Abstract
Background: Development of cirrhosis and hepatocellular carcinoma (HCC) in patients with high alcohol intake is modulated by genetic predispositions. Genetic variation in angiotensin II type 1 receptor (AGTR1) has been described as a risk factor for non-alcoholic fatty liver disease in Asian patients. Methods: We analysed Caucasian patients with alcohol–associated cirrhosis without (n = 238) and with (n = 339) HCC, healthy controls (n = 200), and HCV–infected cirrhotic patients with and without HCC (n = 263) for association with the polymorphisms rs3772622 and rs2276736 in AGTR1. Results: Rs2276736 in AGTR1 was associated with both low–density lipoprotein (LDL) cholesterol levels and hepatic steatosis in patients with alcohol–associated liver disease. The distribution of genotypes for both rs3772622 and rs2276736 in AGTR1 were comparable between controls, cirrhosis patients, and those with HCC. Minor allele frequencies were 32% (44%) in healthy controls, 35%/34% (46%/45%) in alcohol–associated liver disease without/with HCC and 31%/38% (43%/39%) in HCV cirrhosis and HCV HCC, respectively. The genotype of the most important genetic risk factor for fatty liver disease, PNPLA3 I148M, did not interact with the AGTR1 polymorphisms. Conclusion: Genetic variation in AGTR1, although associated with blood lipid levels and hepatic steatosis, is not a risk factor for alcohol–associated cirrhosis or HCC in Caucasians.
1. Introduction
Cirrhosis, the end–stage of chronic liver disease, and hepatocellular carcinoma (HCC), a primary hepatic malignancy arising from cirrhosis, are frequent complications of liver disease worldwide, accounting for approximately 4% of total deaths [1]. In addition to viral hepatitis and metabolic-dysfunction associated steatotic liver disease (MASLD), alcohol–associated liver disease is a very common cause of cirrhosis [2].
However, only a minor subset of patients with heavy alcohol abuse develop cirrhosis [2]. Genetic predisposition is considered a major risk factor for developing liver disease in the presence of alcohol abuse [2]. The most well–known genetic risk factor, among others, is the I148M polymorphism in the patatin-like phospholipase domain-containing protein 3 (PNPLA3)-gene [3]. MASLD, formerly known as non–alcoholic fatty liver disease (NAFLD), and alcohol–associated liver disease display a broad overlap in genetic risk factors [4]. Therefore, loci conferring a predisposition to MASLD are also interesting candidate genes for alcohol–associated liver disease.
In this regard, polymorphisms in the angiotensin II type 1 receptor (AGTR1) gene, most notably the minor G variant of rs3772622, were associated with a decreased risk of NAFLD and less severe fibrosis in NAFLD in a cohort of Japanese patients [5]. In contrast, another study reported more advanced fibrosis in carriers of the G allele of the rs3772622 polymorphism [6]. In the same study, protection from NAFLD by AGTR1 rs2276736 was found in Indian patients [6]. Importantly, the authors of this study also described a significant interaction between rs3772627 and another AGTR1 polymorphism with PNPLA3 rs738409, which corresponds to PNPLA3 I148M. In addition, genetic variation at AGTR1 was associated with high–density lipoprotein levels in a cohort of patients with or at risk for arterial hypertension [7]. Another AGTR1 polymorphism is linked to postprandial lipid levels [8]. Mechanistically, AGTR1 has been implicated in the progression of fibrosis in liver diseases in animal models [9].
Further support for the potential role of AGTR1 polymorphisms in fatty liver disease comes from studies linking genetic variation at the AGTR1 locus to altered lipid profiles in large cohorts [7,8]. In an observational longitudinal study, 314 healthy subjects, in whom obesity, diabetes, and hypertension were carefully excluded, were followed–up for the development of NAFLD and hypertension after they had been genotyped for the AGTR1 rs5186 polymorphism. Genetic variation at AGTR1 rs5186 predicted the development of NAFLD over nearly 10 years with an odds ratio of 1.67. In the same study, 78 non–diabetic patients with NAFLD diagnosed by liver histology were subjected to an oral fat tolerance test. In this subset of patients, genetic variation at AGTR1 rs5186 was not only linked to insulin resistance but also to elevated postprandial very low–density lipoprotein (VLDL) cholesterol and triglyceride levels despite comparable fasting lipid levels. In addition, AGTR1 rs5186 is associated with nuclear factor-κB activation, a central regulator of inflammation, in monocytes [7]. In a second study [8], 213 subjects with normal blood pressure and 242 subjects with pre-hypertension were analysed for AGTR1 polymorphisms and lipid levels. While genetic variation at AGTR1 rs5186 was linked to systolic blood pressure after adjusting for sex and age (p = 0.005), genetic variation at AGTR1 rs276736 was associated with high–density lipoprotein (HDL) cholesterol levels (age and sex–adjusted p value 0.039).
Unraveling genetic factors associated with the occurrence of clinical endpoints in liver disease may not only help to elucidate the relevant molecular pathomechanisms but also stratify patients for surveillance and interventions based on their genetic risk profile.
To clarify whether AGTR1 polymorphisms are linked to the risk of cirrhosis as a terminal stage of fibrosis and to HCC development in alcohol–associated liver disease, we analysed large cohorts of patients for the AGTR1 rs3772622 and rs2276736 polymorphisms.
2. Materials and Methods
2.1. Patients
A total of 577 patients with alcohol–associated cirrhosis, of whom 339 had HCC, were included. Blood specimens for DNA and clinical data were collected at the University Hospital Bonn, Berlin Department of Hepatology and Gastroenterology, and the Division of Hepatology of the Leipzig University Medical Centre. In addition, 263 patients with chronic hepatitis C virus (HCV) infection, of whom 102 had cirrhosis, 161 HCC, respectively, and 200 healthy individuals were used as controls. The healthy controls were blood donors and participants in cancer screening programs without history, clinical, or laboratory signs of liver disease. All participants in this study were Caucasians.
The diagnosis of cirrhosis was made by liver biopsy or based on a typical combination of clinical, laboratory, and radiological findings. Clinical data collected for this study included standard demographic data (such as age, sex, weight, and presence of diabetes) and laboratory data (such as alanine aminotransferase (ALT), aspartate aminotransferase (AST), gamma-glutamyltransferase (GGT), platelet count, bilirubin level, model for end-stage liver disease (MELD) score, serum triglyceride, total cholesterol, low-density lipoprotein (LDL) and high-density lipoprotein (HDL) cholesterol). Where available, the grade of hepatic steatosis, as evaluated by liver biopsy, was recorded. In addition, survival of patients with HCC was assessed.
Patients were classified as having alcohol–associated cirrhosis if their average alcohol intake exceeded 300 g ethanol per week, and other common causes of cirrhosis, such as viral hepatitis or hemochromatosis, were excluded. We chose a cut-off of 300 g ethanol per week to include only patients clearly above the upper limit of alcohol intake deemed acceptable for non-alcoholic fatty liver disease [10].
HCC was diagnosed based on international guidelines [11] and ascertained by histology in cases of uncertainty.
HCV diagnosis relied on the standard high-sensitivity assays used in clinical practice. In detail, patients were deemed to have HCV–induced cirrhosis if hepatitis C RNA was detected in the blood of cirrhotic patients without competing causes of cirrhosis.
2.2. Determination of AGTR1 and PNPLA3 Genotypes
Genomic DNA was extracted from 200 µL of EDTA blood using the QIAamp Blood Mini Kit (Qiagen, Hilden, Germany) following the manufacturer’s instructions. Both AGTR1 and PNPLA3 genotypes were determined using real–time PCR on. a LightCycler® 96 system (Roche, Mannheim, Germany). We used LightSNiP (Simple Probe) assays purchased from TIB-MolBiol (Berlin, Germany) using the rs numbers (rs3772622 and rs2276736) as references. The final volume of samples was 10 µL and contained 1 µL DNA solution, 5 µL Blue Probe qPCR 2× Mix (Biozym Scientific GmbH, Hessisch Oldendorf, Germany), 0.5 µL Primer/Probe Mix, and 3.5 µL PCR grade water. Cycling conditions were set as follows: initial denaturation at 95 °C for 10 min, then 45 cycles of denaturation at 95°C for 10 s, annealing at 60 °C for 10 s, and finally extension at 72 °C for 15 s. At the end of each annealing phase, fluorescence was measured at 60 °C. After PCR was completed, a melting curve of the PCR products was plotted by denaturation at 95 °C for 20 s, holding the sample at 40° C for 20 s, and then gradually heating the sample to 85 °C at a ramp rate of 0.2 °C/s with continuous acquisition of fluorescence. These methods have been previously described in detail [12].
2.3. Statistical Analysis
We used IBM SPSS Statistics software version 28 (IBM, New York, NY, USA) was used for statistical analysis. Allele and genotype frequencies were calculated using SNPStats, a web tool for SNP analysis developed by the Catalan Institute of Oncology. (https://www.snpstats.net/; accessed on several days throughout the year 2023). We tested for significant deviations from the Hardy-Weinberg equilibrium using an exact test. Genotypes were compared with the help of tearson’s goodness–of–fit chi2 test. Forward logistic regression was used to evaluate potential SNP-SNP interactions. Fisher’s exact test was used to compare qualitative data. Student’s t-test and Wilcoxon–Mann–Whitney U test were applied to quantitative data, as appropriate. For survival analysis, we used Kaplan–Meier curves and applied the log–rank test for statistical analysis. A p < 0.05 was the designated threshold for statistical significance.
3. Results
3.1. Study Cohorts
Details of the included patients are presented in Table 1. We analysed clinical data and DNA from 238 patients without and 339 patients with HCC based on of alcohol–associated cirrhosis, 200 healthy controls, and 263 HCV–infected patients, of whom 102 had cirrhosis and 161 had HCC. As expected, the patients in the cirrhotic subgroups were predominantly male and had a mean age well above 50 years. The mean MELD score ranged from 13 to 16 amongst the diseased cohorts, who displayed elevated liver laboratory parameters.

Table 1.
Demographic, clinical, and laboratory details on the study cohorts.
3.2. Association of Genetic Variation at AGTR1 to Lipid Parameters
First, we compared blood and hepatic lipid levels between the different genotypes of AGTR1 rs2276736 and rs3772622 in patients with alcohol–associated cirrhosis to clarify whether the potential impact of the variants on the risk of cirrhosis or HCC might be due to altered lipid levels. While we found that variants of both AGTR1 rs2276736 and rs3772622 were associated with LDL cholesterol levels (Figure 1A,C), homozygous carriers of the AGTR1 rs2276736 minor variant showed increased serum LDL cholesterol levels, and correspondingly decreased intrahepatic lipid levels (Figure 1B,D).
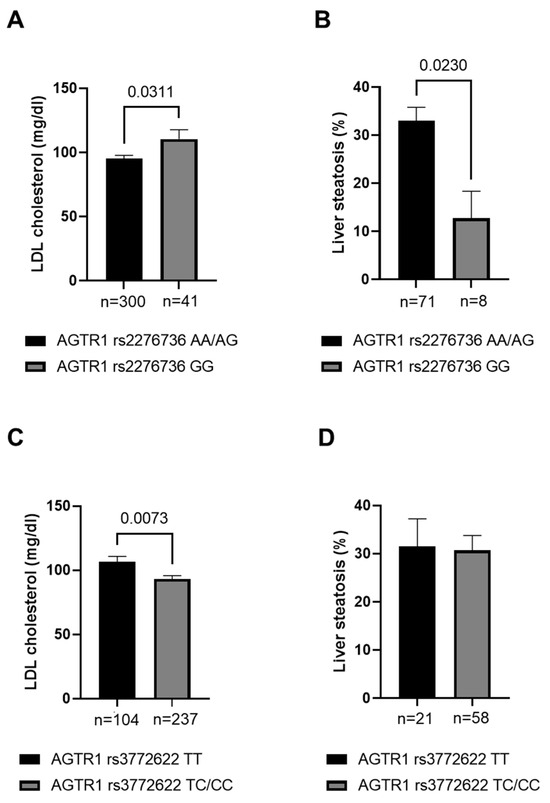
Figure 1.
Serum and hepatic lipid levels in patients with alcohol-associated cirrhosis with different AGTR1 genotypes. Serum LDL cholesterol (A,C) and hepatic lipid levels (B,D) in patients with alcohol-associated cirrhosis carrying different AGTR1 rs2276736 (A,B) and rs3772622 (C,D) genotypes. Statistical analysis with student’s t-test. LDL—Low–density lipoprotein.
3.3. Genotype Distribution Concerning Clinical Endpoints
Detailed information on the genotype distribution of AGTR1 rs3772622 and rs2276736 are shown in Table 2. The minor allele frequency (MAF) for AGTR1 rs2276736 ranged between 31 and 38%, corresponding to 33% reported from the 1000 genomes project [13]; MAF for AGTR1 rs3772622 ranged between 39–46%, corresponding to a frequency of 46% reported from the 1000 genomes project [13]. In all groups, the genotype frequencies of the AGTR1 rs3772622 and rs2276736 polymorphisms were in Hardy–Weinberg equilibrium. We detected no differences in allele and genotype frequencies between patients with and without HCC, between patients with cirrhosis due to HCV–infection and alcohol–associated cirrhosis, or between healthy controls and patients with cirrhosis. The frequencies of carriers of the homozygous minor variant at AGTR1 rs2276736 and of carriers of at least one minor variant at AGTR1 rs3772622 are shown in Figure 2 for the different subgroups, illustrating that significant differences were missing.
Table 2.
Genotype distribution of the AGTR1 rs3772622 and rs2276736 polymorphisms.
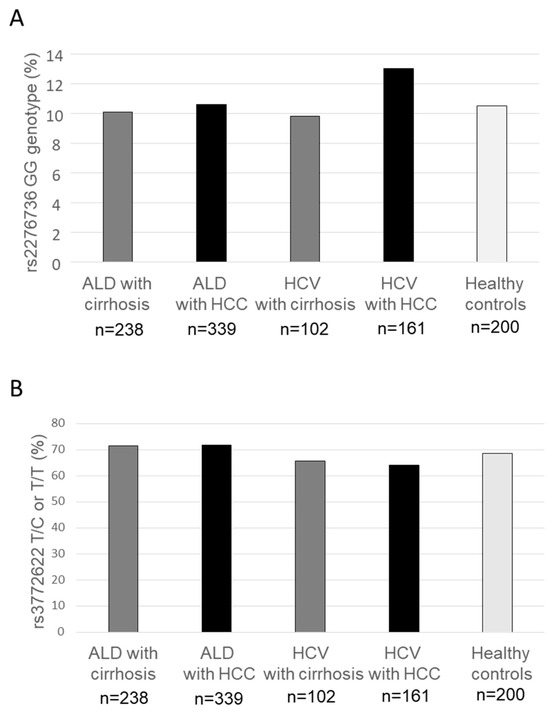
Figure 2.
Frequency of AGTR1 rs2276736 and rs3772622 genotypes in patients with alcohol-associated and HCV–induced cirrhosis and hepatocellular carcinoma. The frequency of homozygous carriers of the AGTR1 rs2276736 minor variant (A) and carriers of the rs3772622 variant (B) were similar among healthy controls and patients with cirrhosis and hepatocellular carcinoma both on the background of alcohol-associated liver disease and hepatitis C virus–induced liver disease. ALD—alcohol-associated liver disease; HCV—hepatitis C virus; HCC—hepatocellular carcinoma.
3.4. Gene-Gene Interaction
Finally, we investigated potential gene–gene interactions between the two AGTR1 polymorphisms and the most important genetic risk variant for alcohol-associated and non-alcoholic fatty liver disease, PNPLA3 I148M [14,15], which might explain why none of the AGTR1 polymorphisms were linked to a clinical endpoint in the overall patient cohort of alcohol-associated cirrhosis and HCC but may indicate that risk modulation for cirrhosis and HCC by the AGTR1 polymorphisms is restricted to a particular genetic background. For NAFLD, relevant gene–gene interactions have been described previously [6]. However, as shown in Table 3 for AGTR1 rs2276736 and in Table 4 for rs3772622, we did not detect any evidence of a relevant gene–gene interaction with PNPLA3 I148M concerning the risk of cirrhosis (Table 3A and Table 4A) or HCC (Table 3B and Table 4B).
Table 3.
(A) SNP-SNP interactions of AGTR1 rs2276736 and PNPLA3 rs738409 concerning the risk of ALD cirrhosis, (B) SNP-SNP interactions of AGTR1 rs2276736 and PNPLA3 rs738409 concerning the risk of alcohol associated HCC.
Table 4.
(A) SNP-SNP interactions of AGTR1 rs3772622 and PNPLA3 rs738409 concerning the risk of ALD cirrhosis, (B) SNP-SNP interactions of AGTR1 rs3772622 and PNPLA3 rs738409 concerning the risk of alcohol associated HCC.
3.5. Survival
Finally, we analysed whether the survival of patients with HCC with a background of alcohol–associated cirrhosis differed according to the AGTR1 rs2276736 and the AGTR1 rs3772622 variants. We found no statistically significant differences in survival based on the genotype at each AGTR1 locus (Figure 3).
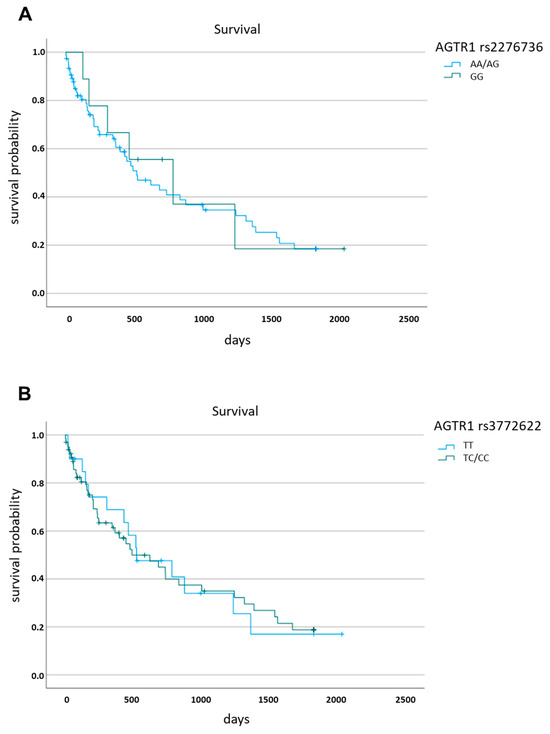
Figure 3.
Survival of patients with hepatocellular carcinoma stratified for the respective AGTR1 genotypes. (A) rs2276736, (B) rs3772622.
4. Discussion
The renin–angiotensin-system (RAS) is a major regulatory mechanism in the human cardiovascular system, modulating fluid balance and blood pressure [16]. Upon reduction in renal blood flow, renin is secreted into the blood by the kidneys after the conversion of a precursor. In the blood, renin transforms angiotensinogen, which is produced by the liver, into angiotensin I. Angiotensin I, in turn, is converted by angiotensin–converting enzyme (ACE) into angiotensin II, which acts both as a strong vasoconstrictor and stimulates the production of aldosterone. Aldosterone increases fluid retention by mediating sodium reabsorption in the renal tubules. The receptor mediating the effect of angiotensin II is angiotensin receptor type I, which is encoded by the gene AGTR1. In addition, an alternative pathway has been discovered [17]: Angiotensin-1-7 is derived from angiotensin I independently from the action of ACE and exerts its effects by the MAS–receptor and angiotensin II–type receptor. This pathway is thought to confer vasodilation, anti-inflammation, and anti-fibrosis effects, as opposed to the classical RAS pathway. Finally, among other pleiotropic functions, angiotensin II has been implicated in the progression of liver fibrosis [18]. In detail, it has been found that activated hepatic stellate cells, the drivers of liver fibrosis, express all components of the RAS system and secrete angiotensin II. When angiotensin II was infused in a rat model of liver disease, fibrosis progressed more rapidly [19]. Furthermore, inflammation markers increased, which may be important because hepatic inflammation drives the development of hepatocellular carcinoma.
Several studies have investigated the role of polymorphisms in AGTR1 to elucidate the impact of angiotensin II on the long-term development of liver disease in humans. Yoneda et al. reported that several genetic variants of AGTR1 were linked to NAFLD in 167 Japanese patients with NAFLD and 435 healthy controls [5]. The AGTR1 rs3772622 polymorphism showed the highest statistical significance with an odd ratio of 1.95. Rs2276736 was reported to be significantly associated with NAFLD among four other single nucleotide polymorphisms in AGTR1. As might be expected, the authors found some degree of linkage disequilibrium between AGTR1 rs3772622 and other AGTR1 polymorphisms associated with fatty liver disease. In addition, a variation in rs3772622 was linked to the stage of fibrosis [5]. Given the role of RAS in the cardiovascular system, Liu et al. analysed whether AGTR1 rs3772622 was linked to cardiovascular disease in Chinese patients with NAFLD. Nearly 600 patients with NAFLD with and without coronary artery disease, more than 300 patients without NAFLD but with coronary artery disease, and more than 300 healthy control patients were genotyped for the AGTR1 rs3772622 polymorphism. While the authors did not detect a significant difference in the distribution of the AGTR1 rs3772622 genotypes between patients with and without NAFLD, they noted a statistically significant association with coronary artery disease, suggesting that the AGTR1 rs3772622 polymorphism might be associated with severe metabolic sequelae compared to NAFLD alone [20]. In a third study [6], several AGTR1 polymorphisms, including rs3772622, were tested for their association with NAFLD in different ethnic subgroups. While the study included a rather small set of 144 patients with NAFLD and 198 controls, the strength of the study is that liver histology was available for all patients with NAFLD. Although the authors failed to replicate an association of the AGTR1 rs3772622 or rs2276736 polymorphism with NAFLD in the overall cohort, they detected a statistically significant association in the subgroup of 31 Indian patients, in contrast to the Chinese and Malay patients. Similarly, the authors did not find a link between AGTR1 rs37772622 or rs2276736 polymorphisms and the severity of NAFLD in the total cohort. However, they also revealed an association between three AGTR1 polymorphisms and non-alcoholic steatohepatitis, including AGTR1 rs2276736. Finally, they performed a gene–gene interaction analysis with PNPLA3 I148M and detected a statistically highly significant interaction. This was also true for a subgroup of Indian patients; however, when patients were regarded separately according to their PNPLA3 I148M genotype, the effect could not be replicated. Concerning histologically proven fibrosis, AGTR1 rs3772622 genotypes again showed a statistically significant association with fibrosis, even after adjusting for the grade of steatosis, ballooning, and lobular inflammation. Again, the authors reported a significant degree of linkage disequilibrium between the different AGTR1 polymorphisms analysed [6].
Drugs such as telmisartan, which blocks angiotensin II receptor 1 and activates peroxisome proliferator-activated receptor (PPAR)γ, a master regulator of hepatic lipid metabolism [21,22], might unite two approaches to tackle steaotic liver disease [23]. In this study, the authors administered telmisartan to mouse models of both diabetes and non–alcoholic steatohepatitis over a period of 6–week and meticulously analysed the liver transcriptome. They observed that non-alcoholic steatohepatitis (NASH) improved along with improvements in the expression of inflammation–and fibrosis-related genes. The interplay between hypertension, RAS activation, and atherosclerosis progression involving lipid metabolism has been discussed for a long time [24]. Nevertheless, the exact mechanistic interplay between AGTR1 and lipid metabolism has still to be elucidated. In addition, although the effect of genetic predisposition to NAFLD was confirmed in a recent genome–wide association meta–analysis by identifying 17 genetic risk loci, AGTR1 was not among these loci [25]. However, the results of this study underline the clinical importance of genetic predisposition: a high genetic risk translated into a 2.5–6-fold elevated risk for NAFLD and, more importantly, cirrhosis and HCC.
Similar to NAFLD, the risk of alcohol–associated cirrhosis and HCC is strongly influenced by genetic predispositions [15]. Genetic predisposition explains why only a subset of patients with severe alcohol abuse develop cirrhosis. NAFLD and alcohol–associated liver disease (ALD) share prominent genetic risk loci. The most well–known is I148M at PNPLA3 [26,27]. The function of PNPLA3 and its pathological consequences have only been partially unravelled so far. In any case, PNPLA3 is involved in the lipid metabolism in the liver cells. Under conditions of high–fat intake, the presence of the minor PNPLA3 148M variant leads to the accumulation of intrahepatic fat. In addition, intrahepatic fat storage in patients carrying the PNPLA3 148M variant prompts an inflammatory hepatic reaction. Another shared genetic risk factor for both NAFLD and ALD is located at transmembrane 6 superfamily 2 (TM6SF2) [28,29]. In contrast to PNPLA3, which seems to be involved in lipid storage, TM6SF2 is implicated in hepatic fat metabolism by mediating the assembly of lipoproteins and VLDL export from the liver. Impaired function in patients with genetic variations leads to increased hepatic fat storage. Similar to PNPLA3, 17β-Hydroxysteroid dehydrogenase type 13 (HSD17B13) [30,31] is located in lipid droplets. Minor variants in HSD17B13 are protective; however, the exact molecular mechanism has not yet been elucidated. Finally, variation in membrane-bound O-acyltransferase domain-containing 7 (MBOAT7) [27,32] is another genetic risk factor for NAFLD and ALD. Its biochemical role is recognized as lysophosphatidylinositol acyltransferase. As such, it may regulate the production of inflammatory lipid mediators, with the MBOAT7 rs641738 minor variant leading to decreased MBOT7 function, increased hepatic lipid storage, and a pro-inflammatory response.
Targeted analysis of AGTR1 polymorphisms in the context of alcohol–associated liver cirrhosis has not been previously reported. Therefore, we analysed a well–characterized and sizable cohort of patients with alcohol–associated cirrhosis to determine the association between the two most prominent AGTR1 polymorphisms, cirrhosis, and the occurrence of HCC. We did not find any connection between the two polymorphisms and the most severe sequelae of ALD, although genetic variation at both loci was associated with peripheral LDL cholesterol levels, and homozygous carriers of the minor AGTR1 rs2276736 variant displayed decreased intrahepatic lipid levels. In addition, we did not find evidence of a gene–gene interaction between PNPLA3 I148M and the investigated AGTR1 polymorphisms, as reported previously for NAFLD patients [6]. Therefore, the lack of association between genetic variation in AGTR1 and ALD complications cannot be attributed to a concealing effect of the variation in PNPLA3.
If we assume that our cohort was sufficiently large to detect any clinically significant difference, two conclusions may be drawn. First, that alcoholic and non–alcoholic liver disease differ concerning the pathophysiological impact of AGTR1. The hypothesis that alcoholic and non–alcoholic fatty liver disease may differ in some aspects regarding genetic predisposition despite all similarities, is supported by the results from a Japanese cohort, which found differences between the two disease groups when analysing 10 different polymorphisms [33]. The second potential conclusion is that ethnic and environmental differences lead to diverging genetic risk factors for fatty liver disease in different regions of the world, as our study comprised only Caucasian patients, while studies reporting AGTR1 polymorphisms as a risk factor for NAFLD included mainly Asian patients [5,6,20]. It is known that ethnic background affects survival in HCC patients, which may not only be linked to genetic predisposition but also to the etiology of HCC, which differs across regions worldwide, and to access to surveillance and treatment programs [34,35,36]. Finally, our results indicate that changes in peripheral and intrahepatic lipid levels may contribute to, but are not sufficient for, the development of overt alcohol–associated liver disease. Thus, the risk of cardiovascular disease, not liver disease, remains the dominant prognostic factor for patients with dyslipidemia [37].
Our study results are limited by the limited number of patients included in our study, which, however, was still large and offers the strength of clinically well–characterised cohorts. In addition, our study had an observational design rather than a prospective design. The latter would be highly desirable but is nearly unfeasible in traits developing over decades, such as cirrhosis and HCC. Third, one may speculate that a substantial number of patients carrying an AGTR1 variant may develop HCC in the near future; therefore we may have missed the effect of genetic variation. However, since we could replicate all major known genetic risk factors for alcohol-associated HCC in our cohort, it is unlikely that the observation time was too short. Finally, the impact on genetic traits may become evident when additional genetic parameters and environmental factors, such as diet, coincide. Therefore, we cannot exclude that under different conditions from the ones investigated here the results may be different.
5. Conclusions
In conclusion, our data do not support a role for the two investigated AGTR1 polymorphisms in the development of cirrhosis or HCC in alcoholic liver disease in Caucasian patients, in contrast to non–alcoholic fatty liver disease in Asian patients, although we found that lipid levels in patients with alcohol-associated cirrhosis were linked to genetic variation in AGTR1. The impact of genetic variation at AGTR1 on the development of complications of ALD in Caucasian patients seems to be low or absent; both polymorphisms may not be suitable for inclusion into models for risk prediction of HCC in alcohol-associated cirrhosis [38].
Author Contributions
Conceptualization, H.D.N. and P.L.; Formal analysis, H.D.N.; Funding acquisition, H.D.N. and J.N.; Investigation, F.S., C.M., B.K. and B.L.; Project administration, P.L.; Resources, J.F., M.M.-S., J.N., T.B., C.P.S. and P.L.; Supervision, J.N. and C.P.S.; Visualization, H.D.N.; Writing—original draft, H.D.N. and P.L.; Writing—review and editing, F.S., J.F., C.M., M.M.-S., B.K., B.L. and T.B. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by Deutsche Krebshilfe (70114349) for HDN. J.N. received funding from the German Research Foundation (DFG SFB/TRR 57 and SPP1937) and the Hector foundation (M88).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki approved by the local ethics committees (Bonn: number 536/20; Leipzig: number 357/19-ek).
Informed Consent Statement
Written informed consent was obtained from all participants involved in the study.
Data Availability Statement
The data presented in this study are available upon request from the corresponding author. The data are not publicly available because of privacy concerns.
Acknowledgments
The authors would like to thank all the study participants, clinicians, and administrative staff who contributed to this study.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Devarbhavi, H.; Asrani, S.K.; Arab, J.P.; Nartey, Y.A.; Pose, E.; Kamath, P.S. Global Burden of Liver Disease: 2023 Update. J. Hepatol. 2023, 79, 516–537. [Google Scholar] [CrossRef] [PubMed]
- Seitz, H.K.; Bataller, R.; Cortez-Pinto, H.; Gao, B.; Gual, A.; Lackner, C.; Mathurin, P.; Mueller, S.; Szabo, G.; Tsukamoto, H. Alcoholic Liver Disease. Nat. Rev. Dis. Primers 2018, 4, 16. [Google Scholar] [CrossRef] [PubMed]
- Vishnubhotla, R.; Kulkarni, A.V.; Sharma, M.; Rao, P.N.; Reddy, D.N. An Update on the Genetics of Alcoholic Liver Disease. Front. Gastroenterol. 2022, 1, 1030399. [Google Scholar] [CrossRef]
- Eslam, M.; George, J. Genetic Contributions to NAFLD: Leveraging Shared Genetics to Uncover Systems Biology. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Yoneda, M.; Hotta, K.; Nozaki, Y.; Endo, H.; Uchiyama, T.; Mawatari, H.; Iida, H.; Kato, S.; Fujita, K.; Takahashi, H.; et al. Association between Angiotensin II Type 1 Receptor Polymorphisms and the Occurrence of Nonalcoholic Fatty Liver Disease. Liver Int. 2009, 29, 1078–1085. [Google Scholar] [CrossRef] [PubMed]
- Zain, S.M.; Mohamed, Z.; Mahadeva, S.; Rampal, S.; Basu, R.C.; Cheah, P.-L.; Salim, A.; Mohamed, R. Susceptibility and Gene Interaction Study of the Angiotensin II Type 1 Receptor (AGTR1) Gene Polymorphisms with Non-Alcoholic Fatty Liver Disease in a Multi-Ethnic Population. PLoS ONE 2013, 8, e58538. [Google Scholar] [CrossRef]
- Fung, M.M.; Rao, F.; Poddar, S.; Mahata, M.; Khandrika, S.; Mahata, S.K.; O’Connor, D.T. Early Inflammatory and Metabolic Changes in Association with AGTR1 Polymorphisms in Prehypertensive Subjects. Am. J. Hypertens. 2011, 24, 225–233. [Google Scholar] [CrossRef]
- Musso, G.; Saba, F.; Cassader, M.; Paschetta, E.; De Michieli, F.; Pinach, S.; Framarin, L.; Berrutti, M.; Leone, N.; Parente, R.; et al. Angiotensin II Type 1 Receptor Rs5186 Gene Variant Predicts Incident NAFLD and Associated Hypertension: Role of Dietary Fat-Induced Pro-Inflammatory Cell Activation. Am. J. Gastroenterol. 2019, 114, 607–619. [Google Scholar] [CrossRef]
- Rajapaksha, I.G.; Gunarathne, L.S.; Angus, P.W.; Herath, C.B. Update on New Aspects of the Renin-Angiotensin System in Hepatic Fibrosis and Portal Hypertension: Implications for Novel Therapeutic Options. J. Clin. Med. 2021, 10, 702. [Google Scholar] [CrossRef]
- Rinella, M.E.; Neuschwander-Tetri, B.A.; Siddiqui, M.S.; Abdelmalek, M.F.; Caldwell, S.; Barb, D.; Kleiner, D.E.; Loomba, R. AASLD Practice Guidance on the Clinical Assessment and Management of Nonalcoholic Fatty Liver Disease. Hepatology 2023, 77, 1797–1835. [Google Scholar] [CrossRef]
- Galle, P.R.; Forner, A.; Llovet, J.M.; Mazzaferro, V.; Piscaglia, F.; Raoul, J.-L.; Schirmacher, P.; Vilgrain, V. EASL Clinical Practice Guidelines: Management of Hepatocellular Carcinoma. J. Hepatol. 2018, 69, 182–236. [Google Scholar] [CrossRef] [PubMed]
- Nischalke, H.D.; Schmalz, F.; Buch, S.; Fischer, J.; Möller, C.; Matz-Soja, M.; Krämer, B.; Langhans, B.; Klüners, A.; Soyka, M.; et al. Genetic Variation of SAMM50 Is Not an Independent Risk Factor for Alcoholic Hepatocellular Carcinoma in Caucasian Patients. Int. J. Mol. Sci. 2022, 23, 15353. [Google Scholar] [CrossRef] [PubMed]
- 1000 Genomes Project Consortium; Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; et al. A Global Reference for Human Genetic Variation. Nature 2015, 526, 68–74. [Google Scholar] [CrossRef]
- Sookoian, S.; Pirola, C.J. Genetics in Non-Alcoholic Fatty Liver Disease: The Role of Risk Alleles through the Lens of Immune Response. Clin. Mol. Hepatol. 2023, 29, S184–S195. [Google Scholar] [CrossRef] [PubMed]
- Stickel, F.; Moreno, C.; Hampe, J.; Morgan, M.Y. The Genetics of Alcohol Dependence and Alcohol-Related Liver Disease. J. Hepatol. 2017, 66, 195–211. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.A.S.; Oudit, G.Y.; Verano-Braga, T.; Canta, G.; Steckelings, U.M.; Bader, M. The Renin-Angiotensin System: Going beyond the Classical Paradigms. Am. J. Physiol.-Heart Circ. Physiol. 2019, 316, H958–H970. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.A.S.; Sampaio, W.O.; Alzamora, A.C.; Motta-Santos, D.; Alenina, N.; Bader, M.; Campagnole-Santos, M.J. The ACE2/Angiotensin-(1–7)/MAS Axis of the Renin-Angiotensin System: Focus on Angiotensin-(1–7). Physiol. Rev. 2018, 98, 505–553. [Google Scholar] [CrossRef] [PubMed]
- Shim, K.Y.; Eom, Y.W.; Kim, M.Y.; Kang, S.H.; Baik, S.K. Role of the Renin-Angiotensin System in Hepatic Fibrosis and Portal Hypertension. Korean J. Intern. Med. 2018, 33, 453–461. [Google Scholar] [CrossRef]
- Bataller, R.; Gäbele, E.; Parsons, C.J.; Morris, T.; Yang, L.; Schoonhoven, R.; Brenner, D.A.; Rippe, R.A. Systemic Infusion of Angiotensin II Exacerbates Liver Fibrosis in Bile Duct-Ligated Rats. Hepatology 2005, 41, 1046–1055. [Google Scholar] [CrossRef]
- Liu, Y.; Lu, L.-L.; Yuan, D.-X.; Geng, N.; Xuan, S.-Y.; Xin, Y.-N. AGTR1 Rs3772622 Gene Polymorphism Increase the Risk of Nonalcoholic Fatty Liver Disease Patients Suffer Coronary Artery Disease in Northern Chinese Han Population. Lipids Health Dis. 2016, 15, 113. [Google Scholar] [CrossRef][Green Version]
- Wang, Y.; Nakajima, T.; Gonzalez, F.J.; Tanaka, N. PPARs as Metabolic Regulators in the Liver: Lessons from Liver-Specific PPAR-Null Mice. Int. J. Mol. Sci. 2020, 21, 2061. [Google Scholar] [CrossRef]
- Francque, S.; Szabo, G.; Abdelmalek, M.F.; Byrne, C.D.; Cusi, K.; Dufour, J.-F.; Roden, M.; Sacks, F.; Tacke, F. Nonalcoholic Steatohepatitis: The Role of Peroxisome Proliferator-Activated Receptors. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 24–39. [Google Scholar] [CrossRef]
- Park, J.G.; Mok, J.S.; Han, Y.I.; Park, T.S.; Kang, K.W.; Choi, C.S.; Park, H.D.; Park, J. Connectivity Mapping of Angiotensin-PPAR Interactions Involved in the Amelioration of Non-Alcoholic Steatohepatitis by Telmisartan. Sci. Rep. 2019, 9, 4003. [Google Scholar] [CrossRef]
- Ferrario, C.M.; Smith, R.; Levy, P.; Strawn, W. The Hypertension-Lipid Connection: Insights into the Relation between Angiotensin II and Cholesterol in Atherogenesis. Am. J. Med. Sci. 2002, 323, 17–24. [Google Scholar] [CrossRef]
- Chen, Y.; Du, X.; Kuppa, A.; Feitosa, M.F.; Bielak, L.F.; O’Connell, J.R.; Musani, S.K.; Guo, X.; Kahali, B.; Chen, V.L.; et al. Genome-Wide Association Meta-Analysis Identifies 17 Loci Associated with Nonalcoholic Fatty Liver Disease. Nat. Genet. 2023, 55, 1640–1650. [Google Scholar] [CrossRef]
- Nischalke, H.D.; Berger, C.; Luda, C.; Berg, T.; Müller, T.; Grünhage, F.; Lammert, F.; Coenen, M.; Krämer, B.; Körner, C.; et al. The PNPLA3 Rs738409 148M/M Genotype Is a Risk Factor for Liver Cancer in Alcoholic Cirrhosis but Shows No or Weak Association in Hepatitis C Cirrhosis. PLoS ONE 2011, 6, e27087. [Google Scholar] [CrossRef]
- Buch, S.; Stickel, F.; Trépo, E.; Way, M.; Herrmann, A.; Nischalke, H.D.; Brosch, M.; Rosendahl, J.; Berg, T.; Ridinger, M.; et al. A Genome-Wide Association Study Confirms PNPLA3 and Identifies TM6SF2 and MBOAT7 as Risk Loci for Alcohol-Related Cirrhosis. Nat. Genet. 2015, 47, 1443–1448. [Google Scholar] [CrossRef]
- Stickel, F.; Buch, S.; Nischalke, H.D.; Weiss, K.H.; Gotthardt, D.; Fischer, J.; Rosendahl, J.; Marot, A.; Elamly, M.; Casper, M.; et al. Genetic Variants in PNPLA3 and TM6SF2 Predispose to the Development of Hepatocellular Carcinoma in Individuals with Alcohol-Related Cirrhosis. Am. J. Gastroenterol. 2018, 113, 1475–1483. [Google Scholar] [CrossRef]
- Longo, M.; Meroni, M.; Paolini, E.; Erconi, V.; Carli, F.; Fortunato, F.; Ronchi, D.; Piciotti, R.; Sabatini, S.; Macchi, C.; et al. TM6SF2/PNPLA3/MBOAT7 Loss-of-Function Genetic Variants Impact on NAFLD Development and Progression Both in Patients and in In Vitro Models. Cell. Mol. Gastroenterol. Hepatol. 2022, 13, 759–788. [Google Scholar] [CrossRef]
- Stickel, F.; Lutz, P.; Buch, S.; Nischalke, H.D.; Silva, I.; Rausch, V.; Fischer, J.; Weiss, K.H.; Gotthardt, D.; Rosendahl, J.; et al. Genetic Variation in HSD17B13 Reduces the Risk of Developing Cirrhosis and Hepatocellular Carcinoma in Alcohol Misusers. Hepatology 2020, 72, 88–102. [Google Scholar] [CrossRef]
- Abul-Husn, N.S.; Cheng, X.; Li, A.H.; Xin, Y.; Schurmann, C.; Stevis, P.; Liu, Y.; Kozlitina, J.; Stender, S.; Wood, G.C.; et al. A Protein-Truncating HSD17B13 Variant and Protection from Chronic Liver Disease. N. Engl. J. Med. 2018, 378, 1096–1106. [Google Scholar] [CrossRef] [PubMed]
- Mancina, R.M.; Dongiovanni, P.; Petta, S.; Pingitore, P.; Meroni, M.; Rametta, R.; Borén, J.; Montalcini, T.; Pujia, A.; Wiklund, O.; et al. The MBOAT7-TMC4 Variant Rs641738 Increases Risk of Nonalcoholic Fatty Liver Disease in Individuals of European Descent. Gastroenterology 2016, 150, 1219–1230.e6. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Kogiso, T.; Taniai, M.; Hashimoto, E.; Tokushige, K. Differences in the Genetic Backgrounds of Patients with Alcoholic Liver Disease and Non-alcoholic Fatty Liver Disease. JGH Open 2019, 3, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Barzi, A.; Zhou, K.; Wang, S.; Dodge, J.L.; El-Khoueiry, A.; Setiawan, V.W. Etiology and Outcomes of Hepatocellular Carcinoma in an Ethnically Diverse Population: The Multiethnic Cohort. Cancers 2021, 13, 3476. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, C.S.; Ruhl, J.; Flynn, G.; Graubard, B.I.; McGlynn, K.A. Trends in Hepatocellular Carcinoma Stage by Racial-Ethnic Group in the United States, 1992–2019. JHEP Rep. 2023, 5, 100868. [Google Scholar] [CrossRef]
- Thylur, R.P.; Roy, S.K.; Shrivastava, A.; LaVeist, T.A.; Shankar, S.; Srivastava, R.K. Assessment of Risk Factors, and Racial and Ethnic Differences in Hepatocellular Carcinoma. JGH Open 2020, 4, 351–359. [Google Scholar] [CrossRef]
- Berberich, A.J.; Hegele, R.A. A Modern Approach to Dyslipidemia. Endocr. Rev. 2022, 43, 611–653. [Google Scholar] [CrossRef]
- Bianco, C.; Jamialahmadi, O.; Pelusi, S.; Baselli, G.; Dongiovanni, P.; Zanoni, I.; Santoro, L.; Maier, S.; Liguori, A.; Meroni, M.; et al. Non-Invasive Stratification of Hepatocellular Carcinoma Risk in Non-Alcoholic Fatty Liver Using Polygenic Risk Scores. J. Hepatol. 2021, 74, 775–782. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).