
The Second Exteroceptive Suppression Period of the Temporalis Muscle Is Altered in Migraine Patients with Allodynia

 ,
,  ,
, 
Abstract
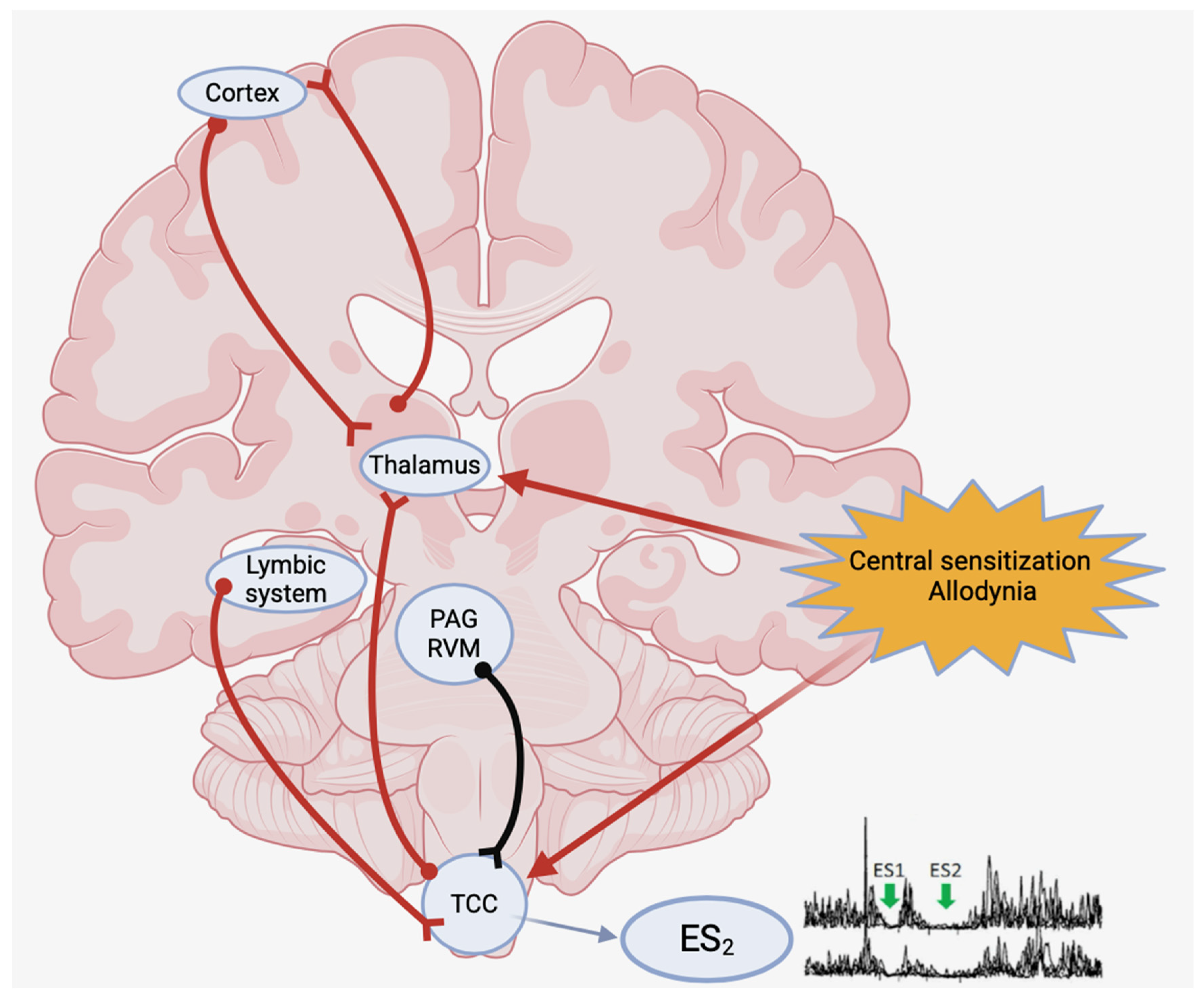
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peres, M.F.P.; Sacco, S.; Pozo-Rosich, P.; Tassorelli, C.; Ahmed, F.; Burstein, R.; Ashina, S.; Uluduz, D.; Husøy, A.K.; Steiner, T.J. Migraine is the most disabling neurological disease among children and adolescents, and second after stroke among adults: A call to action. Cephalalgia 2024, 44, 3331024241267309. [Google Scholar] [CrossRef] [PubMed]
- Coppola, G.; Di Lorenzo, C.; Schoenen, J.; Pierelli, F. Habituation and sensitization in primary headaches. J. Headache Pain 2013, 14, 65. [Google Scholar] [CrossRef] [PubMed]
- Noseda, R.; Burstein, R. Migraine pathophysiology: Anatomy of the trigeminovascular pathway and associated neurological symptoms, cortical spreading depression, sensitization, and modulation of pain. Pain 2013, 154 (Suppl. 1), S44–S53. [Google Scholar] [CrossRef] [PubMed]
- Bigal, M.E.; Lipton, R.B. Concepts and mechanisms of migraine chronification. Headache 2008, 48, 7–15. [Google Scholar] [CrossRef]
- Dodick, D.W. Migraine. Lancet 2018, 391, 1315–1330. [Google Scholar] [CrossRef]
- Burstein, R.; Yamamura, H.; Malick, A.; Strassman, A.M. Chemical stimulation of the intracranial dura induces enhanced responses to facial stimulation in brain stem trigeminal neurons. J. Neurophysiol. 1998, 9, 964–982. [Google Scholar] [CrossRef]
- Burstein, R.; Yarnitsky, D.; Goor-Aryeh, I.; Ransil, B.J.; Bajwa, Z.H. An association between migraine and cutaneous allodynia. Ann. Neurol. 2000, 47, 614–624. [Google Scholar] [CrossRef]
- Chiang, C.Y.; Dostrovsky, J.O.; Sessle, B.J. Periaqueductal gray matter and nucleus raphe magnus involvement in anterior pretectal nucleus-induced inhibition of jaw-opening reflex in rats. Brain Res. 1991, 544, 71–78. [Google Scholar] [CrossRef]
- Schoenen, J. Exteroceptive suppression of temporalis muscle activity: Methodological and physiological aspects. Cephalalgia 1993, 13, 3–10. [Google Scholar] [CrossRef]
- Fields, H.L. Pain modulation: Expectation, opioid analgesia and virtual pain. Prog. Brain Res. 2000, 122, 245–253. [Google Scholar]
- Knight, Y.E.; Goadsby, P.J. The periaqueductal grey matter modulates trigeminovascular input: A role in migraine? Neuroscience 2001, 106, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Rainville, P. Brain mechanisms of pain affect and pain modulation. Curr. Opin. Neurobiol. 2002, 12, 195–204. [Google Scholar] [CrossRef] [PubMed]
- De Tommaso, M.; Losito, L.; Difruscolo, O.; Libro, G.; Guido, M.; Livrea, P. Changes in cortical processing of pain in chronic migraine. Headache 2005, 45, 1208–1218. [Google Scholar] [CrossRef] [PubMed]
- Welch, K.M.A. Contemporary concepts of migraine pathogenesis. Neurology 2003, 61 (Suppl. 4), S2–S8. [Google Scholar] [CrossRef]
- Cruccu, G.; Deuschl, G. The clinical use of brainstem reflex and hand-muscle reflexes. Clin. Neurophysiol. 2000, 111, 371–387. [Google Scholar] [CrossRef]
- Cruccu, G.; Iannetti, G.D.; Marx, J.J.; Thoemke, F.; Truini, A.; Fitzek, S.; Galeotti, F.; Urban, P.P.; Romaniello, A.; Stoeter, P.; et al. Brainstem reflex circuits revisited. Brain 2005, 128, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Headache Classification Committee of the International Headache Society (IHS). The International Classification of Headache Disorders, 3rd edition. Cephalalgia 2018, 38, 1–211. [Google Scholar] [CrossRef]
- Göbel, H.; Schoenen, J. Exteroceptive suppression in headache research. Discussion summary. Cephalalgia 1993, 13, 20. [Google Scholar] [CrossRef]
- Zwart, J.A.; Sand, T. Exteroceptive suppression of temporalis muscle activity: A blind study of tension-type headache, migraine, and cervicogenic headache. Headache 1995, 35, 338–343. [Google Scholar] [CrossRef]
- Melhado, E.M.; Thiers Rister, H.L.; Galego, D.R.; de Oliveira, A.B.; Buttarello, I.A.; Belucio, I.S.; Oliveira Marcos, J.M.; Xavier, M.L.T.; Peres, M.F.P. Allodynia in menstrually related migraine: Score assessment by Allodynia symptom checklist (ASC-12). Headache 2020, 60, 162–170. [Google Scholar] [CrossRef]
- Mínguez-Olaondo, A.; Quintas, S.; Morollón Sánchez-Mateos, N.; López-Bravo, A.; Vila-Pueyo, M.; Grozeva, V.; Belvís, R.; Santos-Lasaosa, S.; Irimia, P. Cutaneous Allodynia in Migraine: A Narrative Review. Front. Neurol. 2022, 12, 831035. [Google Scholar] [CrossRef] [PubMed]
- Schoenen, J.; Jamart, B.; Gerard, P.; Lenarduzzi, P.; Delwaide, P.J. Exteroceptive suppression of temporalis muscle activity in chronic headache. Neurology 1987, 37, 1834–1836. [Google Scholar] [CrossRef] [PubMed]
- Aktekin, B.; Yaltkaya, K.; Ozkaynak, S.; Oguz, Y. Recovery cycle of the blink reflex and exteroceptive suppression of temporalis muscle activity in migraine and tension-type headache. Headache 2001, 41, 142–149. [Google Scholar] [CrossRef]
- Ebinger, F. Exteroceptive suppression of masseter muscle activity in juvenile migraineurs. Cephalalgia 2006, 26, 722–730. [Google Scholar] [CrossRef]
- Tataroglu, C.; Kanik, A.; Sahin, G.; Ozge, A.; Yalçinkaya, D.; Idiman, F. Exteroceptive suppression patterns of masseter and temporalis muscles in central and peripheral headache disorders. Cephalalgia 2002, 22, 444–452. [Google Scholar] [CrossRef]
- Polat, B.; Aysal, F.; Öztürk, M.; Mutluay, B.; Altunkaynak, Y.; Yilmaz, N.H.; Baybaş, S. Blink Reflex in Episodic and Chronic Migraine. Noro Psikiyatr. Ars. 2018, 55, 146–151. [Google Scholar] [CrossRef]
- Uygunoglu, U.; Gunduz, A.; Ertem, H.D.; Uluduz, D.; Saip, S.; Goksan, B.; Siva, A.; Uzun, N.; Karaali-Savrun, F.; Kızıltan, M. Deficient prepulse inhibition of blink reflex in migraine and its relation to allodynia. Neurophysiol. Clin. 2017, 47, 63–68. [Google Scholar] [CrossRef]
- Lee, M.C.; Zambreanu, L.; Menon, D.K.; Tracey, I. Identifying brain activity specifically related to the maintenance and perceptual consequence of central sensitization in humans. J. Neurosci. 2008, 28, 11642–11649. [Google Scholar] [CrossRef]
- Gunduz, A.; Valls-Solé, J.; Serranová, T.; Coppola, G.; Kofler, M.; Jääskeläinen, S.K. The blink reflex and its modulation—Part 2: Pathophysiology and clinical utility. Clin. Neurophysiol. 2024, 160, 75–94. [Google Scholar] [CrossRef]
- Schoenen, J.; Raubüchl, O.; Sianard, J. Pharmacologic modulation of temporalis exteroceptive silent periods in healthy volunteers. Cephalalgia 1991, 11 (Suppl. 11), 16–17. [Google Scholar] [CrossRef]
- Ferrari, M.D.; Saxena, P.R. On serotonin and migraine: A clinical and pharmacological review. Cephalalgia 1993, 13, 151–165. [Google Scholar] [CrossRef]
- Rota, E.; Aguggia, M.; Immovilli, P.; Morelli, N.; Renosio, D.; Barbanera, A. Change in the second exteroceptive suppression period of the temporalis muscle during erenumab treatment. Naunyn Schmiedebergs Arch. Pharmacol. 2022, 395, 607–611. [Google Scholar] [CrossRef] [PubMed]
- Casillo, F.; Sebastianelli, G.; Di Renzo, A.; Cioffi, E.; Parisi, V.; Di Lorenzo, C.; Serrao, M.; Coppola, G. The monoclonal CGRP-receptor blocking antibody erenumab has different effects on brainstem and cortical sensory-evoked responses. Cephalalgia 2022, 42, 1236–1245. [Google Scholar] [CrossRef] [PubMed]
- Thiele, A.; Klehr, L.; Strauß, S.; Angermaier, A.; Schminke, U.; Kronenbuerger, M.; Naegel, S.; Fleischmann, R. Preventive treatment with CGRP monoclonal antibodies restores brain stem habituation deficits and excitability to painful stimuli in migraine: Results from a prospective case-control study. J. Headache Pain 2021, 22, 149. [Google Scholar] [CrossRef] [PubMed]
- Danno, D.; Imai, N.; Kitamura, S.; Ishizaki, K.; Kikui, S.; Takeshima, T. Efficacy of galcanezumab in migraine central sensitization. Sci. Rep. 2024, 14, 21824. [Google Scholar] [CrossRef]
- Kaube, H.; Katsarava, Z.; Przywara, S.; Drepper, J.; Ellrich, J.; Diener, H.C. Acute migraine headache: Possible sensitization of neurons in the spinal trigeminal nucleus? Neurology 2002, 58, 1234–1238. [Google Scholar] [CrossRef]
- Marucco, E.; Lisicki, M.; Magis, D. Electrophysiological Characteristics of the Migraine Brain: Current Knowledge and Perspectives. Curr. Med. Chem. 2019, 26, 6222–6235. [Google Scholar] [CrossRef]
- Coppola, G.; Di Lorenzo, C.; Parisi, V.; Lisicki, M.; Serrao, M.; Pierelli, F. Clinical neurophysiology of migraine with aura. J. Headache Pain 2019, 20, 42. [Google Scholar] [CrossRef]

{kind=link}
| Allodynic vs. Controls | Allodynic vs. Non-allodynic | Migraineurs vs. Controls | Controls (N°: 23) | Non-Allodynic (N°: 25) | Allodynic (N°: 24) | Migraineurs (Total N°: 49) | |
|---|---|---|---|---|---|---|---|
| p: 0.63 | p: 0.51 | p: 0.44 | 41 (20–61) | 43 (18–62) | 38 (22–59) | 40 (18–62) | Age (average, years) (min–max value) |
| - | p: 0.63 | - | - | 12 (2–26) | 16 (3–30) | 14 (2–30) | Headache monthly frequency (min–max value) |
| p: 0.31 | p: 0.75 | p: 0.62 | 13.9 ± 1.3 (11–16.8) | 12.6 ± 1.9 (9.6–15.2) | 13.1 ± 1.3 (11–15.2) | 12.8 ± 1.5 (9.6–16.2) | ES1 latency (average, msec) ± SD (min–max value) |
| p: 0.21 | p: 0.94 | p: 0.12 | 14.1 ± 3.1 (7.8–17.6) | 13.8 ± 3.0 (10.4–18.5) | 14.9 ± 4.7 (10–17.4) | 14.4 ± 3.4 (10–18.5) | ES1 duration (average, msec) ± SD (min–max value) |
| p:0.44 | p: 0.69 | p: 0.61 | 56.8 ± 7.2 (49.9–66) | 60.1 ± 9.7 (55.1–76.9) | 62.1 ± 11.1 (42.2–98.8) | 61.5 ± 8.5 (42.2–98.8) | ES2 latency (average, msec) ± SD (min–max value) |
| p: 0.040 | p: 0.046 | p: 0.78 | 25.6 ± 7.4 (21.2–32.8) | 26.5 ± 6.4 (21.4–28.2) | 31.8 ± 10.0 (24–43.8) | 28.2 ± 7.7 (21.4–43.8) | ES2 duration (average, msec) ± SD (min–max value) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rota, E.; Immovilli, P.; Aguggia, M.; Saracco, M.G.; Ghiglione, E.; Melotti, A.; Morelli, N. The Second Exteroceptive Suppression Period of the Temporalis Muscle Is Altered in Migraine Patients with Allodynia. Neurol. Int. 2025, 17, 76. https://doi.org/10.3390/neurolint17050076
Rota E, Immovilli P, Aguggia M, Saracco MG, Ghiglione E, Melotti A, Morelli N. The Second Exteroceptive Suppression Period of the Temporalis Muscle Is Altered in Migraine Patients with Allodynia. Neurology International. 2025; 17(5):76. https://doi.org/10.3390/neurolint17050076
Chicago/Turabian StyleRota, Eugenia, Paolo Immovilli, Marco Aguggia, Maria Gabriella Saracco, Elisabetta Ghiglione, Antonella Melotti, and Nicola Morelli. 2025. "The Second Exteroceptive Suppression Period of the Temporalis Muscle Is Altered in Migraine Patients with Allodynia" Neurology International 17, no. 5: 76. https://doi.org/10.3390/neurolint17050076
APA StyleRota, E., Immovilli, P., Aguggia, M., Saracco, M. G., Ghiglione, E., Melotti, A., & Morelli, N. (2025). The Second Exteroceptive Suppression Period of the Temporalis Muscle Is Altered in Migraine Patients with Allodynia. Neurology International, 17(5), 76. https://doi.org/10.3390/neurolint17050076