
Neuronal Excitation Induces Tau Protein Dephosphorylation via Protein Phosphatase 1 Activation to Promote Its Binding with Stable Microtubules

{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Antibodies
2.3. Primary Cortical Neuronal Culture
2.4. Biochemical Analysis of Hippocampus
2.5. Microtubule Fractionation
2.6. Immunoblot Analyses
2.7. Phosphatase Assay
2.8. Statistical Analyses
3. Results
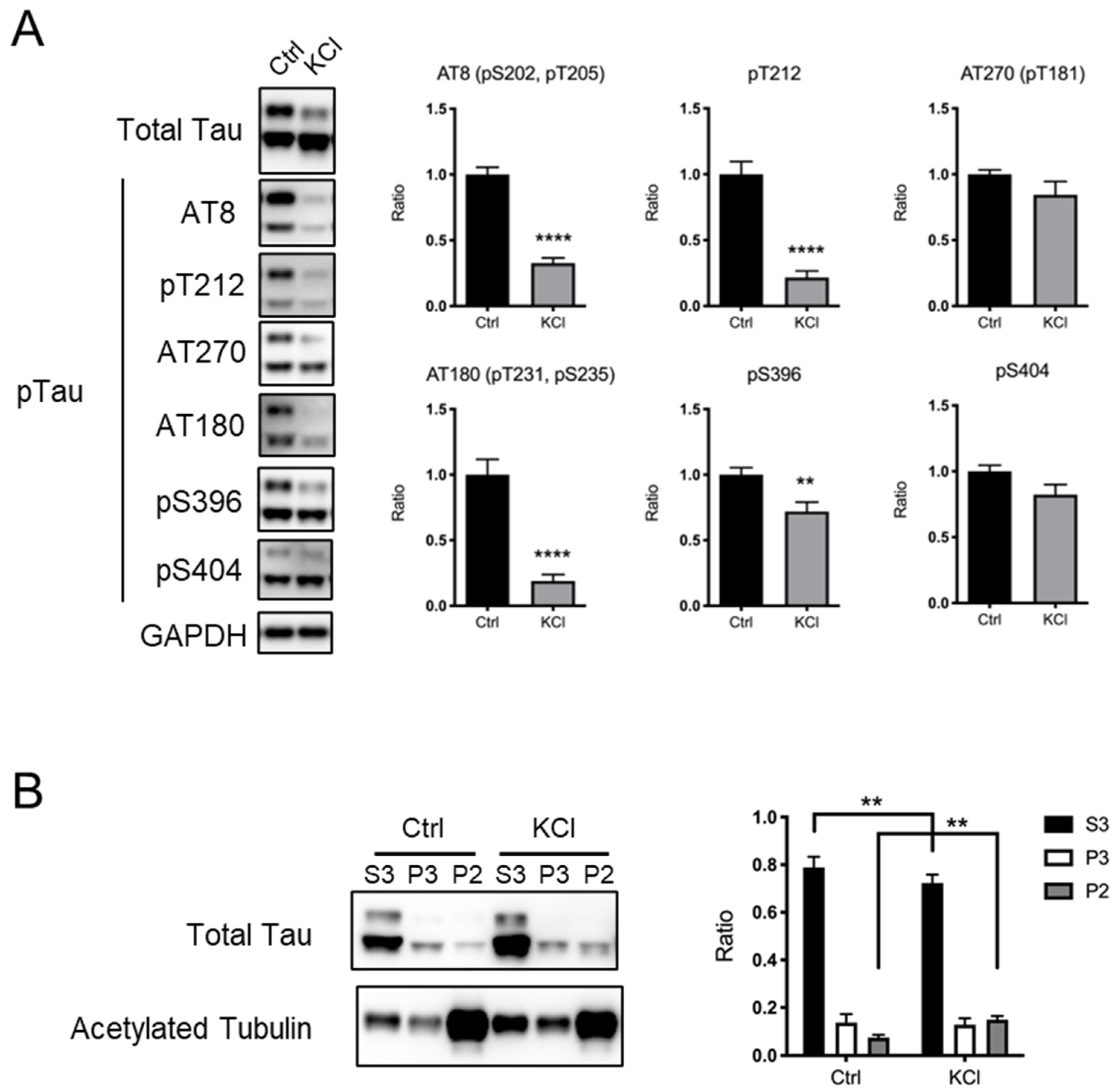
3.1. KCl Treatment Led to Tau Protein Dephosphorylation in Primary Cortical Neurons
3.2. Exposure to Novel Environment Decreases the Level of Phosphorylated Tau Protein
3.3. Exposure to a Novel Environment Increases the Levels of Tau Protein Fractionated with Stable Microtubules
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buée, L.; Bussière, T.; Buée-Scherrer, V.; Delacourte, A.; Hof, P.R. Tau protein isoforms, phosphorylation and role in neurodegenerative disorders. Brain Res. Brain Res. Rev. 2000, 33, 95–130. [Google Scholar] [CrossRef] [PubMed]
- Pires, G.; McElligott, S.; Drusinsky, S.; Halliday, G.; Potier, M.C.; Wisniewski, T.; Drummond, E. Secernin-1 is a novel phosphorylated tau binding protein that accumulates in Alzheimer’s disease and not in other tauopathies. Acta Neuropathol. Commun. 2019, 7, 195. [Google Scholar] [CrossRef] [PubMed]
- Lee, V.M.; Goedert, M.; Trojanowski, J.Q. Neurodegenerative tauopathies. Annu. Rev. Neurosci. 2001, 24, 1121–1159. [Google Scholar] [CrossRef] [PubMed]
- Gustke, N.; Steiner, B.; Mandelkow, E.M.; Biernat, J.; Meyer, H.E.; Goedert, M.; Mandelkow, E. The Alzheimer-like phosphorylation of tau protein reduces microtubule binding and involves Ser-Pro and Thr-Pro motifs. FEBS Lett. 1992, 307, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Biernat, J.; Gustke, N.; Drewes, G.; Mandelkow, E.M.; Mandelkow, E. Phosphorylation of Ser262 strongly reduces binding of tau to microtubules: Distinction between PHF-like immunoreactivity and microtubule binding. Neuron 1993, 11, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.H.; Johnson, G.V. Glycogen synthase kinase 3beta phosphorylates tau at both primed and unprimed sites. Differential impact on microtubule binding. J. Biol. Chem. 2003, 278, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Wagner, U.; Utton, M.; Gallo, J.M.; Miller, C.C. Cellular phosphorylation of tau by GSK-3 beta influences tau binding to microtubules and microtubule organisation. J. Cell Sci. 1996, 109, 1537–1543. [Google Scholar] [CrossRef] [PubMed]
- Bading, H.; Ginty, D.D.; Greenberg, M.E. Regulation of gene expression in hippocampal neurons by distinct calcium signaling pathways. Science 1993, 260, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Hardingham, G.E.; Arnold, F.J.; Bading, H. Nuclear calcium signaling controls CREB-mediated gene expression triggered by synaptic activity. Nat. Neurosci. 1993, 4, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Gong, R.; Park, C.S.; Abbassi, N.R.; Tang, S.J. Roles of glutamate receptors and the mammalian target of rapamycin (mTOR) signaling pathway in activity-dependent dendritic protein synthesis in hippocampal neurons. J. Biol. Chem. 2006, 281, 18802–18815. [Google Scholar] [CrossRef]
- Zhou, X.P.; Wu, K.Y.; Liang, B.; Fu, X.Q.; Luo, Z.G. TrkB-mediated activation of geranylgeranyltransferase I promotes dendritic morphogenesis. Proc. Natl. Acad. Sci. USA 2008, 105, 17181–17186. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Neveu, P.; Kosik, K.S. A coordinated local translational control point at the synapse involving relief from silencing and MOV10 degradation. Neuron 2009, 64, 871–884. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, C.; Takemori, H.; Zhou, Y.; Xiong, Z.Q. TORC1 regulates activity-dependent CREB-target gene transcription and dendritic growth of developing cortical neurons. J. Neurosci. 2009, 29, 2334–2343. [Google Scholar] [CrossRef] [PubMed]
- Fujii, H.; Matsubara, K.; Sakai, K.; Ito, M.; Ohno, K.; Ueda, M.; Yamamoto, A. Dopaminergic differentiation of stem cells from human deciduous teeth and their therapeutic benefits for Parkinsonian rats. Brain Res. 2015, 1613, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Atsumi, Y.; Iwata, R.; Kimura, H.; Vanderhaeghen, P.; Yamamoto, N.; Sugo, N. Repetitive CREB-DNA interactions at gene loci predetermined by CBP induce activity-dependent gene expression in human cortical neurons. Cell Rep. 2024, 43, 113576. [Google Scholar] [CrossRef] [PubMed]
- Adamec, E.; Mercken, M.; Beermann, M.L.; Didier, M.; Nixon, R.A. Acute rise in the concentration of free cytoplasmic calcium leads to dephosphorylation of the microtubule-associated protein tau. Brain Res. 1997, 757, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, N.V.; Desjardins, A.; Leclerc, N. Tau secretion is correlated to an increase of Golgi dynamics. PLoS ONE 2017, 12, e0178288. [Google Scholar] [CrossRef] [PubMed]
- Bramblett, G.T.; Goedert, M.; Jakes, R.; Merrick, S.E.; Trojanowski, J.Q.; Lee, V.M. Abnormal tau phosphorylation at Ser396 in Alzheimer’s disease recapitulates development and contributes to reduced microtubule binding. Neuron 1993, 10, 1089–1099. [Google Scholar] [CrossRef] [PubMed]
- Schneider, A.; Biernat, J.; von Bergen, M.; Mandelkow, E.; Mandelkow, E.M. Phosphorylation that detaches tau protein from microtubules (Ser262, Ser214) also protects it against aggregation into Alzheimer paired helical filaments. Biochemistry 1999, 38, 3549–3558. [Google Scholar] [CrossRef] [PubMed]
- Alonso, A.D.; Di Clerico, J.; Li, B.; Corbo, C.P.; Alaniz, M.E.; Grundke-Iqbal, I.; Iqbal, K. Phosphorylation of tau at Thr212, Thr231, and Ser262 combined causes neurodegeneration. J. Biol. Chem. 2010, 285, 30851–30860. [Google Scholar] [CrossRef]
- Noble, W.; Hanger, D.P.; Miller, C.C.; Lovestone, S. The importance of tau phosphorylation for neurodegenerative diseases. Front. Neurol. 2013, 4, 83. [Google Scholar] [CrossRef] [PubMed]
- Haj-Yahya, M.; Gopinath, P.; Rajasekhar, K.; Mirbaha, H.; Diamond, M.I.; Lashuel, H.A. Site-Specific Hyperphosphorylation Inhibits, Rather than Promotes, Tau Fibrillization, Seeding Capacity, and Its Microtubule Binding. Angew. Chem. Int. Ed. Engl. 2020, 59, 4059–4067. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Ocádiz, R.; Trippa, M.; Zhang, C.L.; Posani, L.; Cocco, S.; Monasson, R.; Schmidt-Hieber, C. A synaptic signal for novelty processing in the hippocampus. Nat. Commun. 2022, 13, 4122. [Google Scholar] [CrossRef] [PubMed]
- Canet, G.; Rocaboy, E.; Laliberté, F.; Boscher, E.; Guisle, I.; Diego-Diaz, S.; Fereydouni-Forouzandeh, P.; Whittington, R.A.; Hébert, S.S.; Pernet, V.; et al. Temperature-induced Artifacts in Tau Phosphorylation: Implications for Reliable Alzheimer’s Disease Research. Exp. Neurobiol. 2023, 32, 423–440. [Google Scholar] [CrossRef]
- Mitsuhashi, S.; Matsuura, N.; Ubukata, M.; Oikawa, H.; Shima, H.; Kikuchi, K. Tautomycetin is a novel and specific inhibitor of serine/threonine protein phosphatase type 1, PP1. Biochem Biophys Res Commun. 2001, 287, 328–331. [Google Scholar] [CrossRef] [PubMed]
- Hagita, A.; Wada-Kakuda, S.; Nobuhara, M.; Kakuda, N.; Miyasaka, T. Quantitative fractionation of tissue microtubules with distinct biochemical properties reflecting their stability and lability. Biochem. Biophys. Res. Commun. 2021, 560, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Planel, E.; Miyasaka, T.; Launey, T.; Chui, D.H.; Tanemura, K.; Sato, S.; Murayama, O.; Ishiguro, K.; Tatebayashi, Y.; Takashima, A. Alterations in glucose metabolism induce hypothermia leading to tau hyperphosphorylation through differential inhibition of kinase and phosphatase activities: Implications for Alzheimer’s disease. J. Neurosci. 2004, 24, 2401–2411. [Google Scholar] [CrossRef]
- Stoothoff, W.H.; Johnson, G.V. Tau phosphorylation: Physiological and pathological consequences. Biochim. Biophys. Acta. 2005, 1739, 280–297. [Google Scholar] [CrossRef] [PubMed]
- Plattner, F.; Angelo, M.; Giese, K.P. The roles of cyclin-dependent kinase 5 and glycogen synthase kinase 3 in tau hyperphosphorylation. J. Biol. Chem. 2006, 281, 25457–25465. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Yamashita, S.; Fukuda, T.; Park, J.M.; Murayama, M.; Mizoroki, T.; Yoshiike, Y.; Sahara, N.; Takashima, A. Hyperphosphorylated tau in parahippocampal cortex impairs place learning in aged mice expressing wild-type human tau. EMBO J. 2007, 26, 5143–5152. [Google Scholar] [CrossRef]
- Verwer, R.W.; Sluiter, A.A.; Balesar, R.A.; Baayen, J.C.; Noske, D.P.; Dirven, C.M.; Wouda, J.; van Dam, A.M.; Lucassen, P.J.; Swaab, D.F. Mature astrocytes in the adult human neocortex express the early neuronal marker doublecortin. Brain. 2007, 130, 3321–3335. [Google Scholar] [CrossRef] [PubMed]
- Hanger, D.P.; Anderton, B.H.; Noble, W. Tau phosphorylation: The therapeutic challenge for neurodegenerative disease. Trends Mol. Med. 2009, 15, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Shahpasand, K.; Uemura, I.; Saito, T.; Asano, T.; Hata, K.; Shibata, K.; Toyoshima, Y.; Hasegawa, M.; Hisanaga, S. Regulation of mitochondrial transport and inter-microtubule spacing by tau phosphorylation at the sites hyperphosphorylated in Alzheimer’s disease. J. Neurosci. 2012, 32, 2430–2441. [Google Scholar] [CrossRef] [PubMed]
- Guisle, I.; Gratuze, M.; Petry, S.; Morin, F.; Keraudren, R.; Whittington, R.A.; Hébert, S.S.; Mongrain, V.; Planel, E. Circadian and sleep/wake-dependent variations in tau phosphorylation are driven by temperature. Sleep 2020, 43, zsz266. [Google Scholar] [CrossRef] [PubMed]
- Brion, J.P.; Smith, C.; Couck, A.M.; Gallo, J.M.; Anderton, B.H. Developmental changes in tau phosphorylation: Fetal tau is transiently phosphorylated in a manner similar to paired helical filament-tau characteristic of Alzheimer’s disease. J. Neurochem. 1993, 61, 2071–2080. [Google Scholar] [CrossRef] [PubMed]
- Arendt, T.; Stieler, J.; Strijkstra, A.M.; Hut, R.A.; Rüdiger, J.; Van der Zee, E.A.; Harkany, T.; Holzer, M.; Härtig, W. Reversible paired helical filament-like phosphorylation of tau is an adaptive process associated with neuronal plasticity in hibernating animals. J. Neurosci. 2003, 23, 6972–6981. [Google Scholar] [CrossRef]
- Kenessey, A.; Yen, S.H. The extent of phosphorylation of fetal tau is comparable to that of PHF-tau from Alzheimer paired helical filaments. Brain Res. 1993, 629, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Planel, E.; Richter, K.E.; Nolan, C.E.; Finley, J.E.; Liu, L.; Wen, Y.; Krishnamurthy, P.; Herman, M.; Wang, L.; Schachter, J.B.; et al. Anesthesia leads to tau hyperphosphorylation through inhibition of phosphatase activity by hypothermia. J. Neurosci. 2007, 27, 3090–3097. [Google Scholar] [CrossRef] [PubMed]
- Su, B.; Wang, X.; Drew, K.L.; Perry, G.; Smith, M.A.; Zhu, X. Physiological regulation of tau phosphorylation during hibernation. J. Neurochem. 2008, 105, 2098–2108. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Run, X.; Liang, Z.; Li, Y.; Liu, F.; Liu, Y.; Iqbal, K.; Grundke-Iqbal, I.; Gong, C.X. Developmental regulation of tau phosphorylation, tau kinases, and tau phosphatases. J. Neurochem. 2009, 108, 1480–1494. [Google Scholar] [CrossRef]
- Stieler, J.T.; Bullmann, T.; Kohl, F.; Tøien, Ø.; Brückner, M.K.; Härtig, W.; Barnes, B.M.; Arendt, T. The physiological link between metabolic rate depression and tau phosphorylation in mammalian hibernation. PLoS ONE 2011, 6, e14530. [Google Scholar] [CrossRef] [PubMed]
- Yagishita, S.; Suzuki, S.; Yoshikawa, K.; Iida, K.; Hirata, A.; Suzuki, M.; Takashima, A.; Maruyama, K.; Hirasawa, A.; Awaji, T. Treatment of intermittent hypoxia increases phosphorylated tau in the hippocampus via biological processes common to aging. Mol. Brain 2017, 10, 2. [Google Scholar] [CrossRef] [PubMed]
- Guisle, I.; Canet, G.; Pétry, S.; Fereydouni-Forouzandeh, P.; Morin, F.; Kérauden, R.; Whittington, R.A.; Calon, F.; Hébert, S.S.; Planel, E. Sauna-like conditions or menthol treatment reduce tau phosphorylation through mild hyperthermia. Neurobiol. Aging 2022, 113, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Prokop, S.; Giasson, B.I. “Don’t Phos Over Tau”: Recent developments in clinical biomarkers and therapies targeting tau phosphorylation in Alzheimer’s disease and other tauopathies. Mol. Neurodegener. 2021, 16, 37. [Google Scholar] [CrossRef] [PubMed]
- Pluta, R.; Czuczwar, S.J. Trans- and Cis-Phosphorylated Tau Protein: New Pieces of the Puzzle in the Development of Neurofibrillary Tangles in Post-Ischemic Brain Neurodegeneration of the Alzheimer’s Disease-like Type. Int. J. Mol. Sci. 2024, 25, 3091. [Google Scholar] [CrossRef] [PubMed]
- Winder, D.G.; Sweatt, J.D. Roles of serine/threonine phosphatases in hippocampal synaptic plasticity. Nat. Rev. Neurosci. 2001, 2, 461–474. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, A.; Taleski, G.; Sontag, E. The protein serine/threonine phosphatases PP2A, PP1 and calcineurin: A triple threat in the regulation of the neuronal cytoskeleton. Mol. Cell Neurosci. 2017, 84, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Aggen, J.B.; Nairn, A.C.; Chamberlin, R. Regulation of protein phosphatase-1. Chem. Biol. 2000, 7, R13–R23. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yagishita, S.; Shibata, M.; Furuno, A.; Wakatsuki, S.; Araki, T. Neuronal Excitation Induces Tau Protein Dephosphorylation via Protein Phosphatase 1 Activation to Promote Its Binding with Stable Microtubules. Neurol. Int. 2024, 16, 653-662. https://doi.org/10.3390/neurolint16030049
Yagishita S, Shibata M, Furuno A, Wakatsuki S, Araki T. Neuronal Excitation Induces Tau Protein Dephosphorylation via Protein Phosphatase 1 Activation to Promote Its Binding with Stable Microtubules. Neurology International. 2024; 16(3):653-662. https://doi.org/10.3390/neurolint16030049
Chicago/Turabian StyleYagishita, Sosuke, Megumi Shibata, Akiko Furuno, Shuji Wakatsuki, and Toshiyuki Araki. 2024. "Neuronal Excitation Induces Tau Protein Dephosphorylation via Protein Phosphatase 1 Activation to Promote Its Binding with Stable Microtubules" Neurology International 16, no. 3: 653-662. https://doi.org/10.3390/neurolint16030049
APA StyleYagishita, S., Shibata, M., Furuno, A., Wakatsuki, S., & Araki, T. (2024). Neuronal Excitation Induces Tau Protein Dephosphorylation via Protein Phosphatase 1 Activation to Promote Its Binding with Stable Microtubules. Neurology International, 16(3), 653-662. https://doi.org/10.3390/neurolint16030049