
Identification of Fungi in Flaxseed (L. usitatissimum L.) Using the ITS1 and ITS2 Intergenic Regions

 , and
, and 
Abstract
:1. Introduction
2. Material and Methods
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sidrim, J.J.C.; Rocha, M.F.G. Micologia Médica à luz de Autores Contemporâneos, 1st ed.; Guanabara Koogan: Rio De Janeiro, Brazil, 2004. [Google Scholar]
- Putzke, J.; Putzek, M.T.L. Os Reinos dos Fungos, 2nd ed.; Edunisc: Santa Cruz do Sul, Brazil, 2002. [Google Scholar]
- Midio, A.F.; Martins, D.I. Toxicologia de Alimentos, 1st ed.; Livraria Varela: São Paulo, Brazil, 2000. [Google Scholar]
- FAO (Food and Agriculture Organization). Mycotoxins. Food Safety and Quality. Available online: http://www.fao.org/food/food-safety-quality/a-z-index/mycotoxins/en/ (accessed on 10 April 2021).
- Iamanaka, B.T.; Oliveira, I.S.; Taniwaki, M.H. Micotoxinas em alimentos. Anais Acad. Pernambucana Ciência Agronômica 2010, 7, 138–161. [Google Scholar]
- Manu, N.; Opit, G.P.; Osekre, E.A.; Arthur, F.H.; Mbata, G.; Armstrong, P.; Danso, J.K.; McNeill, S.G.; Campbell, J.F. Moisture content, insect pest infestation and mycotoxin levels of maize in markets in the northern region of Ghana. J. Stored Prod. Res. 2019, 80, 10–20. [Google Scholar] [CrossRef]
- Ismaiel, A.A.; Papenbrock, J. Mycotoxins: Producing Fungi and Mechanisms of Phytotoxicity. Agriculture 2015, 5, 492–537. [Google Scholar] [CrossRef] [Green Version]
- Giordano, B.N.E. Efeito do Ozônio Sobre a Micoflora e Aflatoxinas Durante a Armazenagem de Castanha-do-Brasil com Casca (Bertholettia Excelsa H.B.K.). Master’s Thesis, Universidade Federal de Santa Catarina, Florianópolis, Brazil, 2009. [Google Scholar]
- Bechlin, T.R.; Granella, S.J.; Christ, D.; Coelho, S.R.M.; Viecelli, C.A. Evaluation of grain and oil quality of packaged and ozonized flaxseed. J. Stored Prod. Res. 2019, 83, 311–316. [Google Scholar] [CrossRef]
- Singh, K.K.; Mridula, D.; Barnwal, P.; Rehal, J. Flaxseed: A Potential Souce of Food, Feed and Fiber. Food Sci. Nut. 2011, 51, 210–222. [Google Scholar]
- Novello, D.; Pollonio, M.A.R. Caracterização físico-química e microbiológica da linhaça dourada e marrom (Linum Usitatissimum L.). Rev. Inst. Adolfo Lutz. 2012, 71, 2. [Google Scholar]
- Lorenz, P.; Schleper, C. Metagenome—A challenging source of enzyme discovery. J. Mol. Catal. B Enzym. 2002, 19–20, 13–19. [Google Scholar] [CrossRef]
- Andrade, H.F. Caracterização Molecular de Fungos da Micoteca/UFPE e Screening da Produção de Taxol. Bachelor’s Thesis, Universidade Federal de Pernambuco, Recife, Brazil, 2015. [Google Scholar]
- Nicolaisen, M.; Justesen, A.; Knorr, K.; Wang, J.; Pinnschmidt, H. Fungal communities in wheat grain show significant co-existence patterns among species. Fungal Ecol. 2014, 11, 145–153. [Google Scholar] [CrossRef]
- Schmidt, P.-A.; Bálint, M.; Greshake, B.; Bandow, C.; Römbke, J.; Schmitt, I. Illumina metabarcoding of a soil fungal community. Soil Biol. Biochem. 2013, 65, 128–132. [Google Scholar] [CrossRef]
- Cock, P.J.A.; Antao, T.; Chang, J.T.; Chapman, B.; Cox, C.; Dalke, A.; Friedberg, I.; Hamelryck, T.; Kauff, F.; Wilczynski, B.; et al. Biopython: Freely available Python tools for computational molecular biology and bioinformatics. Bioinformatics 2009, 25, 1422–1423. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.-B.; Lee, M.; Kim, D.-H.; Meijer, M.; Majoor, E.; Vankuyk, P.A.; Samson, R.A. Aspergillus cibarius sp. nov., from traditional meju in Korea. J. Microbiol. 2012, 50, 712–714. [Google Scholar] [CrossRef] [PubMed]
- Bal, J.; Yun, S.-H.; Yeo, S.-H.; Kim, J.-M.; Kim, D.-H. Metagenomic analysis of fungal diversity in Korean traditional wheat-based fermentation starter nuruk. Food Microbiol. 2016, 60, 73–83. [Google Scholar] [CrossRef]
- Mamede, A.C.P.B. Avaliação da Atividade Antibacteriana de Fungos do Filo Ascomycota e Basidiomycota sobre Staphylococcus Aureus e Escherichia Coli. Completion of Course Work; Universidade Federal de Santa Catarina: Florianópolis, Brazil, 2012. [Google Scholar]
- Wang, Y.-Y.; Liu, B.; Zhang, X.-Y.; Zhou, Q.-M.; Zhang, T.; Li, H.; Yu, Y.-F.; Zhang, X.-L.; Hao, X.-Y.; Wang, M.; et al. Genome characteristics reveal the impact of lichenization on lichen-forming fungus Endocarpon pusillum Hedwig (Verrucariales, Ascomycota). BMC Genom. 2014, 15, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagee, D.; Hardan, A.A.; Botero, J.; Arnone, J.T. Genomic clustering whitin functionally related gene families in Ascomycota fungi. Comput. Struct. Biotechnol. J. 2020, 18, 3267–3277. [Google Scholar] [CrossRef] [PubMed]
- Souza, R.F. Análise de Expressão Diferencial em Transcriptomas. Curso de Introdução à Bioinformática Aplicada a Genômica. Available online: http://www.uel.br/laboratorios/lbi/pages/arquivos/curso/01_curso_transcriptoma.pdf (accessed on 7 March 2021).
- Dilthey, A.T.; Jain, C.; Koren, S.; Phillippy, A.M. Strain-level metagenomic assignment and compositional estimation for long reads with MetaMaps. Nat. Commun. 2019, 10, 3066. [Google Scholar] [CrossRef] [Green Version]
- Demain, A.L.; Martens, E. Production of valuable compounds by molds and yeasts. J. Antibiot. 2017, 70, 347–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, N.; Rawat, R.; Sharma, R.; Oberoi, H.S.; Srivastava, M.; Singh, J. Effect of Nickel–Cobaltite Nanoparticles on Production and Thermostability of Cellulases from Newly Isolated Thermotolerant Aspergillus fumigatus NS (Class: Eurotiomycetes). Appl. Biochem. Biotechnol. 2014, 174, 1092–1103. [Google Scholar] [CrossRef]
- Steenwyk, J.L.; Shen, X.X.; Lind, A.L.; Goldman, G.H.; Rokas, A. A robust phylogenomic timetree by biotechnologically and medically important fungi from Aspergillaceae (Eurotiomycetes, Ascomycota). BioRxiv 2018, 370429. [Google Scholar] [CrossRef] [Green Version]
- Eltayeb, M.M.; Eltigani, S.A.; Taniguchi, T. Pyrosequencing scrutiny of bacterial and fungal communities in two Sudanese sorghum-based fermented foods. Ann. Microbiol. 2020, 70, 53. [Google Scholar] [CrossRef]
- Forwood, D.L.; Caro, E.; Holman, D.B.; Meale, S.J.; Chaves, A.V. Ensiling sorghum with unsalable pumpkin improves feed digestibility with minimal influence on the rumen microbial population using the rumen simulation technique. Appl. Microbiol. Biotechnol. 2021, 105, 3289–3300. [Google Scholar] [CrossRef]
- Hubka, V.; Kolarik, M.; Kubatova, A.; Peterson, S.W. Taxonomic revision of Eurotium and transfer of species to Aspergillus. Mycologia 2013, 105, 912–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klich, M.A.; Thomas, S.H.; Mellon, J.E. Field Studies on the Mode of Entry of Aspergillus flavus into Cotton Seeds. Mycologia 1984, 76, 665. [Google Scholar] [CrossRef]
- Pitt, J.I. Fungal ecology and the occurrence of mycotoxins. In Mycotoxins and Phycotoxins: Advances in Determination, Toxicology and Exposure Management; Wageningen Academic Publishers: Wageningen, The Netherlands, 2006; pp. 33–41. [Google Scholar]
- Pitt, J.I.; Hocking, A.D. Fungi and Food Spoilage, 3rd ed.; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Zalar, P.; De Hoog, G.S.; Schroers, H.-J.; Frank, J.M.; Gunde-Cimerman, N. Taxonomy and phylogeny of the xerophilic genus Wallemia (Wallemiomycetes and Wallemiales, cl. et ord. nov.). Antonie Leeuwenhoek 2005, 87, 311–328. [Google Scholar] [CrossRef] [PubMed]
- Zajc, J.; Gunde-Cimerman, N. The Genus Wallemia—From Contamination of Food to Health Threat. Microorganisms 2018, 6, 46. [Google Scholar] [CrossRef] [Green Version]
- Gruzdeviene, E.; Mankeviciene, A.; Lugauskas, A.; Repeckiene, J. The effect of environmental conditions on the variation of fungi and mycotoxin contents in oil flax seed. Ekologija 2006, 3, 64–70. [Google Scholar]
- Bovo, B.; Andrighetto, C.; Carlot, M.; Corich, V.; Lombardi, A.; Giacomini, A. Yeast population dynamics during pilot-scale storage of grape marcs for the production of Grappa, a traditional Italian alcoholic beverage. Int. J. Food Microbiol. 2009, 129, 221–228. [Google Scholar] [CrossRef]
- Yonzan, H.; Tamang, J.P. Microbiology and Nutritional Value ofSelroti, an Ethnic Fermented Cereal Food of the Himalayas. Food Biotechnol. 2010, 24, 227–247. [Google Scholar] [CrossRef]
- Herman, T.; Trigo-Stockli, D. Mycotoxins in Infeed Grain and Ingredients; Kansas State University: Manhattan, KS, USA, 2008. [Google Scholar]
- Dias, I.E. Crescimento Micelar e Produção de Toxinas por Fungos de Armazenamento Associados a Grãos de Milho sob Diferentes Níveis de Restrição Hídrica. Ph.D. Thesis, Universidade Federal de Lavras, Minas Gerais, Brazil, 2012. [Google Scholar]
- D’Aarce, M.A.B.R. Pós-colheita e Armazenamento de grãos. Material Didático. Departamento de Agroindústria, Alimentos e nutrição, ESALQ/USP. Available online: http://sinueloagropecuaria.com.br/wp-content/uploads/2016/09/armazenamento-de-graos-1.pdf (accessed on 18 April 2020).
- Tsai, G.-J.; Yu, S.-C. Detecting Aspergillus parasiticus in cereals by an enzyme-linked immunosorbent assay. Int. J. Food Microbiol. 1999, 50, 181–189. [Google Scholar] [CrossRef]
- Garcia, D.M. Análise de Atividade de Água em Alimentos Armazenados no Interior de Granjas de Integração Avícola. Master’s Thesis, Universidade Federal do Rio Grande, Rio Grande do Sul, Brazil, 2004. [Google Scholar]
- Sibamoto, T.; Bjeldanes, L.F. Introduction to Food Toxicology, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]

{kind=link}
| Kingdom | Phylum | Class | Order | Family | Genus |
|---|---|---|---|---|---|
| Fungi | Ascomycota | Dothideomycetes | Pleosporales | Pleosporaceae | Alternaria |
| Fungi | Ascomycota | Eurotiomycetes | Eurotiales | Aspergillaceae | Aspergillus |
| Fungi | Ascomycota | Eurotiomycetes | Eurotiales | Aspergillaceae | Aspergillus |
| Fungi | Ascomycota | Eurotiomycetes | Eurotiales | Aspergillaceae | Aspergillus |
| Fungi | Ascomycota | Dothideomycetes | Cladosporiales | Cladosporiaceae | Cladosporium |
| Fungi | Ascomycota | Saccharomycetes | Saccharomycetales | Saccharomycodaceae | Hanseniaspora |
| Fungi | Basidiomycota | Wallemiomycetes | Wallemiales | Unidentified | Wallemia |
| Fungi | Ascomycota | Saccharomycetes | Saccharomycetales | Saccharomycetaceae | Zygosaccharomyces |
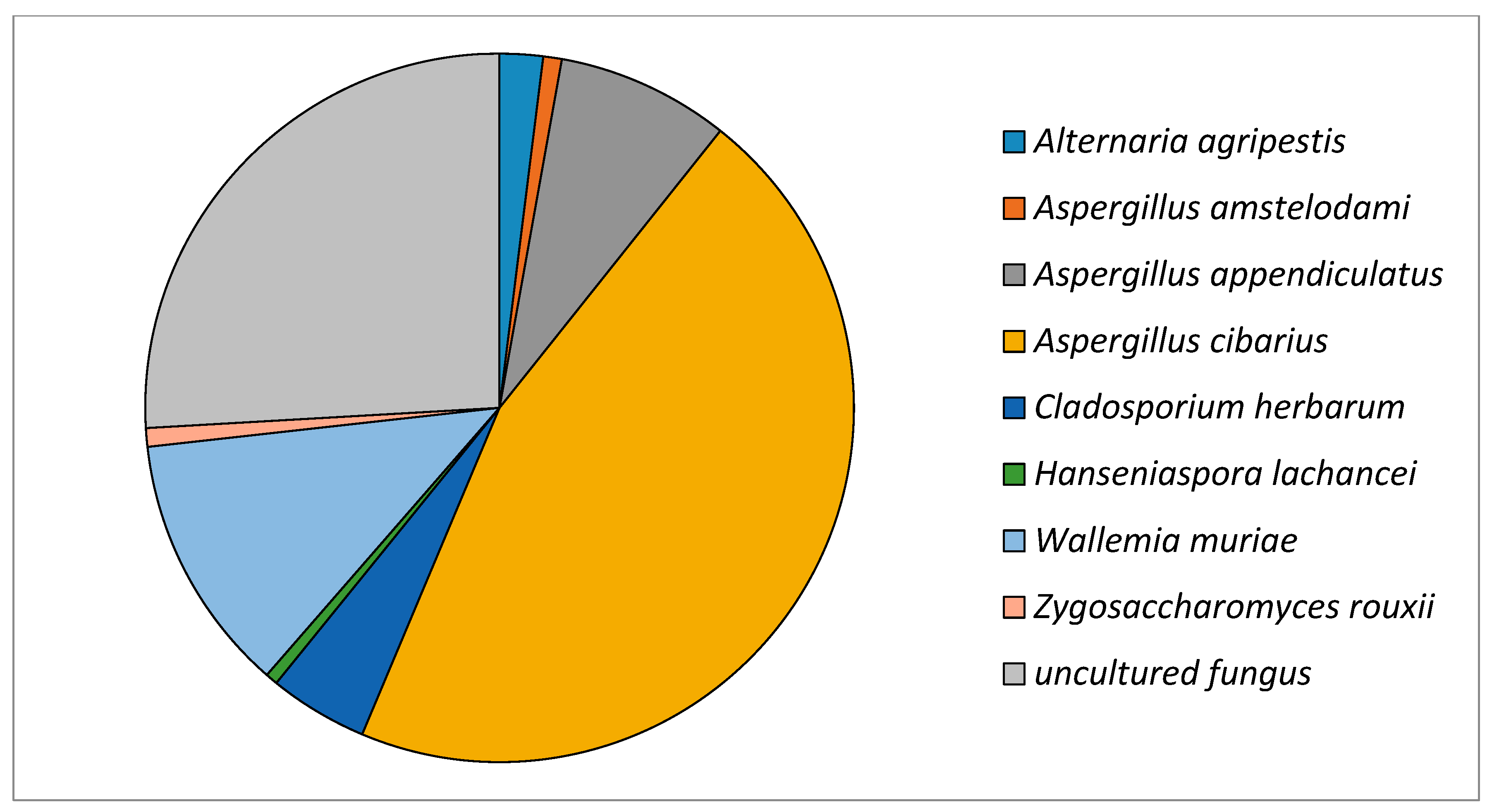
| Identified Species | Number of Reads |
|---|---|
| Alternaria agripestis | 7 |
| Aspergillus amstelodami | 3 |
| Aspergillus appendiculatus | 28 |
| Aspergillus cibarius | 162 |
| Cladosporium herbarum | 16 |
| Hanseniaspora lachancei | 2 |
| Wallemia muriae | 42 |
| Zygosaccharomyces rouxii | 3 |
| Uncultured fungi | 92 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rollemberg, N.d.C.; Hassemer, G.d.S.; Pierezan, M.D.; Maran, B.M.; Dalla Nora, F.M.; Verruck, S. Identification of Fungi in Flaxseed (L. usitatissimum L.) Using the ITS1 and ITS2 Intergenic Regions. Microbiol. Res. 2022, 13, 315-322. https://doi.org/10.3390/microbiolres13020024
Rollemberg NdC, Hassemer GdS, Pierezan MD, Maran BM, Dalla Nora FM, Verruck S. Identification of Fungi in Flaxseed (L. usitatissimum L.) Using the ITS1 and ITS2 Intergenic Regions. Microbiology Research. 2022; 13(2):315-322. https://doi.org/10.3390/microbiolres13020024
Chicago/Turabian StyleRollemberg, Nathalia de Castro, Guilherme de Souza Hassemer, Milena Dutra Pierezan, Bruna Marchesan Maran, Flávia Michelon Dalla Nora, and Silvani Verruck. 2022. "Identification of Fungi in Flaxseed (L. usitatissimum L.) Using the ITS1 and ITS2 Intergenic Regions" Microbiology Research 13, no. 2: 315-322. https://doi.org/10.3390/microbiolres13020024
APA StyleRollemberg, N. d. C., Hassemer, G. d. S., Pierezan, M. D., Maran, B. M., Dalla Nora, F. M., & Verruck, S. (2022). Identification of Fungi in Flaxseed (L. usitatissimum L.) Using the ITS1 and ITS2 Intergenic Regions. Microbiology Research, 13(2), 315-322. https://doi.org/10.3390/microbiolres13020024