
Analysis of the Virulence and Inflammatory Markers Elicited by Enteroaggregative Escherichia coli Isolated from Clinical and Non-Clinical Sources in an Experimental Infection Model, India
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals Used
2.3. Selection of EAEC Strains
2.4. Defining EAEC Strains
2.5. Infection of the Mice
2.6. Mice Colonization Experiments
2.7. Analysis of Inflammation Markers
2.8. Analysis of Stool for Bacterial Burden by Real-Time PCR
2.9. Detection of Virulence Factors by M-PCR
2.10. Phylogenetic Analysis via M-PCR
2.11. Histopathological Analysis
2.12. Statistical Analysis
3. Results
3.1. Mice Colonization Experiment
3.2. Virulence-Related Markers and Phylogeny among EAEC Isolate from Clinical and Non-Clinical Sources Used in the Animal Model
3.3. Analysis of Inflammatory Markers
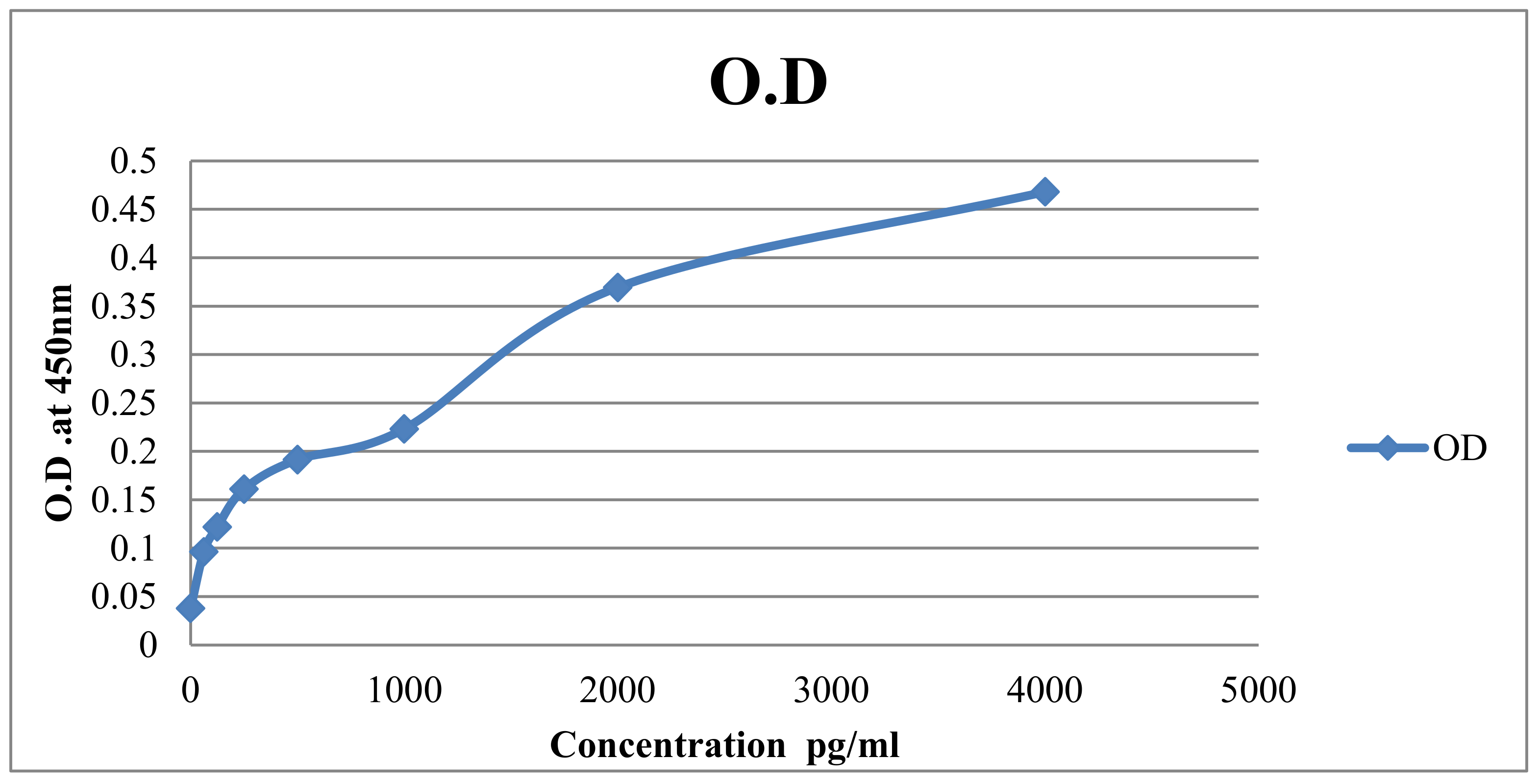
3.3.1. Standard Curve
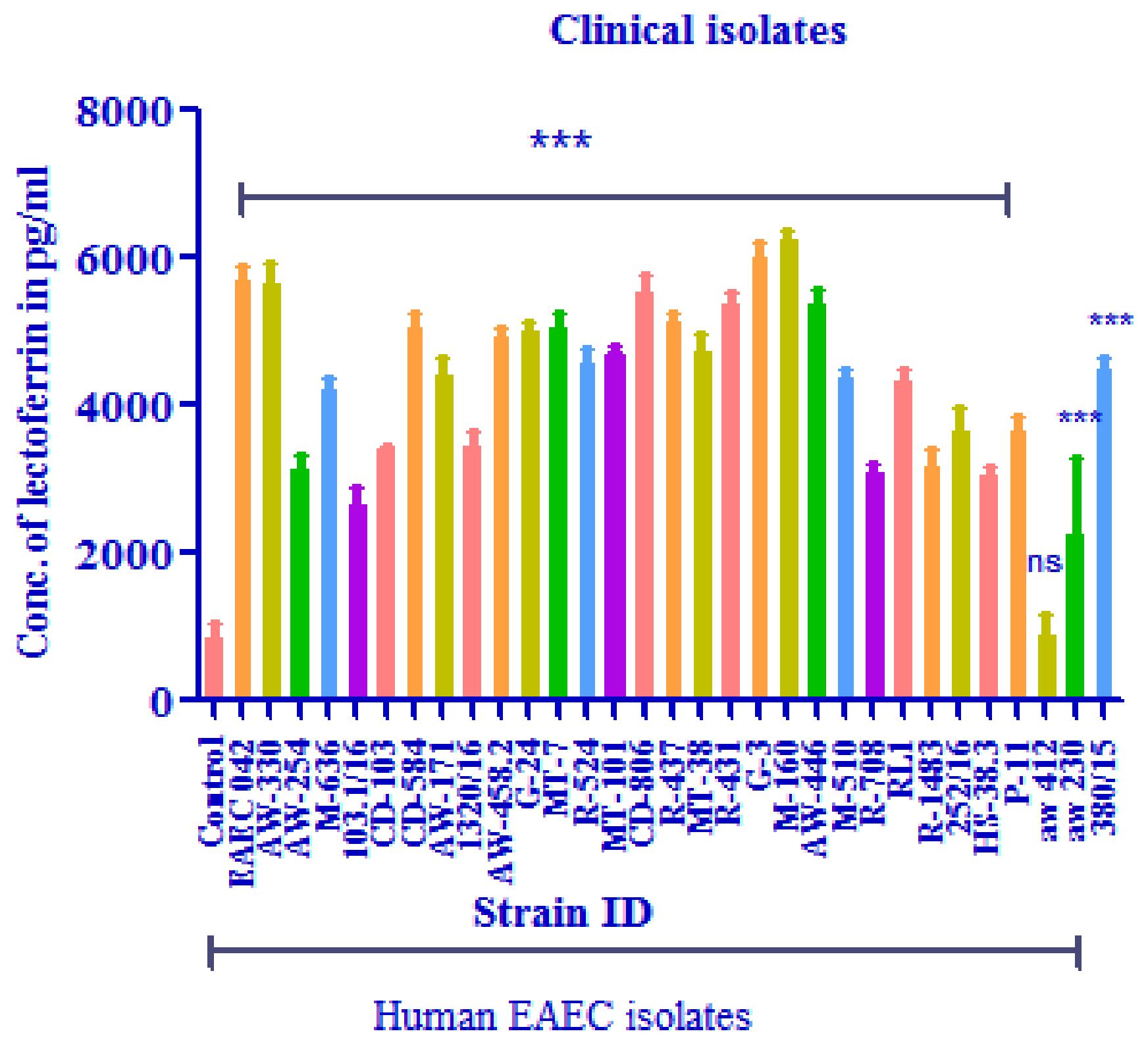
3.3.2. Lactoferrin Levels
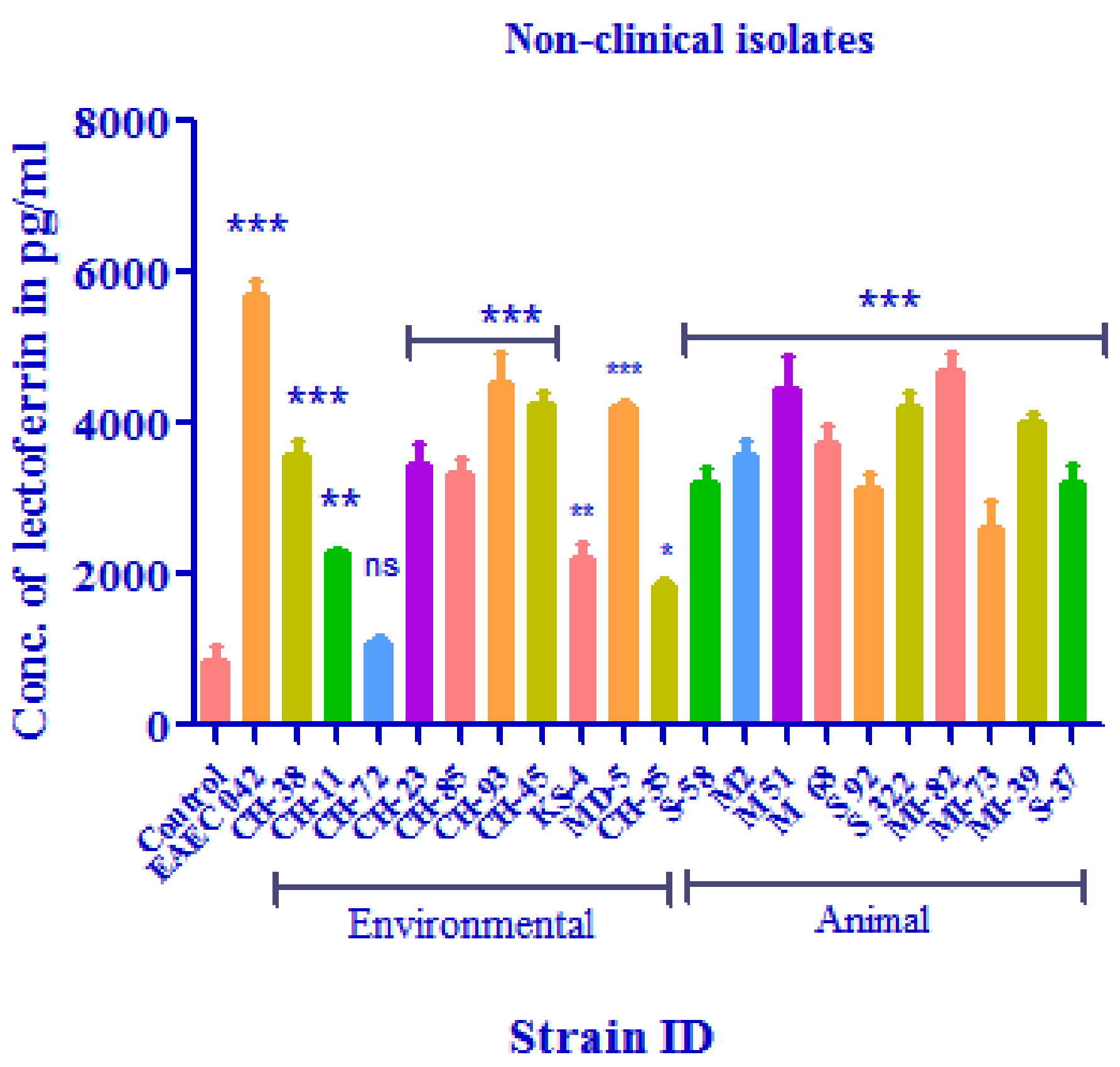
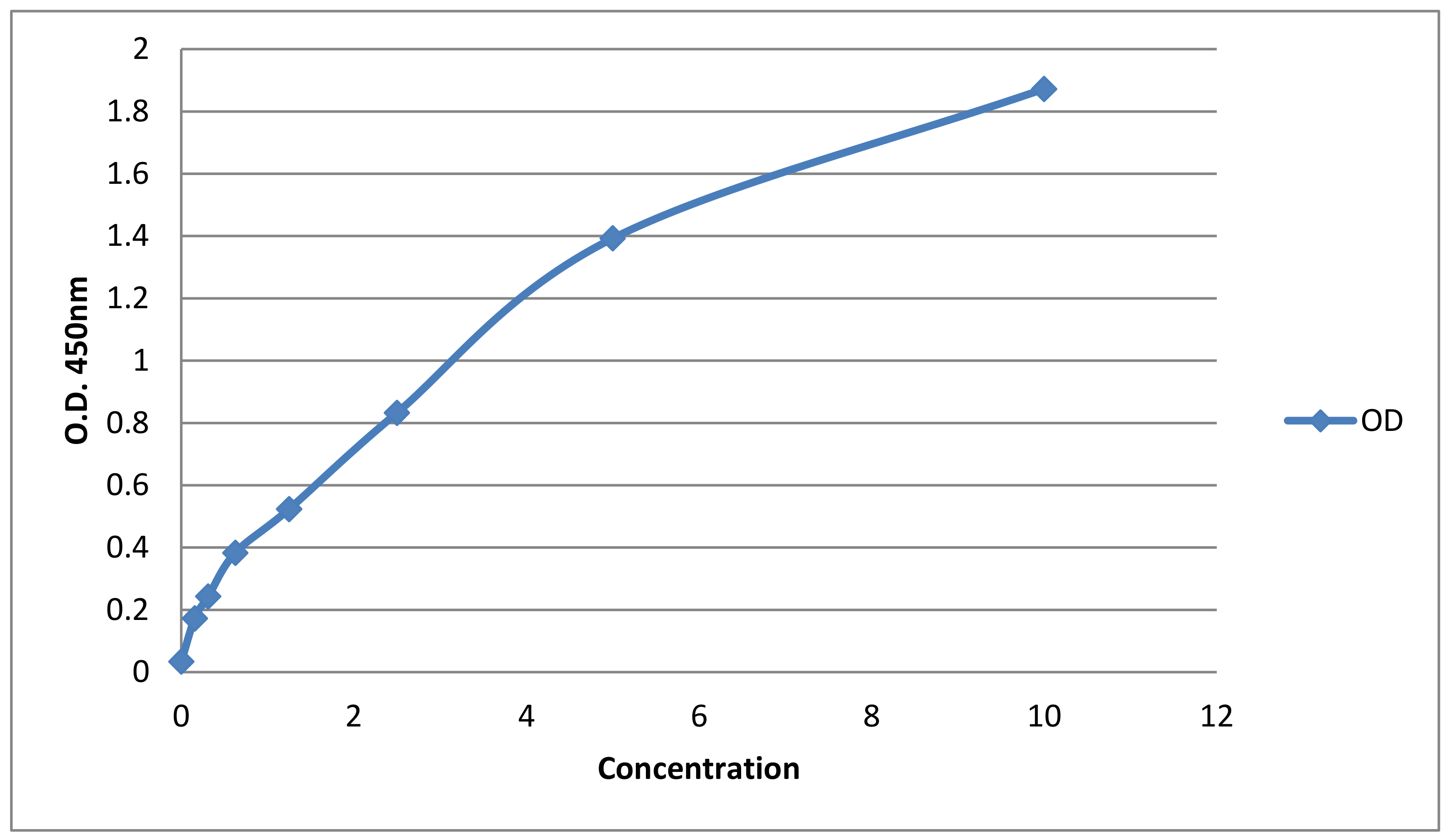
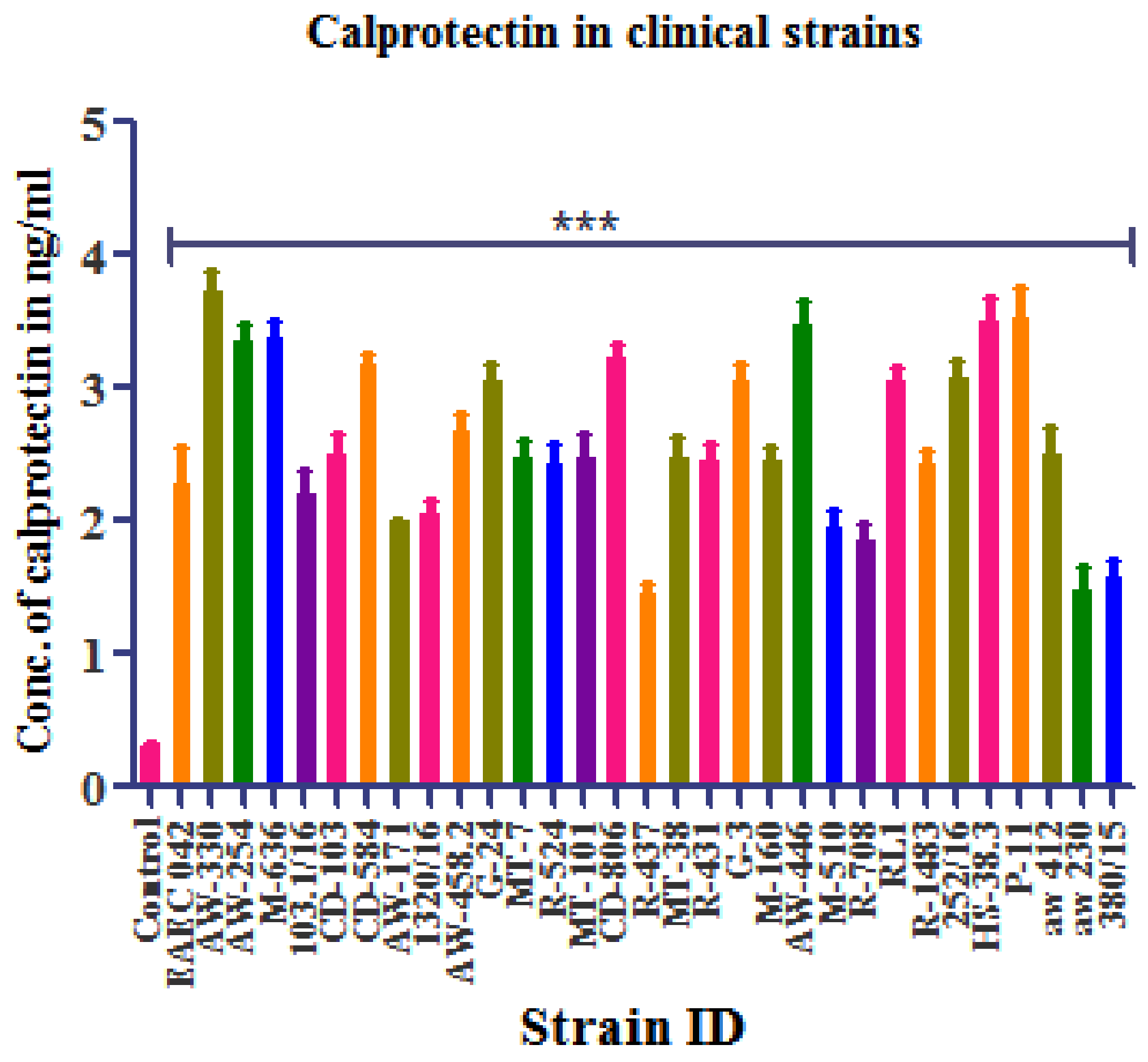
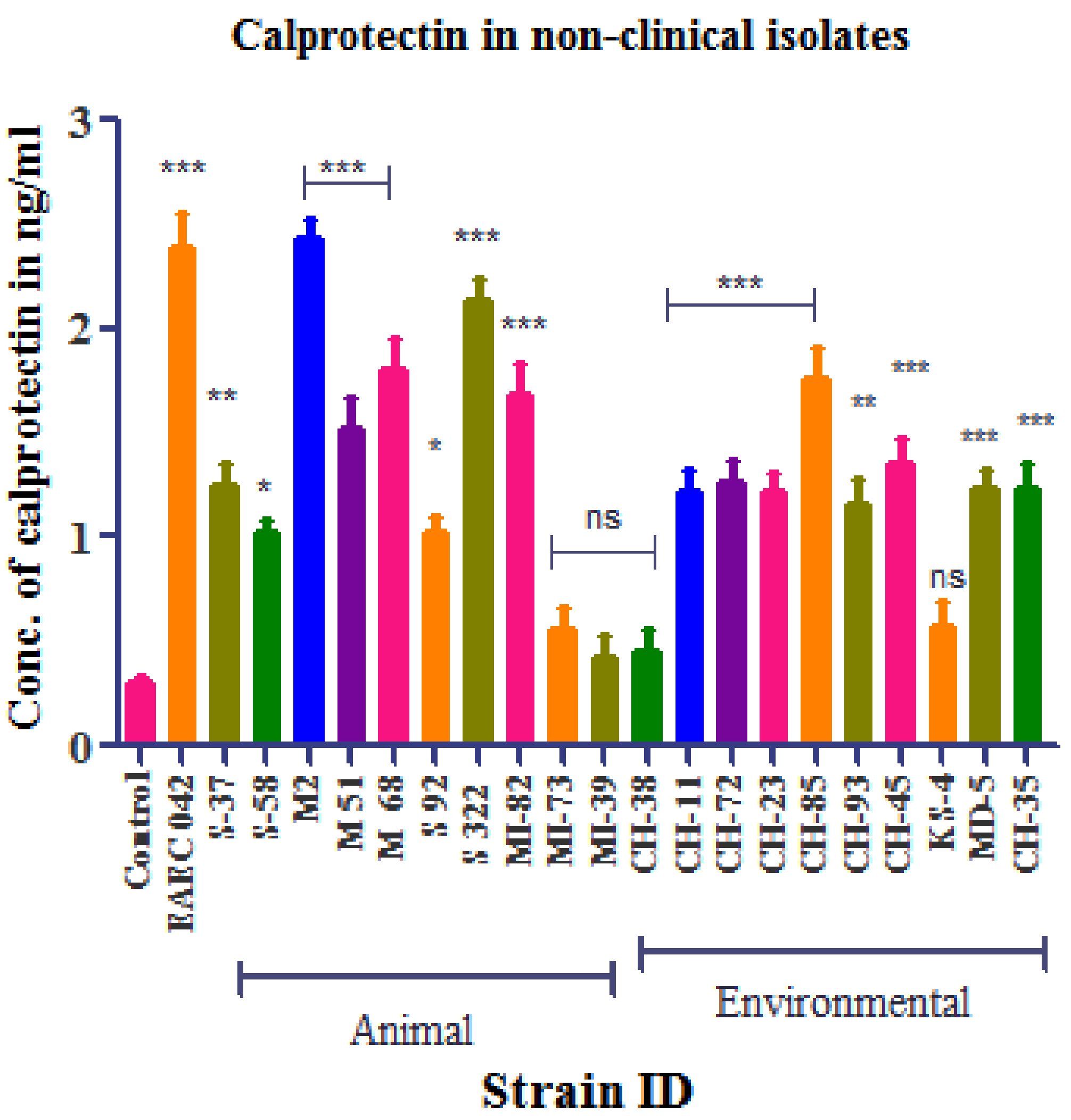
3.4. Calprotectin Detection
Standard Curve
3.5. Association of EAEC with Intestinal Tissue
3.6. Histopathological Analysis:
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elias, W.P.; Navarro-Garcia, F. Enteroaggregative Escherichia coli (EAEC). Escherichia coli Am. 2016. [Google Scholar] [CrossRef]
- Huang, D.B.; Mohanty, A.; DuPont, H.L.; Okhuysen, P.C.; Chiang, T. A review of an emerging enteric pathogen: Enteroaggregative Escherichia coli. J. Med. Microbiol. 2006, 55, 1303–1311. [Google Scholar] [CrossRef] [PubMed]
- Modgil, V.; Mahindroo, J.; Narayan, C.; Kalia, M.; Yousuf; Shahi, V.; Koundal, M.; Chaudhary, P.; Jain, R.; Sandha, K.S.; et al. Comparative analysis of virulence determinants, phylogroups, and antibiotic susceptibility patterns of typical versus atypical enteroaggregative, E. coli in India. PLoS Negl. Trop. Dis. 2020, 14, e0008769. [Google Scholar] [CrossRef] [PubMed]
- Roche, J.K.; Cabel, A.; Sevilleja, J.; Nataro, J.; Guerrant, R.L. Enteroaggregative Escherichia coli (EAEC) impairs growth while malnutrition worsens EAEC infection: A novel murine model of the infection malnutrition cycle. J. Infect. Dis. 2010, 202, 506–514. [Google Scholar] [CrossRef]
- Mathewson, J.J.; Jiang, Z.D.; Zumla, A.; Chintu, C.; Luo, N.; Calamari, S.R.; Genta, R.M.; Steephen, A.; Schwartz, P.; DuPont, H.L. HEp-2 cell-adherent Escherichia coli in patients with human immunodeficiency virus-associated diarrhea. J. Infect. Dis. 1995, 171, 1636–1639. [Google Scholar] [CrossRef]
- Huang, D.B.; Mohamed, J.A.; Nataro, J.P.; DuPont, H.L.; Jiang, Z.-D.; Okhuysen, P.C. Virulence characteristics and the molecular epidemiology of enteroaggregative Escherichia coli isolates from travellers to developing countries. J. Med. Microbiol. 2007, 56, 1386–1392. [Google Scholar] [CrossRef]
- Modgil, V.; Chaudhary, P.; Bharti, B.; Mahindroo, J.; Yousuf; Koundal, M.; Mohan, B.; Taneja, N. Prevalence, Virulence Gene Profiling and Characterization of Enteroaggregative Escherichia coli from Acute Diarrhea, Asymptomatic Nourished and Malnourished Children Less Than 5 Years of Age in India. J. Pediatr. 2021, 234, 106–114.e5. [Google Scholar] [CrossRef]
- Tzipori, S.; Montanaro, J.; Robins-Browne, R.M.; Vial, P.; Gibson, R.; Levine, M.M. Studies with enteroaggregative Escherichia coli in the gnotobiotic piglet gastroenteritis model. Infect. Immun. 1992, 60, 5302–5306. [Google Scholar] [CrossRef]
- Vial, P.A.; Robins-Browne, R.; Lior, H.; Prado, V.; Kaper, J.B.; Nataro, J.P.; Maneval, D.; Elsayed, A.-E.-D.; Levine, M.M. Characterization of enteroadherent-aggregative Escherichia coli, a putative agent of diarrheal disease. J. Infect. Dis. 1988, 158, 70–79. [Google Scholar] [CrossRef]
- Rüttler, M.E.; Renna, N.F.; Balbi, L.; García, B.; Guidone, L.; Fernández, R.; Puig, O.; Ortiz, A. Characterization of enteroaggregative Escherichia coli strains isolated from children with acute diarrhea, in Mendoza, Argentina. Rev. Argent Microbiol. 2002, 34, 167–170. [Google Scholar]
- Nezarieh, R.; Shakibaie, M.R.; Nave, H.H.; Norouzi, A.; Salajegheh, G.; Hayatbakhsh, M. Distribution of virulence genes, enterotoxin and biofilm formation among enteroaggregative Escherichia coli (EAEC) strains isolated from stools of children with diarrhea in South East Iran. Arch. Pediatr. Infect. Dis. 2015, 3. [Google Scholar] [CrossRef]
- Sarantuya, J.; Nishi, J.; Wakimoto, N.; Erdene, S.; Nataro, J.P.; Sheikh, J.; Iwashita, M.; Manago, K.; Tokuda, K.; Yoshinaga, M.; et al. Typical Enteroaggregative Escherichia coli Is the Most Prevalent Pathotype among E. coli Strains Causing Diarrhea in Mongolian Children. J. Clin. Microbiol. 2004, 42, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Boisen, N.; Struve, C.; Scheutz, F.; Krogfelt, K.A.; Nataro, J.P. New adhesin of enteroaggregative Escherichia coli related to the Afa/Dr/AAF family. Infect. Immun. 2008, 76, 3281–3292. [Google Scholar] [CrossRef] [PubMed]
- Nataro, J.P.; Yikang, D.; Giron, J.A.; Savarino, S.J.; Kothary, M.H.; Hall, R. Aggregative adherence fimbria I expression in enteroaggregative Escherichia coli requires two unlinked plasmid regions. Infect. Immun. 1993, 61, 1126–1131. [Google Scholar] [CrossRef]
- Elias, W.P.; Czeczulin, J.R.; Henderson, I.R.; Trabulsi, L.R.; Nataro, J.P. Organization of biogenesis genes for aggregative adherence fimbria II defines a virulence gene cluster in enteroaggregative Escherichia coli. J. Bacteriol. 1999, 181, 1779–1785. [Google Scholar] [CrossRef]
- Ruiz-Perez, F.; Wahid, R.; Faherty, C.S.; Kolappaswamy, K.; Rodriguez, L.; Santiago, A.; Murphy, E.; Cross, A.; Sztein, M.B.; Nataro, J.P. Serine protease autotransporters from Shigella flexneri and pathogenic Escherichia coli target a broad range of leukocyte glycoproteins. Proc. Natl. Acad. Sci. USA 2011, 108, 12881–12886. [Google Scholar] [CrossRef]
- Boisen, N.; Ruiz-Perez, F.; Scheutz, F.; Krogfelt, K.A.; Nataro, J.P. Short report: High prevalence of serine protease autotransporter cytotoxins among strains of enteroaggregative Escherichia coli. Am. J. Trop. Med. Hyg. 2009, 80, 294–301. [Google Scholar] [CrossRef]
- Navarro-Garcia, F.; Gutierrez-Jimenez, J.; Garcia-Tovar, C.; Castro, L.A.; Salazar-Gonzalez, H.; Cordova, V. Pic, an autotransporter protein secreted by different pathogens in the Enterobacteriaceae family, is a potent mucus secretagogue. Infect. Immun. 2010, 78, 4101–4109. [Google Scholar] [CrossRef]
- Boisen, N.; Scheutz, F.; Rasko, D.A.; Redman, J.C.; Persson, S.; Simon, J.; Kotloff, K.L.; Levine, M.M.; Sow, S.; Tamboura, B.; et al. Genomic Characterization of Enteroaggregative Escherichia coli From Children in Mali. J. Infect. Dis. 2012, 205, 431–444. [Google Scholar] [CrossRef]
- Bouckenooghe, A.R.; Rodrigues, S.; Steffen, R.; Dupont, H.L.; Jiang, Z.D.; Mathewson, J.J.; Adachi, J.; Verenkar, M.P. Markers of enteric inflammation in enteroaggregative Escherichia coli diarrhea in travelers. Am. J. Trop. Med. Hyg. 2000, 62, 711–713. [Google Scholar] [CrossRef][Green Version]
- Greenberg, D.E.; Jiang, Z.-D.; Steffen, R.; Verenker, M.P.; DuPont, H.L. Markers of inflammation in bacterial diarrhea among travelers, with a focus on enteroaggregative Escherichia coli pathogenicity. J. Infect. Dis. 2002, 185, 944–949. [Google Scholar] [CrossRef]
- Farnaud, S.; Evans, R.W. Lactoferrin—A multifunctional protein with antimicrobial properties. Mol. Immunol. 2003, 40, 395–405. [Google Scholar] [CrossRef]
- Legrand, D.; Elass, E.; Carpentier, M.; Mazurier, J. Lactoferrin: A modulator of immune and inflammatory responses. Cell Mol. Life Sci. 2005, 62, 2549–2559. [Google Scholar] [CrossRef] [PubMed]
- Philipson, C.W.; Bassaganya-Riera, J.; Hontecillas, R. Animal models of enteroaggregative Escherichia coli infection. Gut Microbes 2013, 4, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Mirsepasi-Lauridsen, H.C.; Halkjaer, S.I.; Mortensen, E.M.; Lydolph, M.C.; Nordgaard-Lassen, I.; Krogfelt, K.A.; Petersen, A.M. Extraintestinal pathogenic Escherichia coli are associated with intestinal inflammation in patients with ulcerative colitis. Sci. Rep. 2016, 6, 31152. [Google Scholar] [CrossRef] [PubMed]
- Kotlowski, R.; Bernstein, C.N.; Sepehri, S.; Krause, D.O. High prevalence of Escherichia coli belonging to the B2+D phylogenetic group in inflammatory bowel disease. Gut 2007, 56, 669–675. [Google Scholar] [CrossRef]
- Harrington, S.M.; Dudley, E.G.; Nataro, J.P. Pathogenesis of enteroaggregative Escherichia coli infection. FEMS Microbiol. Lett. 2006, 254, 12–18. [Google Scholar] [CrossRef]
- Jafari, A.; Aslani, M.M.; Bouzari, S. Enteroaggregative Escherichia coli, a heterogenous, underestimated and under-diagnosed, E. coli pathotype in Iran. Gastroenterol. Hepatol. Bed. Bench 2013, 6, 71. [Google Scholar] [CrossRef]
- Myhal, M.L.; Laux, D.C.; Cohen, P.S. Relative colonizing abilities of human fecal and K 12 strains of Escherichia coli in the large intestines of streptomycin-treated mice. Eur. J. Clin. Microbiol. 1982, 1, 186–192. [Google Scholar] [CrossRef]
- Saha, D.R.; Guin, S.; Krishnan, R.; Nag, D.; Koley, H.; Shinoda, S.; Ramamurthy, T. Inflammatory diarrhea due to enteroaggregative Escherichia coli: Evidence from clinical and mice model studies. Gut Pathog. 2013, 5, 36. [Google Scholar] [CrossRef]
- Tarchouna, M.; Ferjani, A.; Ben-Selma, W.; Boukadida, J. Distribution of uropathogenic virulence genes in Escherichia coli isolated from patients with urinary tract infection. Int. J. Infect. Dis. 2013, 17, e450–e453. [Google Scholar] [CrossRef] [PubMed]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia coli phylo-typing method revisited: Improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol. 2000, 66, 4555–4558. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, R.; Mohankumar, R.; Kannappan, A. Exploring the Anti-quorum Sensing and Antibiofilm Efficacy of Phytol against Serratia marcescens Associated Acute Pyelonephritis Infection in Wistar Rats. Front. Cell. Infect. Microbiol. 2017, 7, 498. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.-D.; Greenberg, D.; Nataro, J.P.; Steffen, R.; DuPont, H.L. Rate of occurrence and pathogenic effect of enteroaggregative Escherichia coli virulence factors in international travelers. J. Clin. Microbiol. 2002, 40, 4185–4190. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Jenkins, C. Enteroaggregative Escherichia coli. Curr. Top. Microbiol. Immunol. 2018, 416, 27–50. [Google Scholar] [CrossRef]
- Chattaway, M.A.; Dallman, T.; Okeke, I.N.; Wain, J. Enteroaggregative E. coli O104 from an outbreak of HUS in Germany 2011, could it happen again? J. Infect. Dev. Ctries 2011, 5, 425–436. [Google Scholar] [CrossRef]
- Steiner, T.S.; Lima, A.A.M.; Nataro, J.P.; Guerrant, R.L. Enteroaggregative Escherichia coli Produce Intestinal Inflammation and Growth Impairment and Cause Interleukin-8 Release from Intestinal Epithelial Cells. J. Infect. Dis. 1998, 177, 88–96. [Google Scholar] [CrossRef]
- Nataro, J.P.; Kaper, J.B.; Robins-Browne, R.; Prado, V.; Vial, P.; Levine, M.M. Patterns of adherence of diarrheagenic Escherichia coli to HEp-2 cells. Pediatr. Infect. Dis. J. 1987, 6, 829–831. [Google Scholar] [CrossRef]
- Opintan, J.A.; Nataro, J.P.; Gepi-Attee, R.; Newman, M.J.; Guerrant, R.L.; Roche, J.K.; Affrim, R.; Sevilleja, J.E.A.D.; Ayeh-Kumi, P.F.; Warren, C.A. Pediatricdiarrhea in Southern Ghana: Etiology and association with intestinal inflammation and malnutrition. Am. J. Trop. Med. Hyg. 2010, 83, 936–943. [Google Scholar] [CrossRef]
- Canizalez-Roman, A.; Navarro-García, F. FodrinCaM-binding domain cleavage by Pet from enteroaggregative Escherichia coli leads to actin cytoskeletal disruption. Mol. Microbiol. 2003, 48, 947–958. [Google Scholar] [CrossRef] [PubMed]
- Morin, N.; Tirling, C.; Ivison, S.M.; Kaur, A.P.; Nataro, J.P.; Steiner, T.S. Autoactivation of the AggR regulator of enteroaggregative Escherichia coli in vitro and in vivo. FEMS Immunol. Med. Microbiol. 2010, 58, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Okeke, I.N.; Lamikanra, A.; Czeczulin, J.; Dubovsky, F.; Kaper, J.B.; Nataro, J.P. Heterogeneous Virulence of Enteroaggregative Escherichia coli Strains Isolated from Children in Southwest Nigeria. J. Infect. Dis. 2000, 181, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Micenková, L.; Frankovičová, L.; Jaborníková, I.; Bosák, J.; Dítě, P.; Šmarda, J.; Vrba, M.; Ševčíková, A.; Kmeťová, M.; Šmajs, D. Escherichia coli isolates from patients with inflammatory bowel disease: ExPEC virulence- and colicin-determinants are more frequent compared to healthy controls. Int. J. Med. Microbiol. 2018, 308, 498–504. [Google Scholar] [CrossRef]
- De La Fuente, M.; Franchi, L.; Araya, D.; Díaz-Jiménez, D.; Olivares, M.; Álvarez-Lobos, M.; Golenbock, U.; González, M.-J.; López-Kostner, F.; Quera, R.; et al. Corrigendum to “Escherichia coli isolates from inflammatory bowel diseases patients survive in macrophages and activate NLRP3 inflammasome”. Int. J. Med. Microbiol. 304 (3–4) (2014) 384–392. Int. J. Med. Microbiol. 2015, 305, 434. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Isolates | Non-Clinical Isolates | |||
|---|---|---|---|---|
| EAEC Virulence Gene | Human EAEC n = 30 (%) | Animal EAEC n = 10 (%) | Environmental and Chutney EAEC n = 10 (%) | p Value |
| astA | 29 (96.6) | 5 (50) | 9 (90) | 0.01 * |
| sigA | 2 (6.6) | 2 (20) | 0 | 0.3 |
| Pic | 2 (6.6) | 0 | 1 (10) | 1.0 |
| SepA | 4 (13.3) | 0 | 0 | 0.14 |
| Sat | 6 (20) | 0 | 0 | 0.06 |
| pet | 4 (13.3) | 1 (10) | 0 | 0.6 |
| aap | 14 (46) | 2 (20) | 5 (50) | 0.5 |
| aaiC | 7 (23.3) | 0 | 0 | 0.03 * |
| agg4A | 16 (53.3) | 1 (10) | 5 (50) | 0.14 |
| aggA | 16 (53.3) | 0 | 3 (30) | 0.008 * |
| aafA | 11 (36.6) | 0 | 0 | 0.001 * |
| agg3A | 12 (40) | 0 | 0 | 0.001 * |
| affC | 10 (33.3) | 0 | 0 | 0.003 * |
| ORF61 | 9 (30) | 3 (30) | 3 (30) | 1.0 |
| eilA | 10 | 3 (30) | 0 | 0.19 |
| capU | 8 (26.6) | 1 (10) | 1 (10) | 0.2 |
| air | 5 (16.6) | 0 | 0 | 0.07 |
| espY | 10 (33.3) | 1 (10) | 0 | 0.03 * |
| rmo | 15 (50) | 1 (10) | 1 (10) | 0.005 * |
| shiA | 4 (13.3) | 1 (10) | 1 (10) | 1.0 |
| aggR | 20 (66.6) | 5 (50) | 5 (50) | 0.2 |
| Phylogroups | ||||
| A | 3 (10) | 2 (20) | 0 | 1.0 |
| B1 | 6 (20) | 3 (30) | 2 (20) | 0.7 |
| B2 | 10 (33.3) | 2 (20) | 2 (20) | 0.5 |
| D | 11 (36.6) | 3 (30) | 6 (60) | 0.5 |
| Strain ID | Source of EAEC Isolate | ast A | sig A | pic | sep A | sat | pet | ORF 3 | aap | aai C | agg R | agg4A | aggA | aafA | agg3A | aafC | ORF61 | eilA | capU | air | espY2 | rmoA | shiA | Phylogroup | Virulence Score | Lactoferrin Conc. | Calprotectin Conc. | Colonization upto (Days) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AW-330 | Healthy | + | + | - | - | - | - | - | - | - | + | - | - | + | + | - | - | + | - | - | - | + | - | B1 | 6 | 5942.33 | 3.86 | 5th |
| AW-254 | Healthy | + | - | - | - | - | - | + | + | + | + | + | - | - | - | + | + | + | - | - | + | + | - | B1 | 8 | 3346.62 | 3.47 | 5th |
| M-636 | Diarrhea | + | - | + | + | - | - | + | + | + | + | + | + | - | - | - | + | - | + | - | + | + | - | B1 | 4 | 4375.16 | 3.51 | 10th |
| G-3 | Diarrhea | + | - | - | - | - | - | - | - | - | + | + | - | + | - | - | - | - | - | - | + | - | - | B2 | 6 | 2896.2 | 2.38 | 15th |
| CD-103 | Diarrhea | + | - | - | - | - | - | - | + | - | - | - | + | + | - | + | - | - | - | - | - | + | - | B2 | 10 | 3465.5 | 2.66 | 5th |
| CD-584 | Diarrhea | + | - | - | + | + | - | + | + | - | + | + | - | - | - | + | + | - | + | + | - | - | - | D | 10 | 5249.9 | 3.2 | 15th |
| AW-171 | Healthy | + | - | - | - | - | - | - | - | - | + | + | + | + | + | - | - | - | - | - | - | + | - | B2 | 6 | 4629.3 | 2.01 | 10th |
| 1320/16 | Diarrhea | + | - | - | - | + | + | - | - | - | + | + | + | + | + | - | - | - | - | - | + | - | - | B2 | 7 | 3642.87 | 2.16 | 10th |
| AW-458.2 | Healthy | + | - | - | - | + | - | - | - | - | + | + | + | + | + | - | - | - | - | - | - | + | - | D | 7 | 5058.58 | 2.8 | 5th |
| G-24 | Diarrhea | + | - | - | + | - | - | + | - | - | + | + | - | - | - | - | - | - | - | - | + | + | D | 11 | 5139.19 | 3.18 | 15th | |
| MT-7 | Healthy | + | - | - | - | - | - | - | - | - | + | + | + | + | + | - | - | - | - | - | + | - | B1 | 13 | 5249.02 | 2.6 | 5th | |
| R-524 | Diarrhea | + | - | - | - | - | - | - | + | + | + | + | - | - | - | - | - | + | - | B2 | 5 | 4778.46 | 2.5 | 15th | ||||
| MT-101 | Healthy | + | - | - | - | - | - | + | - | - | - | - | + | - | - | - | + | - | + | - | - | + | - | B2 | 6 | 4814.73 | 2.64 | 5th |
| CD-806 | Diarrhea | + | - | - | - | - | + | - | - | - | - | - | - | + | + | - | - | + | - | + | - | + | - | D | 11 | 5767.95 | 3.32 | 15th |
| R-437 | Diarrhea | + | - | - | - | - | - | + | + | + | + | + | + | - | - | - | + | - | + | - | - | + | - | D | 7 | 5258.0 | 1.52 | 10th |
| MT-38 | Healthy | + | - | - | - | + | - | - | - | - | + | + | + | - | + | - | - | - | - | - | + | - | - | B1 | 9 | 4954.79 | 2.64 | 5th |
| R-431 | Diarrhea | + | - | - | - | - | - | - | + | - | + | + | - | - | - | + | - | + | + | - | + | + | - | D | 7 | 5902.53 | 2.5 | 5th |
| R-708 | Diarrhea | + | - | - | - | - | - | - | - | - | + | + | + | + | + | - | - | - | - | - | - | + | - | B2 | 7 | 2453.70 | 1.8 | 10th |
| M-160 | Diarrhea | + | - | - | - | + | - | - | + | - | + | + | + | - | + | - | - | - | - | - | - | + | - | B2 | 8 | 6202.23 | 3.17 | 5th |
| AW-446 | Healthy | + | - | - | - | - | - | - | - | + | - | - | - | + | + | - | - | - | - | - | - | - | + | A | 6 | 6365.47 | 2.5 | 5th |
| M-510 | Diarrhea | + | - | - | - | - | - | + | - | + | + | - | + | - | - | - | + | + | + | - | - | - | + | A | 4 | 5574.48 | 3.65 | 5th |
| RL1 | Diarrhea | + | - | - | - | - | + | - | + | - | + | + | - | + | + | - | - | - | - | - | - | + | - | B1 | 6 | 4803.38 | 2.8 | 5th |
| R-1483 | Diarrhea | + | + | - | - | - | - | - | - | - | + | - | - | + | + | - | - | + | - | - | - | + | - | D | 10 | 2456.57 | 2.2 | 5th |
| 103.1/16 | Diarrhea | + | - | - | - | - | - | + | + | + | + | + | - | - | - | + | + | + | - | - | + | + | - | D | 7 | 2352.54 | 2.1 | 15th |
| 252/16 | Diarrhea | + | - | + | + | - | - | + | + | + | + | + | + | - | - | - | + | - | + | - | + | + | - | B2 | 8 | 3389.95 | 2.53 | 15th |
| HS-38.3 | Diarrhea | + | - | - | - | - | - | - | - | - | + | + | - | + | - | - | - | - | - | - | + | B2 | 7 | 3947.17 | 3.20 | 5th | ||
| P-11 | Diarrhea | + | - | - | - | - | - | - | + | - | - | - | + | + | - | + | - | - | - | - | - | + | - | A | 8 | 3157.19 | 3.69 | 5th |
| AW 412 | Healthy | + | - | - | + | + | - | + | + | - | + | + | - | - | - | + | + | - | + | + | - | - | D | 5 | 3836.3 | 3.74 | 1st | |
| AW 230 | Healthy | + | - | - | - | - | - | - | - | - | + | + | + | + | + | - | - | - | - | - | - | + | - | B1 | 8 | 1276.96 | 2.69 | 5th |
| 380/15 | Diarrhea | + | - | - | - | + | + | - | - | - | + | + | + | + | + | - | - | - | - | - | + | - | - | D | 8 | 3423.2 | 1.65 | 5th |
| Mean virulence score of human EAEC isolates | 7 | Median Conc. = 4703.34 | Median Conc. = 2.65 | |||||||||||||||||||||||||
| Strain ID | Source | ast A | sig A | pic | sep A | sat | pet | ORF 3 | aap | aai C | agg R | agg4A | aggA | aafA | agg3A | aafC | ORF61 | eilA | capU | air | espY2 | rmoA | shiA | Phylogroup | Virulence Score | Lactoferrin Conc. | Calprotectin Conc. | Colonization upto (Days) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S-37 | Animal | - | + | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | A | 2 | 3443.36 | 1.34 | 1st |
| S-58 | Animal | + | - | - | - | - | - | - | + | - | - | - | - | - | - | - | - | - | - | - | - | - | - | B1 | 2 | 3403.05 | 1.08 | 5th |
| M2 | Animal | - | - | - | - | - | - | - | - | - | + | + | - | - | - | + | - | - | - | - | - | - | D | 3 | 3785.95 | 2.51 | 5th | |
| M 51 | Animal | + | - | - | - | - | - | + | + | - | + | - | - | - | - | + | - | - | - | + | - | - | D | 6 | 4874.18 | 1.66 | 5th | |
| M 68 | Animal | + | - | - | - | - | - | + | - | - | + | + | - | - | - | - | - | - | - | - | - | - | - | B2 | 4 | 3967.32 | 1.94 | 5th |
| S 92 | Animal | - | - | - | - | - | - | - | - | - | + | - | - | - | - | - | + | - | - | - | - | - | - | B2 | 2 | 3342.59 | 1.09 | 5th |
| S 322 | Animal | - | - | - | - | - | - | - | - | - | + | - | - | - | - | - | - | + | - | - | - | - | - | D | 2 | 4420.75 | 2.23 | 5th |
| MI-82 | Animal | + | + | - | - | - | - | - | + | - | - | - | - | - | - | - | - | + | - | - | - | - | + | A | 5 | 4934.64 | 1.8 | 15th |
| MI-73 | Animal | + | - | - | - | - | + | - | - | - | - | - | - | - | - | - | + | + | + | - | - | - | - | B1 | 5 | 2949.62 | 0.66 | 5th |
| MI-39 | Animal | - | - | - | - | - | - | - | - | - | - | + | - | - | - | - | + | - | - | - | - | - | - | B1 | 2 | 4138.6 | 0.52 | 5th |
| Mean virulence score of animal EAEC isolates | 3 | Median Conc. = 3876.63 | Median Conc. = 1.5 | |||||||||||||||||||||||||
| Strain ID | Source | ast A | sig A | pic | sep A | sat | pet | ORF 3 | aap | aai C | agg R | agg4A | aggA | aafA | agg3A | aafC | ORF61 | eilA | capU | air | espY2 | rmoA | shiA | Phylogroup | Virulence Score | Lactoferrin Conc. | Calprotectin Conc. | Colonization upto (Days) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CH-38 | Environment | + | - | - | - | - | - | - | - | - | - | - | + | - | + | - | + | + | - | - | - | - | - | D | 5 | 3755.7 | 0.55 | 5th |
| CH-11 | Environment | + | - | - | - | - | - | - | - | - | + | + | + | - | - | - | - | - | - | - | - | + | - | D | 5 | 2254.36 | 1.32 | 5th |
| CH-72 | Environment | + | - | + | - | - | - | - | - | - | - | + | + | + | - | - | - | - | - | - | - | - | - | B1 | 5 | 1176.20 | 1.36 | 5th |
| CH-23 | Environment | + | - | - | - | - | - | - | - | - | - | + | - | - | - | - | - | - | - | - | - | - | - | B2 | 2 | 3745.64 | 1.30 | 1st |
| CH-85 | Environment | + | - | - | - | - | - | - | - | - | + | + | + | - | - | - | + | + | - | - | - | - | - | D | 6 | 3544.12 | 1.90 | 10th |
| CH-93 | Environment | + | - | - | - | - | - | - | - | + | - | - | - | - | + | - | - | + | - | - | - | - | - | B1 | 4 | 4934.64 | 1.27 | 5th |
| CH-45 | Environment | + | - | - | - | - | - | - | - | + | + | + | - | - | - | + | - | - | - | - | - | - | D | 4 | 4420.75 | 1.47 | 5th | |
| KS-4 | Environment | + | - | - | - | - | - | - | - | - | + | + | + | - | - | - | + | + | - | - | - | - | - | D | 6 | 2415.58 | 0.68 | 5th |
| MD-5 | Environment | - | - | - | - | - | - | + | - | - | - | - | - | - | - | - | - | - | - | - | + | + | - | B2 | 3 | 4269.61 | 1.3 | 1st |
| CH-35 | Environment | + | - | - | - | - | - | - | - | - | + | + | - | - | - | + | + | - | - | - | - | - | - | D | 5 | 1942.00 | 1.34 | 5th |
| Mean virulence score of animal EAEC isolates | 4 | Median Conc. = 3644.88 | Median Conc. = 1.3 | |||||||||||||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Modgil, V.; Narayan, C.; Kaur, H.; Yadav, V.K.; Chaudhary, N.; Kant, V.; Mohan, B.; Bhatia, A.; Taneja, N. Analysis of the Virulence and Inflammatory Markers Elicited by Enteroaggregative Escherichia coli Isolated from Clinical and Non-Clinical Sources in an Experimental Infection Model, India. Microbiol. Res. 2022, 13, 882-897. https://doi.org/10.3390/microbiolres13040062
Modgil V, Narayan C, Kaur H, Yadav VK, Chaudhary N, Kant V, Mohan B, Bhatia A, Taneja N. Analysis of the Virulence and Inflammatory Markers Elicited by Enteroaggregative Escherichia coli Isolated from Clinical and Non-Clinical Sources in an Experimental Infection Model, India. Microbiology Research. 2022; 13(4):882-897. https://doi.org/10.3390/microbiolres13040062
Chicago/Turabian StyleModgil, Vinay, Chandradeo Narayan, Harpreet Kaur, Vivek Kumar Yadav, Naveen Chaudhary, Vishal Kant, Balvinder Mohan, Alka Bhatia, and Neelam Taneja. 2022. "Analysis of the Virulence and Inflammatory Markers Elicited by Enteroaggregative Escherichia coli Isolated from Clinical and Non-Clinical Sources in an Experimental Infection Model, India" Microbiology Research 13, no. 4: 882-897. https://doi.org/10.3390/microbiolres13040062
APA StyleModgil, V., Narayan, C., Kaur, H., Yadav, V. K., Chaudhary, N., Kant, V., Mohan, B., Bhatia, A., & Taneja, N. (2022). Analysis of the Virulence and Inflammatory Markers Elicited by Enteroaggregative Escherichia coli Isolated from Clinical and Non-Clinical Sources in an Experimental Infection Model, India. Microbiology Research, 13(4), 882-897. https://doi.org/10.3390/microbiolres13040062