
Functional Characterisation of Two Novel Deacetylases from Streptococcus pyogenes

 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Bioinformatic Analysis
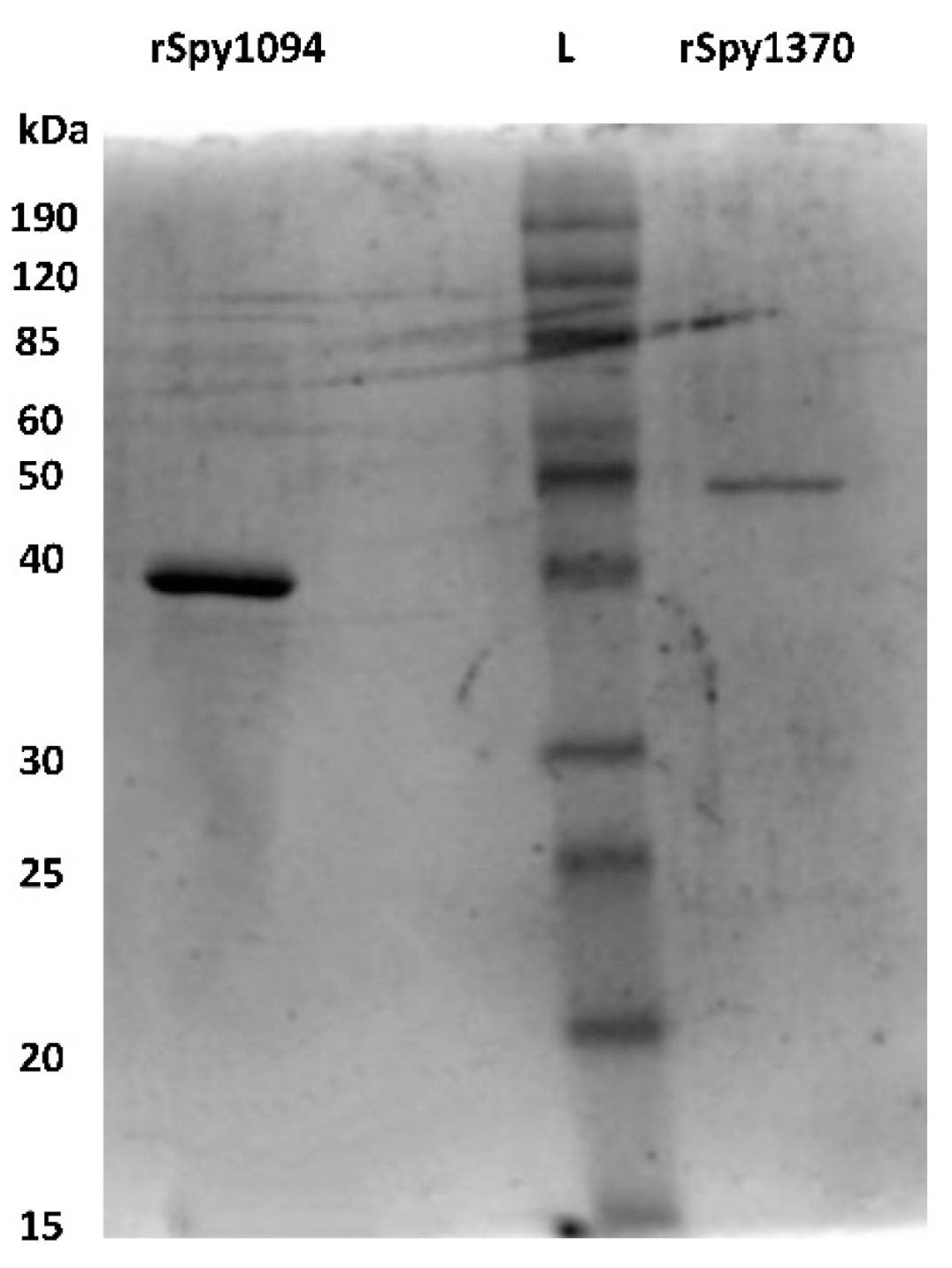
2.2. Generation of Recombinant Spy1370 and Spy1094
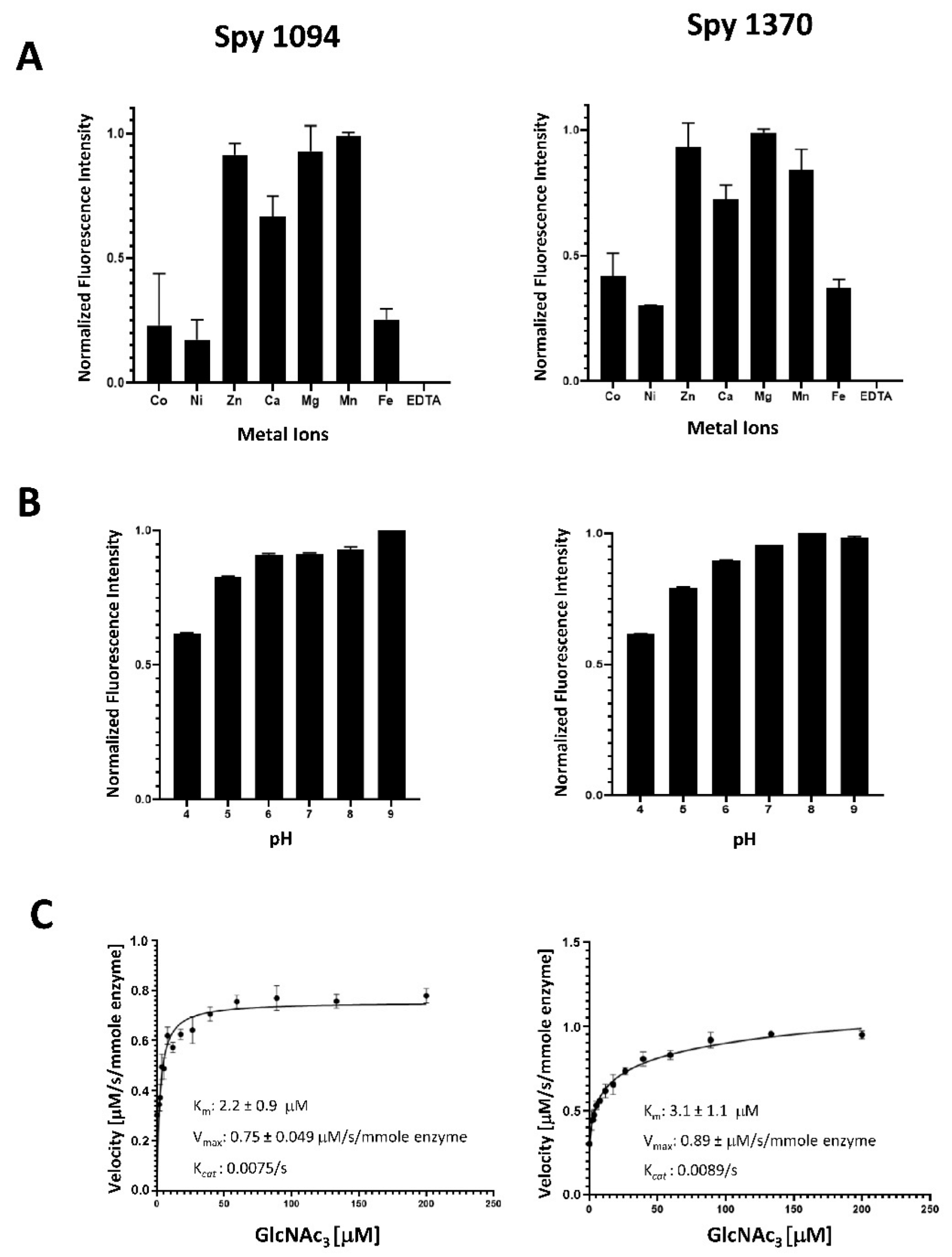
2.3. Biochemical Assays
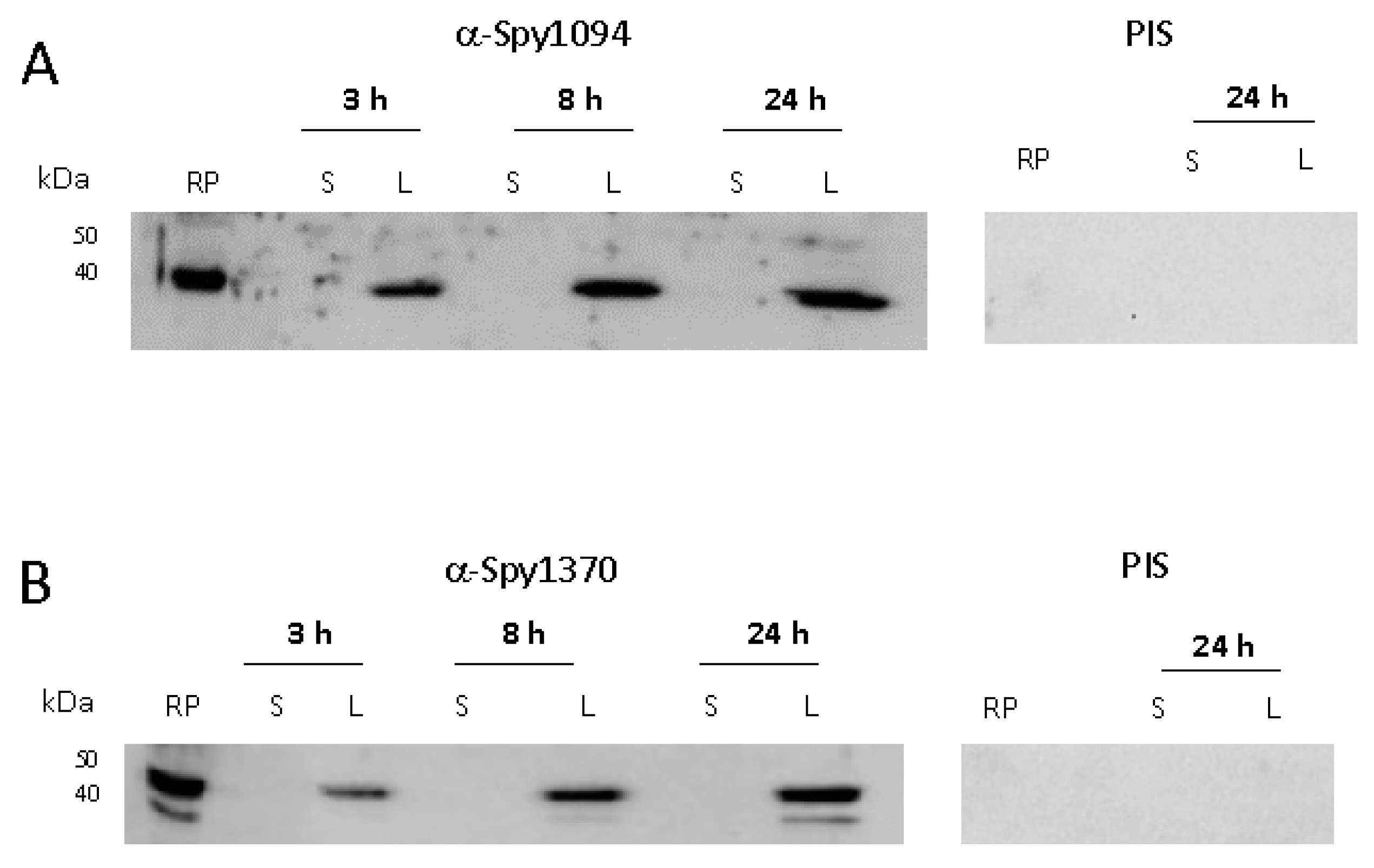
2.4. Generation of Enzyme-Specific Antibodies
2.5. Western-Blot Analysis
3. Results and Discussion
3.1. Identification of Peptidoglycan Deacetylases in S. pyogenes
3.2. Biochemical Analysis of Spy1094 and Spy1370
3.3. Expression of Spy1094 and Spy1370 during S. pyogenes Growth in Culture
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Botteaux, A.; Budnik, I.; Smeesters, P.R. Group A Streptococcus infections in children: From virulence to clinical management. Curr. Opin. Infect. Dis. 2018, 31, 224–230. [Google Scholar] [CrossRef]
- Cunningham, M.W. Pathogenesis of group A streptococcal infections. Clin. Microbiol. Rev. 2000, 13, 470–511. [Google Scholar] [CrossRef]
- Stevens, D.L. Streptococcal toxic-shock syndrome: Spectrum of disease, pathogenesis, and new concepts in treatment. Emerg. Infect. Dis. 1995, 1, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Stevens, D.L.; Bryant, A.E. Severe Group A Streptococcal Infections. In Streptococcus pyogenes: Basic Biology to Clinical Manifestations; Ferretti, J.J., Stevens, D.L., Fischetti, V.A., Eds.; The University of Oklahoma Health Sciences Center: Oklahoma City, OK, USA, 2016. [Google Scholar]
- Cunningham, M.W. Streptococcus and rheumatic fever. Curr. Opin. Rheumatol. 2012, 24, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.J.; Steer, A.C.; Smeesters, P.R.; Keeble, J.; Inouye, M.; Carapetis, J.; Wicks, I.P. Post-infectious group A streptococcal autoimmune syndromes and the heart. Autoimmun. Rev. 2015, 14, 710–725. [Google Scholar] [CrossRef]
- Mosquera, J.; Pedreanez, A. Acute post-streptococcal glomerulonephritis: Analysis of the pathogenesis. Int. Rev. Immunol. 2020, 20, 381–400. [Google Scholar] [CrossRef] [PubMed]
- Carapetis, J.R.; Steer, A.C.; Mulholland, E.K.; Weber, M. The global burden of group A streptococcal diseases. The Lancet. Infect. Dis. 2005, 5, 685–694. [Google Scholar] [CrossRef]
- Brouwer, S.; Barnett, T.C.; Rivera-Hernandez, T.; Rohde, M.; Walker, M.J. Streptococcus pyogenes adhesion and colonization. FEBS Lett. 2016, 590, 3739–3757. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.N.; Barnett, T.C.; Nizet, V.; Walker, M.J. Molecular insight into invasive group A streptococcal disease. Nat. Rev. Microbiol. 2011, 9, 724–736. [Google Scholar] [CrossRef]
- Olsen, R.J.; Shelburne, S.A.; Musser, J.M. Molecular mechanisms underlying group A streptococcal pathogenesis. Cell. Microbiol. 2009, 11, 1–12. [Google Scholar] [CrossRef]
- Proft, T.; Fraser, J. Superantigens: Just like peptides only different. J. Exp. Med. 1998, 187, 819–821. [Google Scholar] [CrossRef] [PubMed]
- Proft, T.; Fraser, J.D. Streptococcal superantigens. Chem. Immunol. Allergy 2007, 93, 1–23. [Google Scholar] [CrossRef]
- Glick, A.D.; Ranhand, J.M.; Cole, R.M. Degradation of group A streptococcal cell walls by egg-white lysozyme and human lysosomal enzymes. Infect. Immun. 1972, 6, 403–413. [Google Scholar] [CrossRef]
- Aragunde, H.; Biarnes, X.; Planas, A. Substrate Recognition and Specificity of Chitin Deacetylases and Related Family 4 Carbohydrate Esterases. Int. J. Mol. Sci. 2018, 19, 412. [Google Scholar] [CrossRef] [PubMed]
- Moynihan, P.J.; Sychantha, D.; Clarke, A.J. Chemical biology of peptidoglycan acetylation and deacetylation. Bioorg. Chem. 2014, 54, 44–50. [Google Scholar] [CrossRef]
- Blair, D.E.; Schuttelkopf, A.W.; MacRae, J.I.; van Aalten, D.M. Structure and metal-dependent mechanism of peptidoglycan deacetylase, a streptococcal virulence factor. Proc. Natl. Acad. Sci. USA 2005, 102, 15429–15434. [Google Scholar] [CrossRef] [PubMed]
- Vollmer, W.; Tomasz, A. Peptidoglycan N-acetylglucosamine deacetylase, a putative virulence factor in Streptococcus pneumoniae. Infect. Immun. 2002, 70, 7176–7178. [Google Scholar] [CrossRef]
- Fittipaldi, N.; Sekizaki, T.; Takamatsu, D.; de la Cruz Dominguez-Punaro, M.; Harel, J.; Bui, N.K.; Vollmer, W.; Gottschalk, M. Significant contribution of the pgdA gene to the virulence of Streptococcus suis. Mol. Microbiol. 2008, 70, 1120–1135. [Google Scholar] [CrossRef]
- Kobayashi, K.; Sudiarta, I.P.; Kodama, T.; Fukushima, T.; Ara, K.; Ozaki, K.; Sekiguchi, J. Identification and characterization of a novel polysaccharide deacetylase C (PdaC) from Bacillus subtilis. J. Biol. Chem. 2012, 287, 9765–9776. [Google Scholar] [CrossRef] [PubMed]
- Boneca, I.G.; Dussurget, O.; Cabanes, D.; Nahori, M.A.; Sousa, S.; Lecuit, M.; Psylinakis, E.; Bouriotis, V.; Hugot, J.P.; Giovannini, M.; et al. A critical role for peptidoglycan N-deacetylation in Listeria evasion from the host innate immune system. Proc. Natl. Acad. Sci. USA 2007, 104, 997–1002. [Google Scholar] [CrossRef] [PubMed]
- Branda, S.S.; Vik, S.; Friedman, L.; Kolter, R. Biofilms: The matrix revisited. Trends Microbiol. 2005, 13, 20–26. [Google Scholar] [CrossRef]
- Milani, C.J.E.; Aziz, R.K.; Locke, J.B.; Dahesh, S.; Nizet, V.; Buchanan, J.T. The novel polysaccharide deacetylase homologue Pdi contributes to virulence of the aquatic pathogen Streptococcus iniae. Microbiology 2010, 156, 543–554. [Google Scholar] [CrossRef]
- Proft, T.; Webb, P.D.; Handley, V.; Fraser, J.D. Two novel superantigens found in both group A and group C Streptococcus. Infect Immun. 2003, 71, 1361–1369. [Google Scholar] [CrossRef]
- Pokrovskaya, V.; Poloczek, J.; Little, D.J.; Griffiths, H.; Howell, P.L.; Nitz, M. Functional characterization of Staphylococcus epidermidis IcaB, a de-N-acetylase important for biofilm formation. Biochemistry 2013, 52, 5463–5471. [Google Scholar] [CrossRef]
- Psylinakis, E.; Boneca, I.G.; Mavromatis, K.; Deli, A.; Hayhurst, E.; Foster, S.J.; Varum, K.M.; Bouriotis, V. Peptidoglycan N-acetylglucosamine deacetylases from Bacillus cereus, highly conserved proteins in Bacillus anthracis. J. Biol. Chem. 2005, 280, 30856–30863. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Zhang, F.; Kang, J.; Zhang, W.; Deng, G.; Xin, Y.; Ma, Y. Mycobacterium tuberculosis Rv1096 protein: Gene cloning, protein expression, and peptidoglycan deacetylase activity. BMC Microbiol. 2014, 14, 174. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aspell, T.; Khemlani, A.H.J.; Loh, J.M.S.; Tsai, C.J.-Y.; Proft, T. Functional Characterisation of Two Novel Deacetylases from Streptococcus pyogenes. Microbiol. Res. 2022, 13, 323-331. https://doi.org/10.3390/microbiolres13020025
Aspell T, Khemlani AHJ, Loh JMS, Tsai CJ-Y, Proft T. Functional Characterisation of Two Novel Deacetylases from Streptococcus pyogenes. Microbiology Research. 2022; 13(2):323-331. https://doi.org/10.3390/microbiolres13020025
Chicago/Turabian StyleAspell, Tiger, Adrina Hema Jethanand Khemlani, Jacelyn Mei San Loh, Catherine Jia-Yun Tsai, and Thomas Proft. 2022. "Functional Characterisation of Two Novel Deacetylases from Streptococcus pyogenes" Microbiology Research 13, no. 2: 323-331. https://doi.org/10.3390/microbiolres13020025
APA StyleAspell, T., Khemlani, A. H. J., Loh, J. M. S., Tsai, C. J.-Y., & Proft, T. (2022). Functional Characterisation of Two Novel Deacetylases from Streptococcus pyogenes. Microbiology Research, 13(2), 323-331. https://doi.org/10.3390/microbiolres13020025