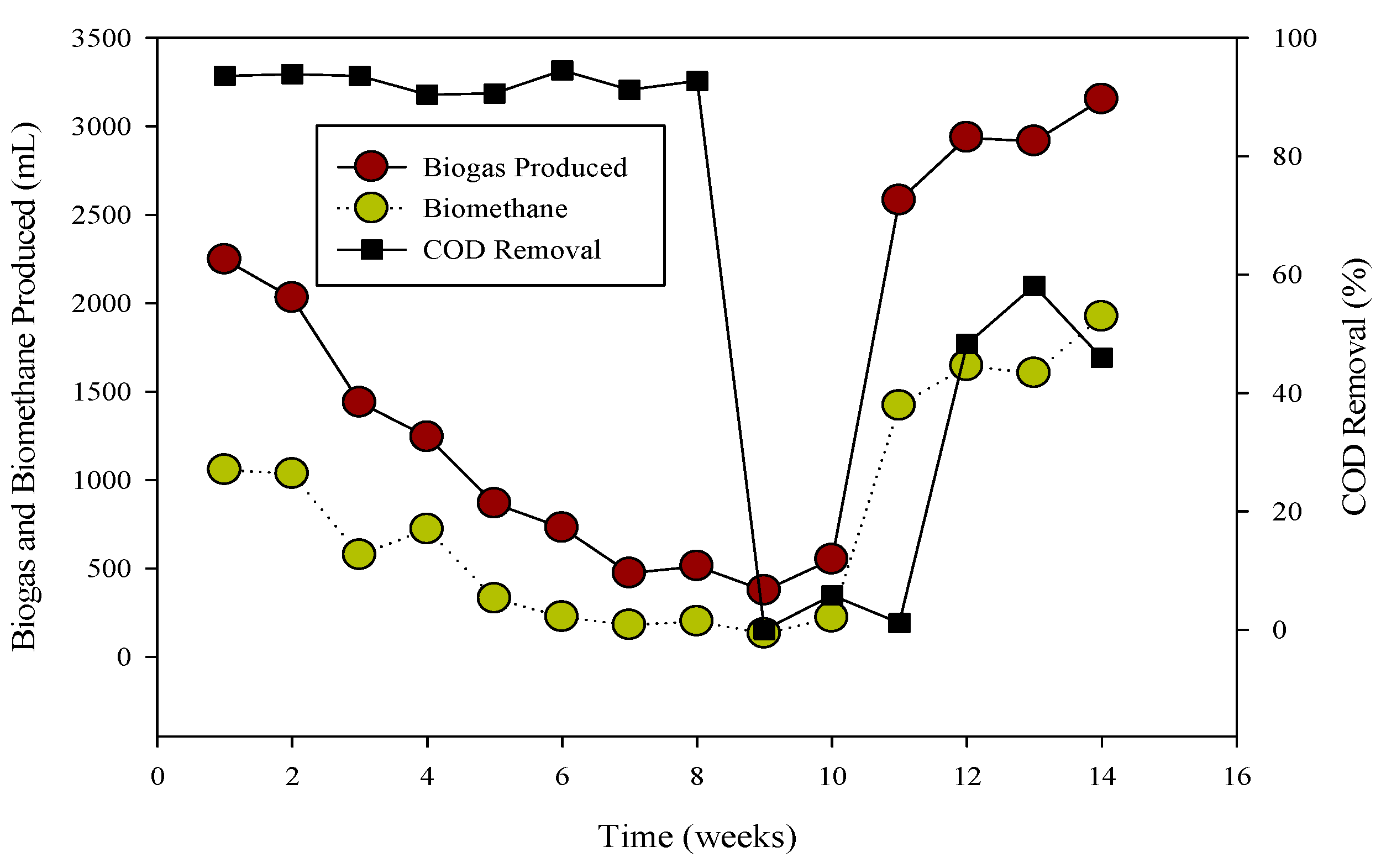
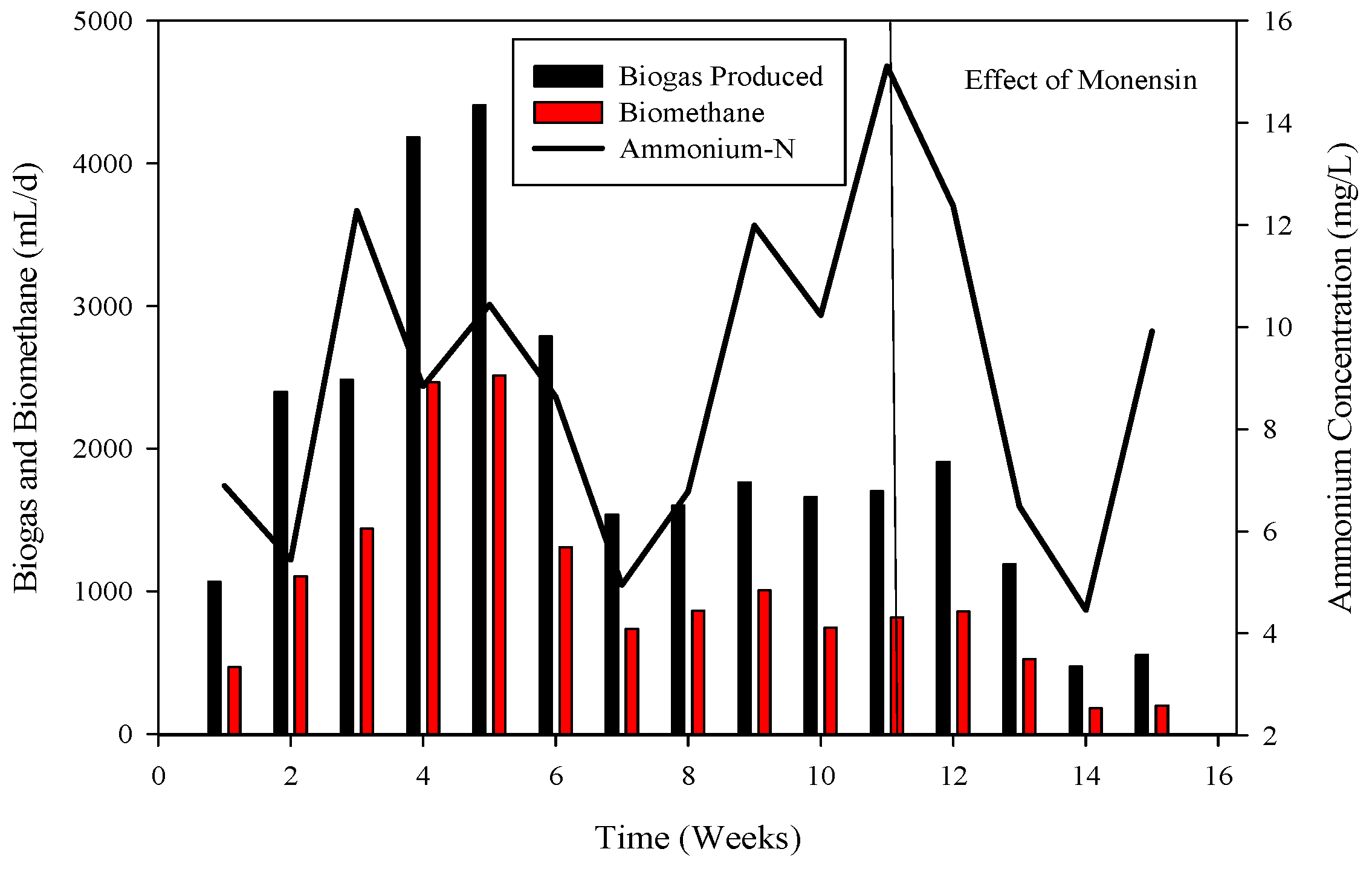
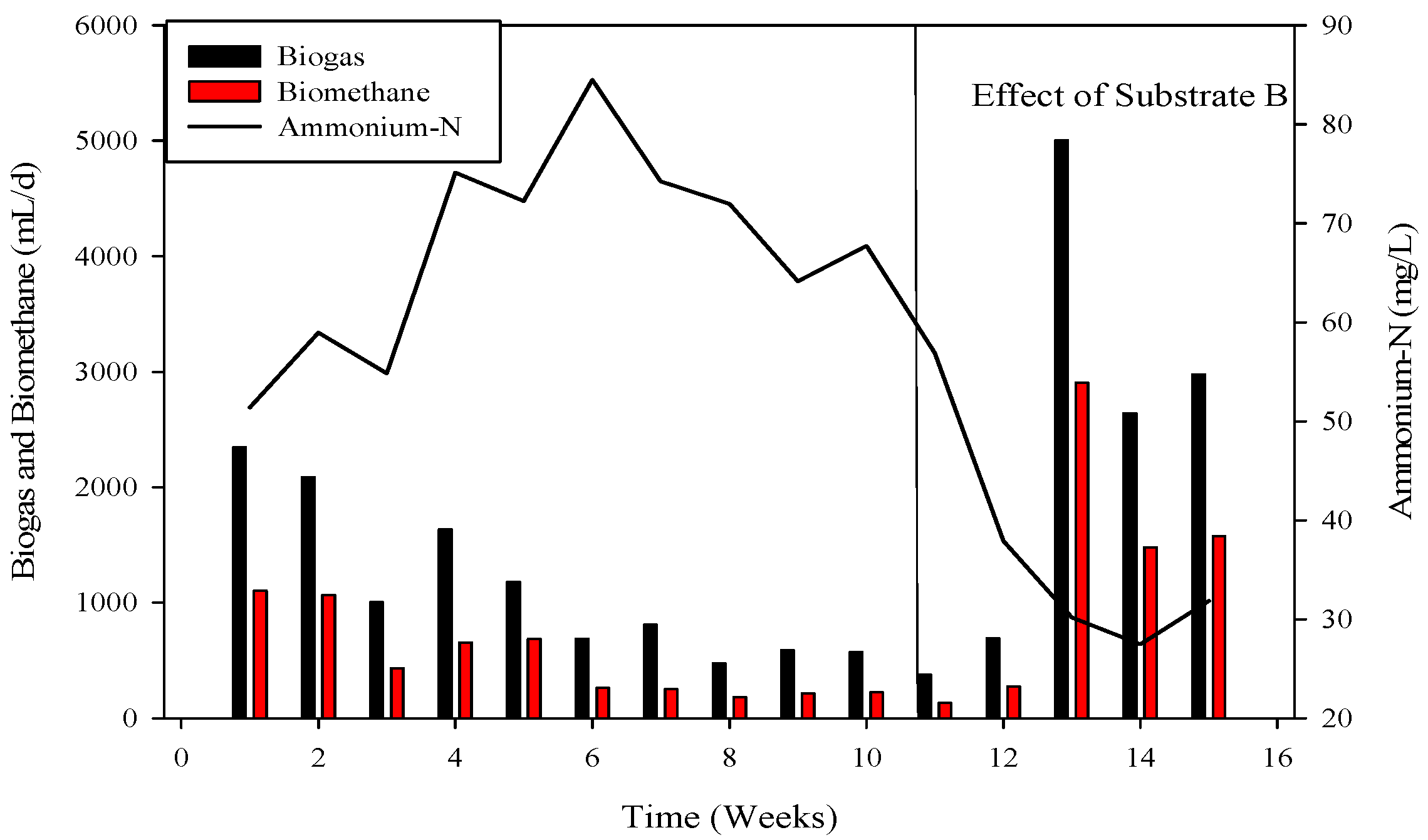
3.2. Process Stability and Biogas Generation
3.3. Perturbed System with Monensin
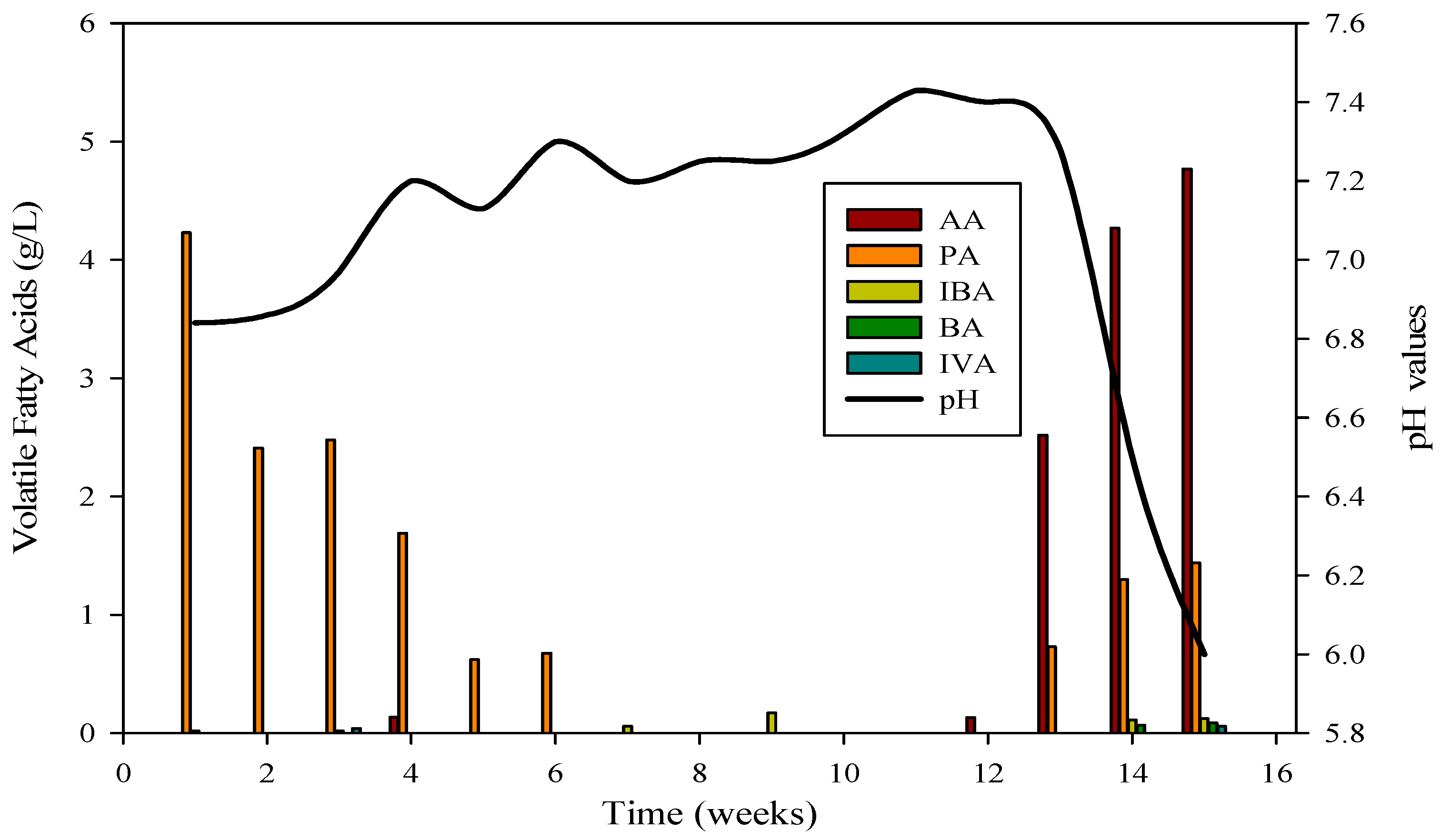
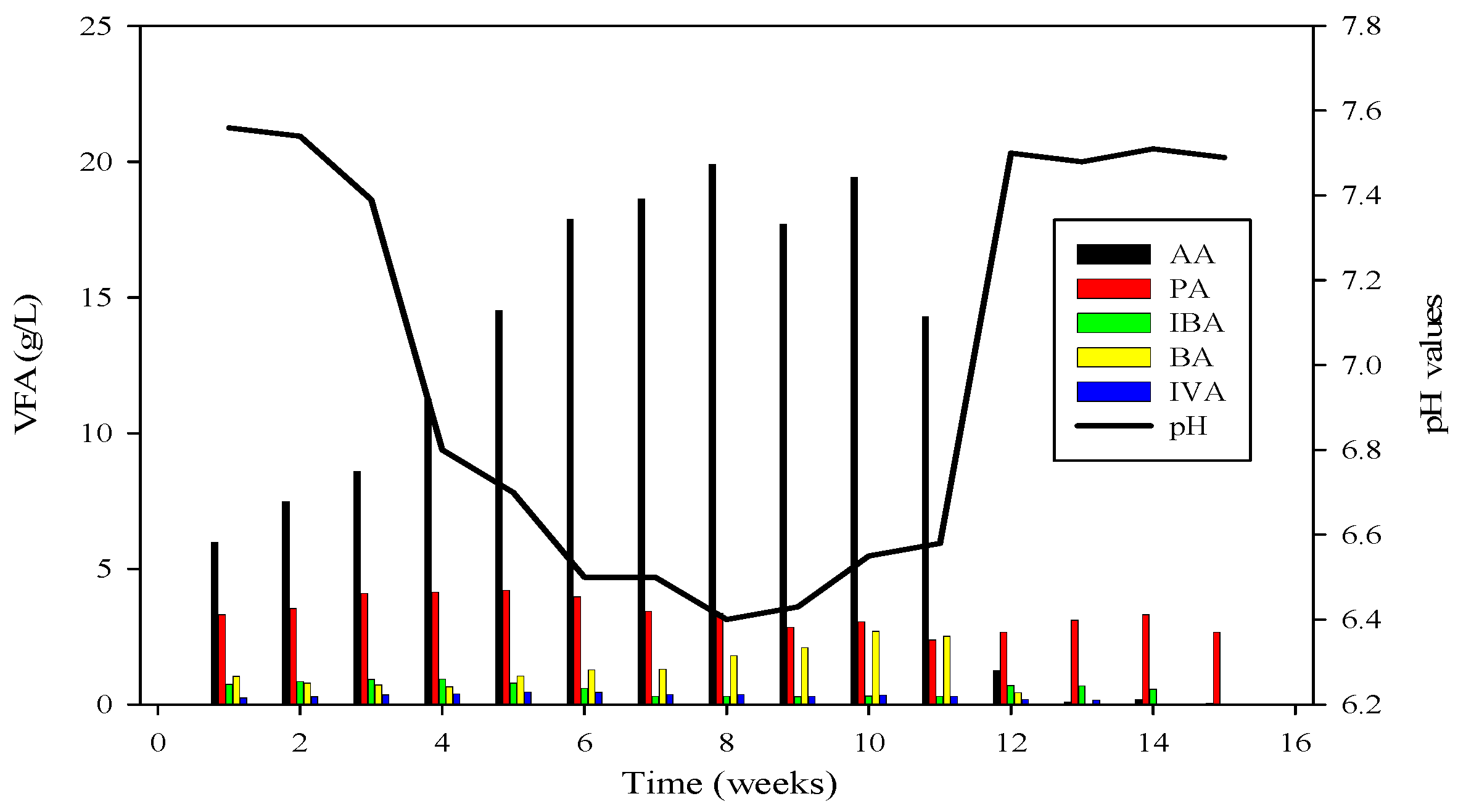
3.4. Volatile Fatty Acid (VFA) Profile
3.5. Effect of Ammonia
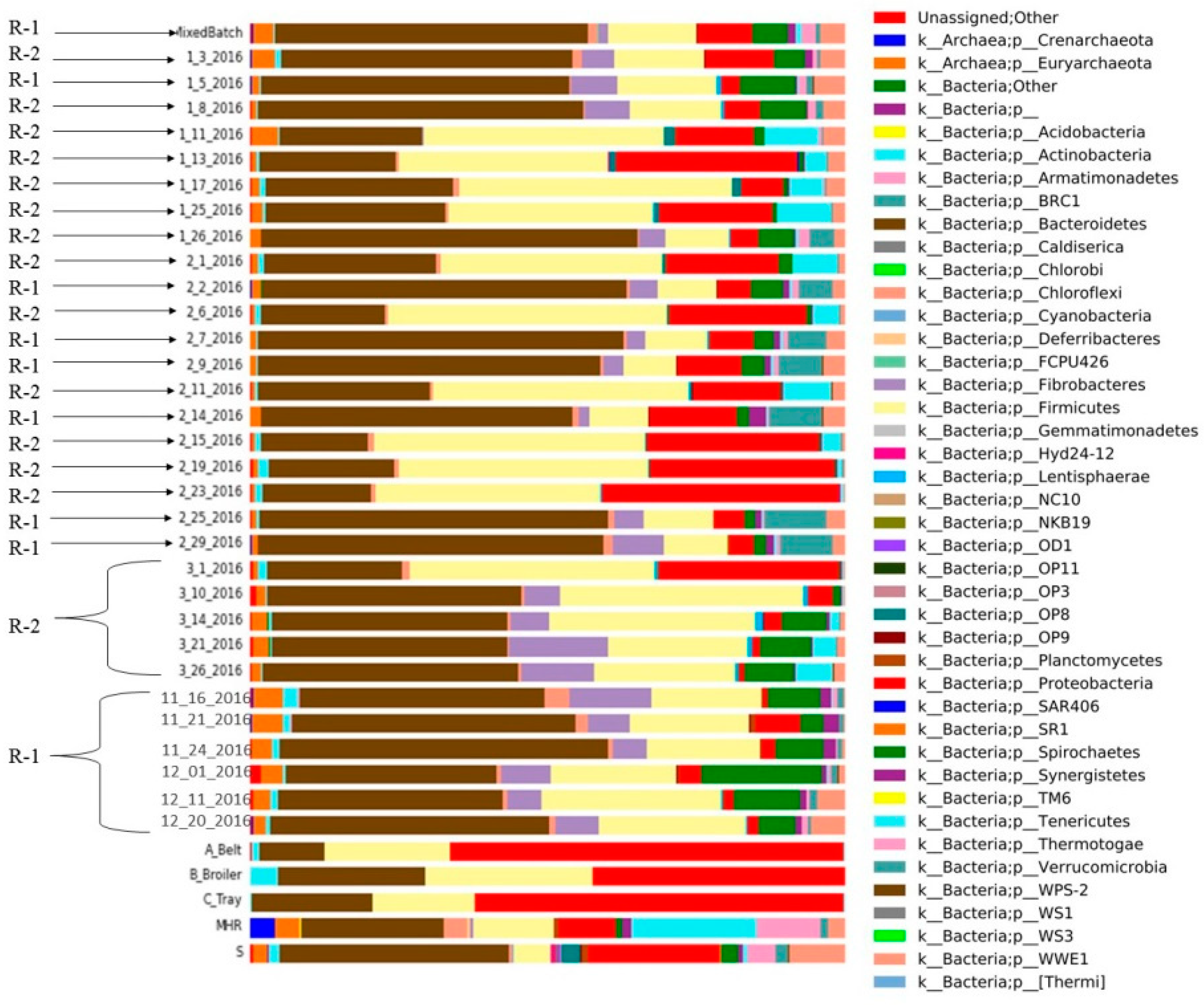
3.6. Microbial Composition
4. Discussion
5. Conclusions
- Poultry substrate B, processed in the R-1 CSTR with cow rumen inoculum, demonstrated superior and consistent biogas generation compared to cattle inoculum in R-1 and municipal sanitary inoculum in R-2. The study highlights the pivotal role of the optimal C: N ratio (17:1) in substrate B, contributing to successful AD.
- Co-digestion of belt waste (A) and tray hatchery resulted in substantial biogas and biomethane yield, particularly adaptable to cattle inoculum. Inhibitory effects observed in stillage, attributed to elevated sulfur content, led to a substrate switch to poultry B, ensuring cumulative biogas production and reactor stability on the 80th day.
- Bacteriodes predominated in R-1, while fermicutes exhibited lower abundance compared to R-2. Proteobacteria, initially less prevalent in R-1, showed consistent presence in the substrate and inoculum. Spirochetes were present in both reactors, with higher abundance in R-1 initially. Fibrobacteria showed high initial abundance but decreased by the experiment’s end. Chloroflexi bacteria were consistently present in both reactors, while Tenericutes were exclusive to R-2.
- The study proposes the use of natural accelerants like laterite and industrial accelerants such as fly ash, bag filter gas dust, and phosphorus dust from the fertilizer industry. These additions aim to enhance microbial syntrophy, providing essential trace elements to mitigate ammonia toxicity in the biogas digester, thereby improving overall processing within the circular economy.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Archana, K.; Visckram, A.S.; Senthil Kumar, P.; Manikandan, S.; Saravanan, A.; Natrayan, L. A review on recent technological breakthroughs in anaerobic digestion of organic biowaste for biogas generation: Challenges towards sustainable development goals. Fuel 2024, 358, 130298. [Google Scholar] [CrossRef]
- Beausang, C.; McDonnell, K.; Murphy, F. Anaerobic digestion of poultry litter—A consequential life cycle assessment. Sci. Total Environ. 2020, 735, 139494. [Google Scholar] [CrossRef] [PubMed]
- Bhatnagar, N.; Ryan, D.; Murphy, R.; Enright, A. A comprehensive review of green policy, anaerobic digestion of animal manure and chicken litter feedstock potential–Global and Irish perspective. Renew. Sustain. Energy Rev. 2022, 154, 111884. [Google Scholar] [CrossRef]
- Mansour, M.-N.; Lendormi, T.; Drévillon, L.; Naji, A.; Louka, N.; Maroun, R.G.; Hobaika, Z.; Lanoisellé, J.-L. Influence of substrate/inoculum ratio, inoculum source and ammonia inhibition on anaerobic digestion of poultry waste. Environ. Technol. 2022, 1–14. [Google Scholar] [CrossRef]
- De Vrieze, J.; Gildemyn, S.; Vilchez-Vargas, R.; Jáuregui, R.; Pieper, D.H.; Verstraete, W.; Boon, N. Inoculum selection is crucial to ensure operational stability in anaerobic digestion. Appl. Microbiol. Biotechnol. 2015, 99, 189–199. [Google Scholar] [CrossRef]
- Shah, F.A.; Mahmood, Q.; Iqbal, A.; Mackie, R.I. Inhibition of biomethane production under antibiotics in poultry litter. Arab. J. Geosci. 2020, 13, 1–10. [Google Scholar] [CrossRef]
- Fotidis, I.A.; Karakashev, D.; Kotsopoulos, T.A.; Martzopoulos, G.G.; Irini Angelidaki, I. Effect of ammonium and acetate on methanogenic pathway and methanogenic community composition. FEMS Microbiol. Ecol. 2013, 83, 38–48. [Google Scholar] [CrossRef]
- Rhee, C.; Park, S.-G.; Kim, D.W.; Yu, S.I.; Shin, J.; Hwang, S.; Shin, S.G. Tracking microbial community shifts during recovery process in overloaded anaerobic digesters under biological and non-biological supplementation strategies. Bioresour. Technol. 2021, 340, 125614. [Google Scholar] [CrossRef]
- Mahdy, A.; Bi, S.; Song, Y.; Qiao, W.; Dong, R. Overcome inhibition of anaerobic digestion of chicken manure under ammonia-stressed condition by lowering the organic loading rate. Bioresour. Technol. Rep. 2020, 9, 100359. [Google Scholar] [CrossRef]
- Christou, M.; Vasileiadis, S.; Kalamaras, S.; Karpouzas, D.; Angelidaki, I.; Kotsopoulos, T. Ammonia-induced inhibition of manure-based continuous biomethanation process under different organic loading rates and associated microbial community dynamics. Bioresour. Technol. 2021, 320, 124323. [Google Scholar] [CrossRef]
- Laloučková, K.; Skřivanová, E. Antibiotic resistance in livestock breeding: A review. Sci. Agric. Bohem. 2019, 50, 15–22. [Google Scholar] [CrossRef]
- Chen, Y.; Cheng, J.J.; Creamer, K.S. Inhibition of anaerobic digestion process: A review. Bioresour. Technol. 2008, 99, 4044–4064. [Google Scholar] [CrossRef]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 10 July 2023).
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Kunin, V.; Engelbrektson, A.; Ochman, H.; Hugenholtz, P. Wrinkles in the rare biosphere: Pyrosequencing errors can lead to artificial inflation of diversity estimates. Environ. Microbiol. 2010, 12, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.R.; Wang, Q.; Cardenas, E.; Fish, J.; Chai, B.; Farris, R.J.; Kulam-Syed-Mohideen, A.; McGarrell, D.M.; Marsh, T.; Garrity, G.M. The Ribosomal Database Project: Improved alignments and new tools for rRNA analysis. Nucleic Acids Res. 2009, 37 (Suppl. 1), D141–D145. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Zhang, M.; Fan, Z.; Hu, Z.; Luo, X. Enhanced anaerobic digestion with the addition of chelator-nickel complexes to improve nickel bioavailability. Sci. Total Environ. 2021, 759, 143458. [Google Scholar] [CrossRef]
- Li, X.; Wu, M.; Xue, Y. Nickel-loaded shrimp shell biochar enhances batch anaerobic digestion of food waste. Bioresour. Technol. 2022, 352, 127092. [Google Scholar] [CrossRef] [PubMed]
- Oosterkamp, M.J.; Méndez-García, C.; Kim, C.-H.; Bauer, S.; Ibáñez, A.B.; Zimmerman, S.; Hong, P.-Y.; Cann, I.K.; Mackie, R.I. Lignocellulose-derived thin stillage composition and efficient biological treatment with a high-rate hybrid anaerobic bioreactor system. Biotechnol. Biofuels 2016, 9, 120. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, W.; Wang, X.; Gabauer, W.; Ortner, M.; Li, Z. Tackling ammonia inhibition for efficient biogas production from chicken manure: Status and technical trends in Europe and China. Renew. Sustain. Energy Rev. 2018, 97, 186–199. [Google Scholar] [CrossRef]
- Hook, S.E.; Northwood, K.S.; Wright, A.D.; McBride, B.W. Long-term monensin supplementation does not significantly affect the quantity or diversity of methanogens in the rumen of the lactating dairy cow. Appllied Environ. Microbiol. 2009, 75, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Schnürer, A.; Dolfing, J.; Westerholm, M. Syntrophic entanglements for propionate and acetate oxidation under thermophilic and high-ammonia conditions. ISME J. 2023, 17, 1966–1978. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, C.; Yuan, Z.; Wang, R.; Angelidaki, I.; Zhu, G. Syntrophy mechanism, microbial population, and process optimization for volatile fatty acids metabolism in anaerobic digestion. Chem. Eng. J. 2023, 452, 139137. [Google Scholar] [CrossRef]
- Beneragama, N.; Moriya, Y.; Yamashiro, T.; Iwasaki, M.; Lateef, S.A.; Ying, C.; Umetsu, K. The survival of cefazolin-resistant bacteria in mesophilic co-digestion of dairy manure and waste milk. Waste Manag. Res. 2013, 31, 843–848. [Google Scholar] [CrossRef]
- Rajagopal, R.; Massé, D.I.; Singh, G. A critical review on inhibition of anaerobic digestion process by excess ammonia. Bioresour. Technol. 2013, 143, 632–641. [Google Scholar] [CrossRef]
- Liu, F.; Zhang, Y.; Zhang, Y.; Yang, J.; Shen, W.; Yang, S.; Liu, B.; Yuan, Z.; Zhang, Y. Thermodynamic restrictions determine ammonia tolerance of functional floras during anaerobic digestion. Bioresour. Technol. 2023, 391, 129919. [Google Scholar] [CrossRef]
- Song, M.; Shin, S.G.; Hwang, S. Methanogenic population dynamics assessed by real-time quantitative PCR in sludge granule in upflow anaerobic sludge blanket treating swine wastewater. Bioresour. Technol. 2010, 101, S23–S28. [Google Scholar] [CrossRef]
- Bassani, I.; Kougias, P.G.; Treu, L.; Angelidaki, I. Biogas upgrading via hydrogenotrophic methanogenesis in two-stage continuous stirred tank reactors at mesophilic and thermophilic conditions. Environ. Sci. Technol. 2015, 49, 12585–12593. [Google Scholar] [CrossRef] [PubMed]
- Belostotskiy, D.E.; Ziganshina, E.E.; Siniagina, M.; Boulygina, E.A.; Miluykov, V.A.; Ziganshin, A.M. Impact of the substrate loading regime and phosphoric acid supplementation on performance of biogas reactors and microbial community dynamics during anaerobic digestion of chicken wastes. Bioresour. Technol. 2015, 193, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Hinks, J.; Edwards, S.; Sallis, P.J.; Caldwell, G.S. The steady state anaerobic digestion of Laminaria hyperborea–Effect of hydraulic residence on biogas production and bacterial community composition. Bioresour. Technol. 2013, 143, 221–230. [Google Scholar] [CrossRef]
- De Francisci, D.; Kougias, P.G.; Treu, L.; Campanaro, S.; Angelidaki, I. Microbial diversity and dynamicity of biogas reactors due to radical changes of feedstock composition. Bioresour. Technol. 2015, 176, 56–64. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| R-1 (Cattle Inoculum) | Stage 1 Day (1–100) | Stage 2 (Days 101–108) | Stage 3 (Days 109–120) |
| Substrate B (100 mL) | Monensin (Daily I mL) | No monensin | |
| R-2 (Municipal Sanitary inoculum) | Stage 1 Days (1–37) | Stage 2 Days (37–57) | Stage 3 Days (57–90) |
| Added A and T | S:B:T | B |
| Physicochemical Composition | Poultry A | Poultry B [21] | Poultry C |
|---|---|---|---|
| TS (%) | 20 | 19 | 22 |
| VS (%) | 15 | 18 | 19 |
| pH | 7.84 | 5.54 | 7.35 |
| NH4-N | 433.58 mM | 19.86 mM | 179.16 mM |
| C:N ratio | 4:1 | 17:1 | 8:1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shah, F.A.; Mackie, R.I.; Mahmood, Q. Biogas Dynamics and Microbial Composition Employing Different Inocula and Substrates in Continuous Stirred-Tank Reactors. Microbiol. Res. 2024, 15, 174-186. https://doi.org/10.3390/microbiolres15010012
Shah FA, Mackie RI, Mahmood Q. Biogas Dynamics and Microbial Composition Employing Different Inocula and Substrates in Continuous Stirred-Tank Reactors. Microbiology Research. 2024; 15(1):174-186. https://doi.org/10.3390/microbiolres15010012
Chicago/Turabian StyleShah, Fayyaz Ali, Roderick I. Mackie, and Qaisar Mahmood. 2024. "Biogas Dynamics and Microbial Composition Employing Different Inocula and Substrates in Continuous Stirred-Tank Reactors" Microbiology Research 15, no. 1: 174-186. https://doi.org/10.3390/microbiolres15010012
APA StyleShah, F. A., Mackie, R. I., & Mahmood, Q. (2024). Biogas Dynamics and Microbial Composition Employing Different Inocula and Substrates in Continuous Stirred-Tank Reactors. Microbiology Research, 15(1), 174-186. https://doi.org/10.3390/microbiolres15010012