Abstract
Studies of newly isolated strains of lactic acid bacteria (LAB) are a good basis for expanding the potential for their applications in functional foods, probiotic food supplements, and other probiotic products. They exhibit various functional properties, including such with antiviral activity. Probiotic strains can manifest their antiviral effects by various mechanisms, including direct interaction with viruses, production of antiviral compounds, or immune system modulation. Ten newly isolated LAB strains from traditional fermented food products have been tested for the determination of their antiviral activity. This study was performed to evaluate the effect of cell-free supernatants (CFSs) from the studied strains for the effect on viral replication of Human alphaherpesvirus—HHV-1 and HHV-2 as well as for direct virucidal activity. The CFSs of the LAB strains were used in non-toxic concentrations of 25%, 6.25%, and 1.6%. No direct virucidal activity was observed in tested CFSs, but five of the strains observed a well-defined effect of viral replication inhibition with the selective index (SI) from 4.40 to >54. For two of these five strains, Lactobacillus delbrueckii subsp. bulgaricus KZM 2-11-3 and Lactiplantibacillus plantarum KC 5-12 strong activity against HHV-2 with a selective index (SI) over 45 was detected, which is a good basis for further research.
1. Introduction
Lactic acid bacteria (LAB) are widely used in areas such as the food industry, as they are used as starter cultures [1,2], bio-protectives [3], and probiotics [4,5,6]. They are used as starter cultures in fermented foods and beverages because they can improve nutritional, organoleptic, and technological characteristics as well as the shelf life of the products [7]. Probiotics have applications not only in the food industry but also in the medical and pharmaceutical industries, as they exhibit health-improving properties [8]. The probiotic effect helps the assimilation of cholesterol, lactose intolerance; controls viral, bacterial, and antibiotic diarrheal diseases, inflammatory bowel disease, allergies, atopic dermatitis, and intestinal carcinogens; stimulates the immune system on its surface; and increases resistance to infections of bacterial and viral origin. They have various mechanisms of action such as regulating intestinal microbiota and producing bioactive compounds with activity against different pathogenic bacteria, fungi, or viruses. Based on their adhesive properties, LAB prevent pathogens’ interaction with epithelial tissues. Modulation of the immune system shows capacity of the LAB to affect various diseases and induce various health benefits. Each probiotic microorganism can manifest different mechanisms of action and different health-promoting effects, which define the probiotic potential of LAB as species- and strain-specific [9,10]. LAB are the sources of multiple biologically active substances. During metabolic processes, lactic acid bacteria produce vital substances, including lactic acid, bacteriocins, amino acids, enzymes, exopolysaccharides, fatty acids, and hydrogen peroxide [11,12]. The antimicrobial potential of lactic acid is unique among living microorganisms and represents a great potential in the application, which is still underused. The effects of bacteriocins and bacteriocin-producing LAB have also been extensively studied in many test systems [13]. Hydrogen peroxide (H2O2) and lactic acid produced by lactobacilli play an important role as natural antimicrobial substances and are toxic to a number of organisms, including viruses [14].
Infection caused by members of the genus Simplex virus (Human alphaherpesvirus HHV 1 and 2) is a global health threat. Its prevalence is based on viral ability in many cases to be transmitted asymptomatically. After the primary inoculation, the virus rapidly establishes life-long latency within infected neurons [15]. All this results in a massive proportion of the world’s population living with herpes infection [16]. Latent infection is expressed in the recurrence of symptoms (painful sores and blisters around the mouth and lips (when a person is infected with HHV-1) and on the genitals and anus (HHV-2). The higher risk groups are the immunocompromised patients and neonates in whom the lack of an adequate immune response is the cause of encephalitis, meningitis, neonatal herpes, and even death. There is no drug that can eradicate the virus from the body. For this reason, the preparations used in therapy, for example, nucleoside analogs such as acyclovir (ACV) and its derivatives can only deal with the symptoms of primary and recurrent HHV infections. Their common and improper use leads to the development of resistant viral mutants [17]. This emerging resistance to antiviral agents is causing a growing concern worldwide and directs researchers’ attention to the search for alternatives in completely different fields [18]. The knowledge of the antiviral activity of lactic acid bacteria is limited and this area of research is expanding intensively.
There have been several reports of antiviral effects of metabolic by-products of environmental and intestinal bacteria, including probiotic bacteria, on enveloped and non-enveloped viruses [1,19,20,21,22,23]. In the summary according to Muhialdin et al. from the studies p by different authors, it was found that strains isolated both from fermented foods and isolates of human origin have shown that they have antiviral activity mainly against influenza viruses and enteroviruses [24]. Hotta et al. reported an antiviral effect of Enterococcus faecalis (previously known as Streptococcus faecalis) against HHV-1 and adenovirus type 12 [25]. As shown in many intervention studies, probiotic bacteria can have a beneficial effect against some human viruses such as rotavirus and HIV-induced diarrhea [26]. The cell-free supernatants (CFSs) containing the metabolites of five yogurts fermented under anaerobic conditions with Lactobacillus acidophilus, Lacticaseibacillus rhamnosus, Lactiplantibacillus plantarum, Streptococcus thermophilus, and Bifidobacterium bifidum, respectively, showed antiviral activity against seven RNA viruses (Coxsackie A virus16, coxsackie B virus 3, coxsackie B virus 4 virus, influenza virus A/PR/8/34, influenza virus A/WS/33, influenza virus B/Lee/40 and porcine epidemic diarrhea virus (PEDV) CV 777) [27]. Slight inhibition of viral propagation was observed for murine norovirus 1 and Tulane virus in the continuous presence of CFSs prepared from the growth of Bacillus subtilis 168 and Enterococcus faecalis ATCC 19433, respectively [28]. Sirichokchatchawan et al. reported that the CFSs and live LAB showed protective effects against the pandemic strain of the porcine epidemic diarrhea virus in a strain-specific manner [29].
Inspired by the above research, we considered the possibility to expand the knowledge about the antiviral activity of lactic acid bacteria and to contribute and help in the field of research for alternatives against viruses. Our previous research has shown that isolated LAB strains have good antimicrobial activity against several pathogens, bacteria, and fungi [30]. Therefore, we decided to deepen our research on these strains by testing them against human alphaherpesvirus HHV-1 and HHV-2. This study aims to determine the antiviral activity of cell-free supernatants from ten newly isolated LAB strains from different fermented foods, against HHV-1 and HHV-2.
2. Materials and Methods
2.1. Studied LAB Strains and CFS Preparation
Ten LAB strains were investigated in this study (Table 1). All these strains were newly isolated from different traditional fermented foods. The strains were previously identified as Lactiplantibacillus plantarum—five isolates KC 5-12, KC 5-14, KZC 8-21-1, KZC 8-23-5, and KO 4-4; Pediococcus pentosaceus—one isolate KC 5-13; Latilactobacillus sakei—one isolate C 10-31-3; Loigolactobacillus coryniformis—one isolate KO 3-7-5; Lactobacillus delbrueckii subsp. bulgaricus—two isolates KZM 2-11-1 and KZM 2-11-3 [30].

Table 1.
Studied strains of lactic acid bacteria from different traditional fermented foods and CFS sample numbering.
All studied strains were incubated for 24 h in de Man Rogosa, and Sharpe (MRS) broth (Merck, Darmstadt, Germany), at 41 °C; and 37 °C, thermophilic strains; and 30 °C mesophilic strains; and two strains KZM2-11-3 and KZM2-11-1 were also cultivated in 10% skimmed milk (SM) (Himedia, Mumbai, India). The cultures of all strains at concentrations of 108 CFU/mL were prepared for the next experiment by centrifugation at 6000× g for 10 min at 10 °C. Supernatants (CFS) were collected, and the pH values were measured and neutralized with 1N NaOH to pH 6.5–7. The supernatants were then filtered with a 0.20 μm syringe bacterial filter to remove any remaining bacterial cells. Cell-free supernatants (CFS) from all 24 h cultures were used for future analysis. The numbering of each sample is described in Table 1.
2.2. Cytotoxicity Test
The cytotoxicity was determined by colorimetric MTT assay [31]. Confluent monolayers of MDBK cells in 96-well plates were overlaid with 0.1 mL/well maintenance medium, 0.1 mL/well of serial two-fold dilutions of the CFSs (in maintenance medium), or 0.1 mL/well only maintenance medium (in cell controls) and were incubated at 37 °C for 48 h. On the second day, 0.02 mL of MTT (Sigma-Aldrich)—(5 mg/mL in PBS) was added to each well and the plates were incubated for 2 h at 37 °C. The optical densities (OD) were determined by a plate reader at 540 nm. Cytotoxicity was calculated by the following formula [(ODexp.)/(ODcell control)] × 100, where (ODexp.) and (ODcell control) indicate the absorptions of the test sample and the cell control, respectively. The 50% cytotoxicity concentration (CC50) was calculated by regression analysis of the dose-response curves generated from the data.
2.3. Antiviral Activity of the Bacterial Supernatant
The assessment of the potential for antiviral activity was carried out against the viral models HHV-1 and HHV-2.
2.3.1. MTT-Based Colorimetric Assay for Detection of HHV Replication Inhibition
A modification of an MTT assay [5] was used to screen anti-HHV compounds. Acyclovir was used as a reference substance (ACV) (Sigma-Aldrich) diluted in 0.5% DMSO and maintenance medium ex tempore. Confluent monolayers of MDBK cells in 96-well plates were overlaid with 0.1 mL/well of virus suspension—MOI (multiplicity of infection) = 100 TCID50/well. The plates were incubated for 1 h at 37 °C and dilutions of the CFSs or only sterile medium (as control) were added after that. Uninoculated cells were used for cell control. On day 5, p.i. plates were treated in the same way as described in the method for measuring cell viability. The percentage of protection was calculated by the following formula: [(ODexp.)—(ODvirus control)/(ODcell control)—(ODvirus control)] ×100, where (ODexp.), (ODvirus control), and (ODcell control) indicate the absorptions of the test sample, the virus control, and the cell control, respectively. The 50% effective concentration (EC50) was calculated by regression analysis of the dose-response curves generated from the data.
The selective index (SI) was calculated as CC50/EC50.
2.3.2. Virucidal Activity
The direct virus-inactivating effect of the CFSs was tested by direct contact assay. Undiluted stock virus suspensions were treated with equal volumes of the samples in MNC (maximal nontoxic concentration), prepared in a maintenance medium, and incubated at 37 °C for 5, 15, 30, 60, 120, 240, and 360 min. Undiluted stock virus suspensions were treated with equal volumes of compound-free sterile medium for control (VC). At the end of each time interval, the control and the treated viruses were frozen, and the difference in the biological activities between them was determined on the base of infectivity. The surviving infectious virus titers were determined in CPE assay using the method of Reed and Muench [31].
2.4. Data Analyses
The experiments were performed in triplicate. Dose-dependent curves of the results as mean along with calculated standard deviation were built using Origin Pro software.
3. Results
3.1. Cytotoxicity
Prior to an antiviral activity study, the cytotoxicity of CFSs was determined against the used cell culture. Cell growth, morphological effects, and activity over cellular mitochondrial metabolism were determined [32]. Twelve studied CFSs from lactic acid bacteria were used between concentrations of 100% and 1.5% using two-fold dilutions (Figure 1) on the MDBK cell line. All of them showed a dose-depending effect over the used cell line. The maximal tolerance concentration (MTC) was determined microscopically based on morphological changes in the monolayer. All studied CFSs have no visible morphological effect over MDBK culture below 25% concentration.
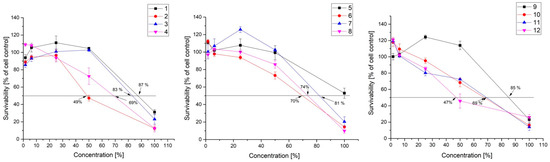
Figure 1.
The cytotoxicity of samples of CFSs from the studied LAB strains 1. KC 5-12, 2. KC 5-13, 3. KC 5-14, 4. KZC 8-21-1, 5. KZC 8-23-5, 6. C 10-31-3, 7. KO 4-4, 8. KO 3-7-5, 9. KZM 2-11-3, 10. KZM 2-11-1, 11. KZM 2-11-3 (SM), 12. KZM 2-11-1 (SM) on the MDBK cell line.
Dose-response curves were generated based on the cytotoxicity data and the CC50 concentration was calculated for 11 from 12 CFSs. Only KZC 8-23-5 even in 100% concentration did not achieve 50% cytotoxicity (47%). The CC50 value is between 47% and 85% for all other CFSs.
The supernatants with the highest toxicity are KZM 2-11-1 (SM) with CC50 of 47% and KC 5-13 with 49%. Most CFSs have CC50 around 70%—KZC 8-21-1, KZM 2-11-1, KZM 2-11-3 (SM media) (69%), C 10-31-3 (70%), and KO 3-7-5 (74%). With lower toxicity are KO 4-4 (81%), KC 5-14 (83%), KZM 2-11-3 (85%), and KC 5-12 (87%).
It is interesting that cultivation in SM media seems to increase the toxic activity of the CFSs against used cell culture. For KZM 2-11-3 CC50 increases from 85% to 69% and for KZM 2-11-1—69% to 47%. This can be based on the growth medium of LAB cultivation. According to Mani-Lopez, one of the factors that contribute to the CFS content is the growth medium which can conclude with the production of different components from the same strain depending on the medium where they are cultivated [33].
All studied CFSs exhibit overall low cytotoxicity over epithelial cells (Figure 1). The data are confirmed by other similar studies that have been based on alike cell lines—Vero and MDCK [27].
3.2. Evaluation of HHV Replication Inhibition from the Bacterial CFSs
The antiviral activity is based on the sequential administration of viral stock and the bacterial CFSs over the cells. With this, the virus has time to initiate its life cycle and the antiviral activity will be limited to the intracellular part of the viral replication.
All 12 of the CFSs were used in non-toxic concentrations of 25%, 6.25%, and 1.6%. Dose-response curves are built (Figure 2) and the 50% effective concentration (EC50) is calculated.
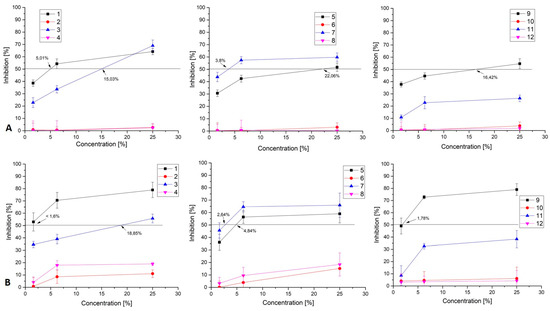
Figure 2.
Antiviral activity of CFSs from the studied LAB strains 1. KC 5-12, 2. KC 5-13, 3. KC 5-14, 4. KZC 8-21-1, 5. KZC 8-23-5, 6. C 10-31-3, 7. KO 4-4, 8. KO 3-7-5, 9. KZM 2-11-3, 10. KZM 2-11-1, 11. KZM 2-11-3 (SM), 12. KZM 2-11-1 (SM) against (A) HHV 1 and (B) HHV 2.
Five CFSs exhibit antiviral activity over the replication of both viral strains. The CFSs that exhibit antiviral activity belong to the strains KC 5-12, KC 5-14, KZC 8-23-5, KO 4-4, and KZM 2-11-3. KC 5-12 has EC50 of 5% against HHV-1 and under 1.6% against HHV-2, thus making it the most effective agent of all twelve tested with SI over 54 (see Table 2). KC 5-14 has a much lesser efficiency with EC50 of 15% and 19%, respectively. In it, MTC showed the highest antiviral capacity with nearly 70% inhibition of the viral replication and a very clear dose-depended effect.
Table 2.
Antiviral activity of CFSs from the studied LAB strains HHV-1 and HHV-2 and determination of selective index.
Against HHV-1 CFSs with the strongest effect is probably KO 4-4 with EC50 of 3.8%, which results in SI over 21.
The observed effect is purely based on the activity over the replication cycle of the virus inside the host cells, as with one hour of incubation the observed results cannot contribute to the effect over viral adsorption/penetration or any other form of virucidal effect previously described [14,34]. Overall, the dose-dependent antiviral effect is observed on similar products [31,34,35] where authors propose a multi-targeted effect from the complex of compounds, produced in CFSs.
3.3. Selective Index (SI)
The selective index (SI) of the studied CFSs for strains which exhibit antiviral activity over the replication of both viral types was determined based on their cytotoxicity (50% cytotoxic concentration, CC50) and inhibition (50% effective concentration, EC50) against both viral strains (Table 2).
Four of the CFSs from strains KZM 2-11-3, KC 5-12, KC 5-14, and KO 4-4 show a selective index over fifteen against one or both viral models. Although these established antiviral activities are lower compared to commercially available products, such as acyclovir with SI 560 (determined in parallel analysis by the described method), they can have practical importance.
The high SI values, determinate for some CFSs from LAB strains in this study, support the thesis of other researchers as well, that the antiviral effect is probably due to the multi-targeted activity of the complex of various biologically active substances in CFSs, which were produced by LABs [12,13,14,34,35,36].
3.4. Virucidal Activity
The CFSs from two of the studied strains, KC 5-12 and KZM2-11-3, which have shown the highest activity against HHV-2, have been selected and their virucidal potential against the same viral model was investigated. Over 6 h of a direct inactivation experiment, both samples showed very low activity against the virions of the HHV-2 (Figure 3). No more than 0.5 log difference (68.3%) between samples and viral control was observed. The determination of the virucidal effect is applied to evaluate the activity during the early stages of the virus life cycle (binding and entry) or directly on the virion and is usually based on the activity of such metabolites as hydrogen peroxide or acids produced by LAB strains [14,36]. The lack of virucidal effect of the two selected samples supports the thesis that the established antiviral activity during replication of the viral model is determined by other molecular mechanisms and by the complex of produced biologically active substances.
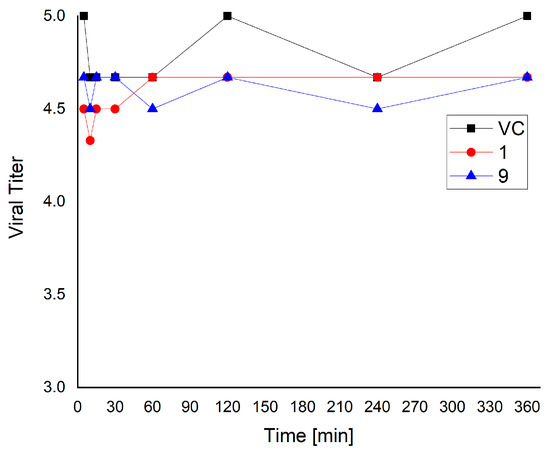
Figure 3.
Virucidal activity of CFSs from KC5-12 and KZM2-11-3 against HHV-2.
4. Discussion and Conclusions
Some of the lactic acid bacteria such as L. rhamnosus [36,37], Lactococcus lactis subsp. cremoris [37], and Lactobacillus gasseri [18] can affect HHV-1 and L. brevis can affect HHV-2 [31]. According to Khani et al., L. rhamnosus was able to elicit some antiviral effects against HHV-1, which may be through various mechanisms including competition with the virus for cell surface adhesion and increasing macrophage stability by 90%, after stimulation of pro-inflammatory responses [36]. The antiviral activity of the bacterial extract and cell wall fragments obtained from L. brevis cells after removal of the S layer was significantly reduced compared to untreated cells, suggesting that the inhibitory activity is likely to be due to a bacterial component resistant to the heat of the surface of cells [31]. Lactobacillus crispatus have an antiviral effect against HHV-2 (one of the most prevalent etiologies of sexually transmitted diseases, a risk factor for acquiring HIV) in a mammalian cell line according to two loading mechanisms suggested by authors: (1) the formation of L. crispatus microcolonies in the cell surface could block HHV-2 receptors and prevent viral entry to cells in initial infection steps; and (2) entrapment of viral particles by direct interaction of L. crispatus with HHV-2 envelope [38]. Exopolysaccharide from L. plantarum LRCC5310 offers protection against rotavirus-induced diarrhea and regulates inflammatory response [39]. Exopolysaccharides secreted during LAB culture, in addition to their probiotic activity, can act selectively on pathogenic microbes and viruses [40]. Cyclic dipeptides from lactic acid bacteria inhibit the proliferation of the influenza A virus [41]. According to the study performed by Yang et al., LAB may have the potential to prevent PEDV infection in pigs and restore the damaged intestinal to healthy conditions [42]. The lactic acid bacteria that stand out the most for their antiviral activity against various viruses are mainly L. plantarum, L. rhamnosus, L. gasseri, L. delbrueckii subsp. Bulgaricus, Bifidobacterium sp., Streptococcus thermophilus, etc. [43].
In the present study, all studied CFSs exhibit overall low cytotoxicity over epithelial cells. Ongoing, it was established that CFSs from five LAB strains exhibit antiviral activity over the replication of both viral models HHV-1 and HHV-2 and show a selective index over fifteen against one or both viral models. Four of the strains, KC 5-12, KC 5-14, KZC 8-23-5, and KO 4-4, that exhibit antiviral activity belong to the species Lactiplantibacillus plantarum, and one strain, KZM 2-11-3, belongs to Lactobacillus delbrueckii subsp. bulgaricus. The CFSs from the strains KC 5-12 and KZM2-11-3 have shown the highest activity against HHV-2, respectively, >54 and 47.75. Other authors have also described the antiviral activity of the strains of these two species Lactiplantibacillus plantarum and Lactobacillus delbrueckii subsp. bulgaricus, which have shown activity against influenza viruses, as well as other viruses that have caused high mortality in the population. L. plantarum KAU007 has shown strong antiviral activity against the H1N1 virus [44], and L. plantarum Probio-88 inhibits the replication activity of the SARS-COV-2 [45]. L. delbrueckii can inhibit virus infection of influenza A virus by stimulating the immune system and by inhibiting the virus [46,47]. Vilhelmova-Ilieva et al. reported that Lactiplantibacillus plantarum inhibit the replication of HHV-1 with a high selective index [34]. In this study, it was established for the first time that the strain KZM 2-11-3 of the species Lactobacillus delbrueckii subsp. bulgaricus inhibits the replication of HHV-1 and HHV-2 and has a high selective index at HHV-2 (Table 2), which makes this strain have the probiotic potential and applicable for the prevention of viral infections. Except this, the strain KC 5-12 of the species Lactiplantibacillus plantarum inhibits the replication of both HHV-1 and HHV-2, with a high selective index for HHV-2. It is important to note that the use of probiotics over decades has shown that they are safe and have beneficial effects on the user. Based on the obtained results, lactic acid bacteria can be an alternative for improving the quality of life of patients suffering from viral infections. They can contribute to the inhibition of the replication of viruses present in the human body by the complex of different antiviral components produced by these bacteria, as well as to the improved response of the immune system against viral infections. Lactic acid bacteria can be considered an alternative treatment as they have a wide range of the action of inhibiting viruses ranging from respiratory to sexually transmitted viruses [48,49,50]. Although the mechanism of action of probiotic strains to inhibit viral replication is not yet fully understood, the attention of the scientific and medical communities is now increasingly focused on the benefits that will come from supporting probiotics in the fight against viruses. Researchers have proven that probiotics were employed to prevent and reduce symptoms of infections caused by common viruses, especially respiratory tract viruses [10,51], but also for herpes simplex virus [43]. LAB may exert their antiviral activity by direct probiotic virus interaction; the production of antiviral inhibitory metabolites; and via immune system stimulation [18]. Furthermore, conducting more in-depth studies on the application of lactic acid bacteria for prevention or co-therapy of viral infections is necessary.
Author Contributions
Conceptualization, D.N., D.T. and R.H.; methodology, D.T. and K.S.; formal analysis, R.H., D.N., Y.E. and A.H.; writing—original draft preparation, R.H., D.N. and D.T.; writing—review and editing, D.N., D.T., A.H., Y.E. and K.S.; supervision, D.N. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Bulgarian Ministry of Education and Science under the NRP “Healthy Foods for a Strong Bio-Economy and Quality of Life” approved by DCM # 577 /17.08.2018.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
This work is partially supported by the Operational Program “Science and Education for Smart Growth”, Bulgaria, grant number BG05M2OP001-1.002-0012.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhou, T.; Huo, R.; Lai-Yu, K.; Li, C.; Ma, Y.; Mi, Z.; Chen, Y. Effects of applying Lactobacillus helveticus H9 as adjunct starter culture in yogurt fermentation and storage. J. Dairy Sci. 2019, 102, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.N.; Frank, J.F. Starter Cultures and Their Use. In Applied Dairy Microbiology, 2nd ed.; revised and expanded; Marth, E.H., Steele, J.L., Eds.; CRC Press: Boca Raton, FL, USA, 2001; Volume 6, p. 736. [Google Scholar]
- Russo, P.; Fares, C.; Longo, A.; Spano, G.; Capozzi, V. Lactobacillus plantarum with Broad Antifungal Activity as a Protective Starter Culture for Bread Production. Foods 2017, 6, 110. [Google Scholar] [CrossRef] [PubMed]
- Bintsis, T. Lactic acid bacteria: Their applications in foods. J. Bacteriol. Mycol. 2018, 6, 89–94. [Google Scholar]
- Bai, M.; Huang, T.; Guo, S.; Wang, Y.; Wang, J.; Kwok, L.; Dan, T.; Zhang, H.; Bilige, M. Probiotic Lactobacillus casei Zhang improved the properties of stirred yogurt. Food Biosci. 2020, 37, 100718. [Google Scholar] [CrossRef]
- Jing, W.; Haifeng, J.; Sixin, W.; Hui, L.; Wei, Z.; Dongyan, Z.; Yamin, W. Probiotic Lactobacillus plantarum Promotes Intestinal Barrier Function by Strengthening the Epithelium and Modulating Gut Microbiota. Front. Microbiol. 2018, 9, 1953. [Google Scholar]
- Smit, G.; Smit, B.A.; Engels, W.J.M. Flavour formation by lactic acid bacteria and biochemical flavour profiling of cheese products. FEMS Microbiol. Rev. 2005, 29, 591–610. [Google Scholar] [CrossRef]
- Ramzi, A.; Alsaheb, A.; Aladdin, A.; Othman, N.Z.; Malek, R.M.; Leng, O.M.; Aziz, R.; El Enshasy, H.A. Lactic acid applications in pharmaceutical and cosmeceutical industries. J. Chem. Pharm. Res. 2015, 7, 729–735. [Google Scholar]
- Ren, C.; Faas, M.M.; de Vos, P. Disease managing capacities and mechanisms of host effects of lactic acid bacteria. Critical Rev. Food Sci. Nutr. 2021, 61, 1365–1393. [Google Scholar] [CrossRef]
- Tiwari, S.K.; Dicks, L.M.T.; Popov, I.V.; Karaseva, A.; Ermakov, A.M.; Suvorov, A.; Tagg, J.R.; Weeks, R.; Chikindas, M.L. Probiotics at War Against Viruses: What Is Missing from the Picture? Front. Microbiol. 2020, 11, 1877. [Google Scholar] [CrossRef]
- Gaspar, C.; Donders, G.G.; Palmeira-de-Oliveira, R.; Queiroz, J.A.; Tomaz, C.; Martinez-de-Oliveira, J.; Palmeira-de-Oliveira, A. Bacteriocin production of the probiotic Lactobacillus acidophilus KS400. AMB Expr. 2018, 8, 153. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, X.; Pan, W.; Shen, X.; He, Y.; Yin, H.; Zhou, K.; Zou, L.; Chen, S.; Liu, S. Exopolysaccharides produced by yogurt-texture improving Lactobacillus plantarum RS20D and the immunoregulatory activity. Int. J. Biol. Macromol. 2019, 121, 342–349. [Google Scholar] [CrossRef]
- Nes, I.F.; Kjos, M.; Diep, D.B. Antimicrobial Compounds of Lactic Acid Bacteria. In Lactic Acid Bacteria Microbiological and Functional Aspects, 4th ed.; Lahtinen, S., Ouwehand, A.C., Salminen, S., von Wright, A., Eds.; CRC Press: Boca Raton, FL, USA, 2012; Volume 5, pp. 285–329. [Google Scholar]
- Conti, C.; Malacrino, C.; Mastromarino, P. Inhibition of herpes simplex virus type 2 by vaginal lactobacilli. J. Physiol. Pharmacol. 2009, 6, 19–26. [Google Scholar]
- Akhtar, J.; Shukla, D. Viral entry mechanisms: Cellular and viral mediators of herpes simplex virus entry. FEBS J. 2009, 276, 7228–7236. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.who.int/news-room/fact-sheets/detail/herpes-simplex-virus (accessed on 10 March 2022).
- Morfin, F.; Thouvenot, D. Herpes simplex virus resistance to antiviral drugs. J. Clin. Virol. 2003, 26, 29–37. [Google Scholar] [CrossRef]
- Al Kassaa, I.; Hober, D.; Hamze, M.; Chihib, N.E.; Drider, D. Antiviral potential of lactic acid bacteria and their bacteriocins. Probiotics Antimicrob. Proteins 2014, 6, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Carver, D.H.; Naficy, K. Inhibition of arbor viruses (group A) by a protein-like constituent of a Corynebacterium. Proc. Soc. Exp. Biol. Med. 1962, 111, 356–360. [Google Scholar] [CrossRef]
- Cliver, D.O.; Herrmann, J.E. Proteolytic and microbial inactivation of enteroviruses. Water Res. 1972, 6, 797–805. [Google Scholar] [CrossRef]
- Deng, M.Y.; Cliver, D.O. Inactivation of poliovirus type 1 in mixed human and swine wastes and by bacteria from swine manure. Appl. Environ. Microbiol. 1992, 58, 2016–2021. [Google Scholar] [CrossRef]
- Deng, M.Y.; Cliver, D.O. Antiviral effects of bacteria isolated from manure. Microb. Ecol. 1995, 30, 43–54. [Google Scholar] [CrossRef]
- Munoz, J.A.; Chenoll, E.; Casinos, B.; Bataller, E.; Ramon, D.; Genoves, S.; Montava, R.; Ribes, J.M.; Buesa, J.; Fabrega, J.; et al. Novel probiotic Bifidobacterium longum subsp. infantis CECT 7210 strain active against rotavirus infections. Appl. Environ. Microbiol. 2011, 77, 8775–8783. [Google Scholar] [CrossRef]
- Muhialdin, B.J.; Zawawi, N.; Abdul Razis, A.F.; Bakar, J.; Zarei, M. Review Antiviral activity of fermented foods and their probiotics bacteria towards respiratory and alimentary tracts viruses. Food Control 2021, 127, 108140. [Google Scholar] [CrossRef]
- Hotta, S.; Kojima, M.; Fujisaki, M.; Uchida, S.; Kuroda, H.; Hamada, C. An antiviral substance extracted from Streptococcus faecalis. Nature 1977, 268, 733–734. [Google Scholar] [CrossRef] [PubMed]
- Botić, T.; Danø, T.; Weingartl, H.; Cencič, A. A novel eukaryotic cell culture model to study antiviral activity of potential probiotic bacteria. Int. J. Food Microbiol. 2007, 115, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J.; Song, J.H.; Ahn, Y.J.; Baek, S.H.; Kwon, D.H. Antiviral activities of cell-free supernatants of yogurts metabolites against some RNA viruses. Eur. Food Res. Technol. 2009, 228, 945–950. [Google Scholar] [CrossRef]
- Shearer, A.E.; Hoover, D.G.; Kniel, K.E. Effect of bacterial cell-free supernatants on infectivity of norovirus surrogates. J. Food Prot. 2014, 77, 145–149. [Google Scholar] [PubMed]
- Sirichokchatchawan, W.; Temeeyasen, G.; Nilubol, D.; Prapasarakul, N. Protective effects of cell-free supernatant and live lactic acid bacteria isolated from Thai pigs against a pandemic strain of porcine epidemic diarrhea virus. Probiotics Antimicrob. Proteins 2018, 10, 383–390. [Google Scholar] [CrossRef]
- Hoxha, R.; Evstatieva, Y.; Nikolova, D. New lactic acid bacterial strains from traditional fermented foods—Bioprotective and probiotic potential. J. Chem. Technol. Metall. 2023, 58, 252–269. [Google Scholar]
- Mastromarino, P.; Cacciotti, F.; Masci, A.; Mosca, L. Antiviral activity of Lactobacillus brevis towards herpes simplex virus type 2: Role of cell-wall associated components. Anaerobe 2011, 17, 334–336. [Google Scholar] [CrossRef]
- Li, W.; Zhou, J.; Xu, Y. Study of the in vitro cytotoxicity testing of medical devices. Biomed. Rep. 2015, 3, 617–620. [Google Scholar] [CrossRef]
- Mani-López, E.; Arrioja-Bretón, D.; López-Malo, A. The impacts of antimicrobial and antifungal activity of cell-free supernatants from lactic acid bacteria in vitro and foods. Compr. Rev. Food Sci Food Saf. 2022, 21, 604–641. [Google Scholar] [CrossRef]
- Vilhelmova-Ilieva, N.; Atanasov, G.; Simeonova, L.; Dobreva, L.; Mancheva, K.; Trepechova, M.; Danova, S. Anti-herpes virus activity of lactobacillus’ postbiotics. BioMedicine 2022, 12, 21. [Google Scholar] [CrossRef]
- Möller, N.P.; Scholz-Ahrens, K.E.; Roos, N.; Schrezenmeir, J. Bioactive peptides and proteins from foods: Indication for health effects. Eur. J. Nutr. 2008, 47, 171–182. [Google Scholar] [CrossRef]
- Khani, S.; Motamedifar, M.; Golmoghaddam, H.; Hosseini, H.M.; Hashemizadeh, Z. In vitro study of the effect of a probiotic bacterium Lactobacillus rhamnosus against herpes simplex virus type 1. Braz. J. Infect. Dis. 2012, 16, 129–135. [Google Scholar] [PubMed]
- Cirrincione, S.; Luganini, A.; Lamberti, C.; Manfredi, M.; Cavallarin, L.; Giuffrida, M.G.; Pessione, E. Donkey Milk Fermentation by Lactococcus lactis subsp. cremoris and Lactobacillus rhamnosus Affects the Antiviral and Antibacterial Milk Properties. Molecules 2021, 26, 5100. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, E.; Makvandi, M.; Teimoori, A.; Ataei, A.; Ghafari, S.; Samarbaf-Zadeh, A. Antiviral effects of Lactobacillus crispatus against HSV-2 in mammalian cell lines. J. Chin. Med. Assoc. 2018, 81, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Lee, G.; Thanh, H.D.; Kim, J.; Konkit, M.; Yoon, S.; Park, M.; Yang, S.; Park, E.; Kim, W. Exopolysaccharide from Lactobacillus plantarum LRCC5310 offers protection against rotavirus-induced diarrhea and regulates inflammatory response. J. Dairy Sci. 2017, 101, 5702–5712. [Google Scholar] [CrossRef]
- Biliavska, L.; Pankivska, Y.; Povnitsa, O.; Zagorodnya, S. Antiviral Activity of Exopolysaccharides Produced by Lactic Acid Bacteria of the genera Pediococcus, Leuconostoc and Lactobacillus against Human Adenovirus Type 5. Medicina 2019, 55, 519. [Google Scholar] [CrossRef]
- Kwak, M.; Liu, R.; Kwon, J.; Kim, M.; Kim, A.H.; Kang, S. Cyclic Dipeptides from Lactic Acid Bacteria Inhibit Proliferation of the Influenza A Virus. J. Microbiol. 2013, 51, 836–843. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Li, S.; Lu, Y.; Jansen, C.h.A.; Savelkoul, H.F.J.; Liu, G. Oral administration of Lactic acid bacteria inhibits PEDV infection in young piglets. Virology 2023, 579, 1–8. [Google Scholar] [CrossRef]
- Lopez-Santamarina, A.; Lamas, A.; del Carmen Mondragón, A.; Cardelle-Cobas, A.; Regal, P.; Rodriguez-Avila, J.A.; Miranda, J.M.; Franco, C.M.; Cepeda, A. Probiotic Effects against Virus Infections: New Weapons for an Old War. Foods 2021, 10, 130. [Google Scholar] [CrossRef]
- Rather, I.A.; Kamli, M.R.; Sabir, J.S.M.; Paray, B.A. Potential Antiviral Activity of Lactiplantibacillus plantarum KAU007 against Influenza Virus H1N1. Vaccines 2022, 10, 456. [Google Scholar] [CrossRef]
- Rather, I.A.; Choi, S.-B.; Kamli, M.R.; Hakeem, K.R.; Sabir, J.S.M.; Park, Y.-H.; Hor, Y.-Y. Potential Adjuvant Therapeutic Effect of Lactobacillus plantarum Probio-88 Postbiotics against SARS-COV-2. Vaccines 2021, 9, 1067. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, E.; Sawabuchi, T.; Kimoto, T.; Sakai, S.; Kido, H. Lactobacillus delbrueckii ssp. bulgaricus OLL1073R-1 feeding enhances humoral immune responses, which are suppressed by the antiviral neuraminidase inhibitor oseltamivir in influenza A virus–infected mice. J. Dairy Sci. 2019, 102, 9559–9569. [Google Scholar] [CrossRef] [PubMed]
- Serkedjieva, J.; Danova, S.; Ivanova, I. Anti-influenza virus activity of a bacteriocin produced by Lactobacillus delbrueckii. Appl. Biochem. Biotechnol. 2000, 88, 285–298. [Google Scholar] [CrossRef]
- Wahab, S.; Almaghaslah, D.; Mahmood, S.E.; Ahmad, M.F.; Alsayegh, A.A.; Abu Haddash, Y.M.; Rahman, M.A.; Ahamd, I.; Ahmad, W.; Khalid, M.; et al. Pharmacological Efficacy of Probiotics in Respiratory Viral Infections: A Comprehensive Review. J. Pers. Med. 2022, 12, 1292. [Google Scholar] [CrossRef] [PubMed]
- Lehtoranta, L.; Pitkäranta, A.; Korpela, R. Probiotics in respiratory virus infections. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 1289–1302. [Google Scholar] [CrossRef] [PubMed]
- Martín, V.; Maldonado, A.; Fernández, L.; Rodríguez, J.M.; Connor, R.I. Inhibition of human immunodeficiency virus type 1 by lactic acid bacteria from human breastmilk. Breastfeed Med. 2010, 5, 153–158. [Google Scholar] [CrossRef]
- Chourasia, R.; Padhi, S.; Chiring Phukon, L.; Abedin, M.M.; Singh, S.P.; Rai, A.K. A Potential Peptide from Soy Cheese Produced Using Lactobacillus delbrueckii WS4 for Effective Inhibition of SARS-CoV-2 Main Protease and S1 Glycoprotein. Front. Mol. Biosci. 2020, 7, 601753. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).