
In Vivo Pathogenicity of Methicillin-Susceptible Staphylococcus aureus Strains Carrying Panton–Valentine Leukocidin Gene
 ,
,  , ,
, ,
Abstract
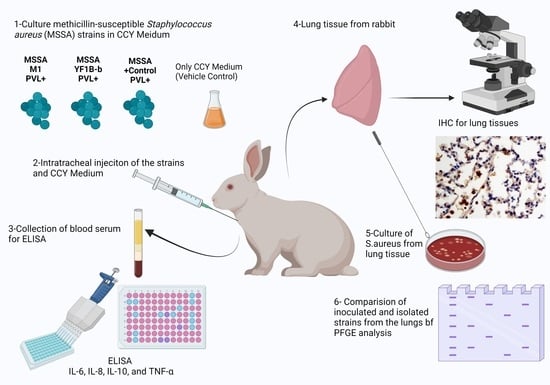
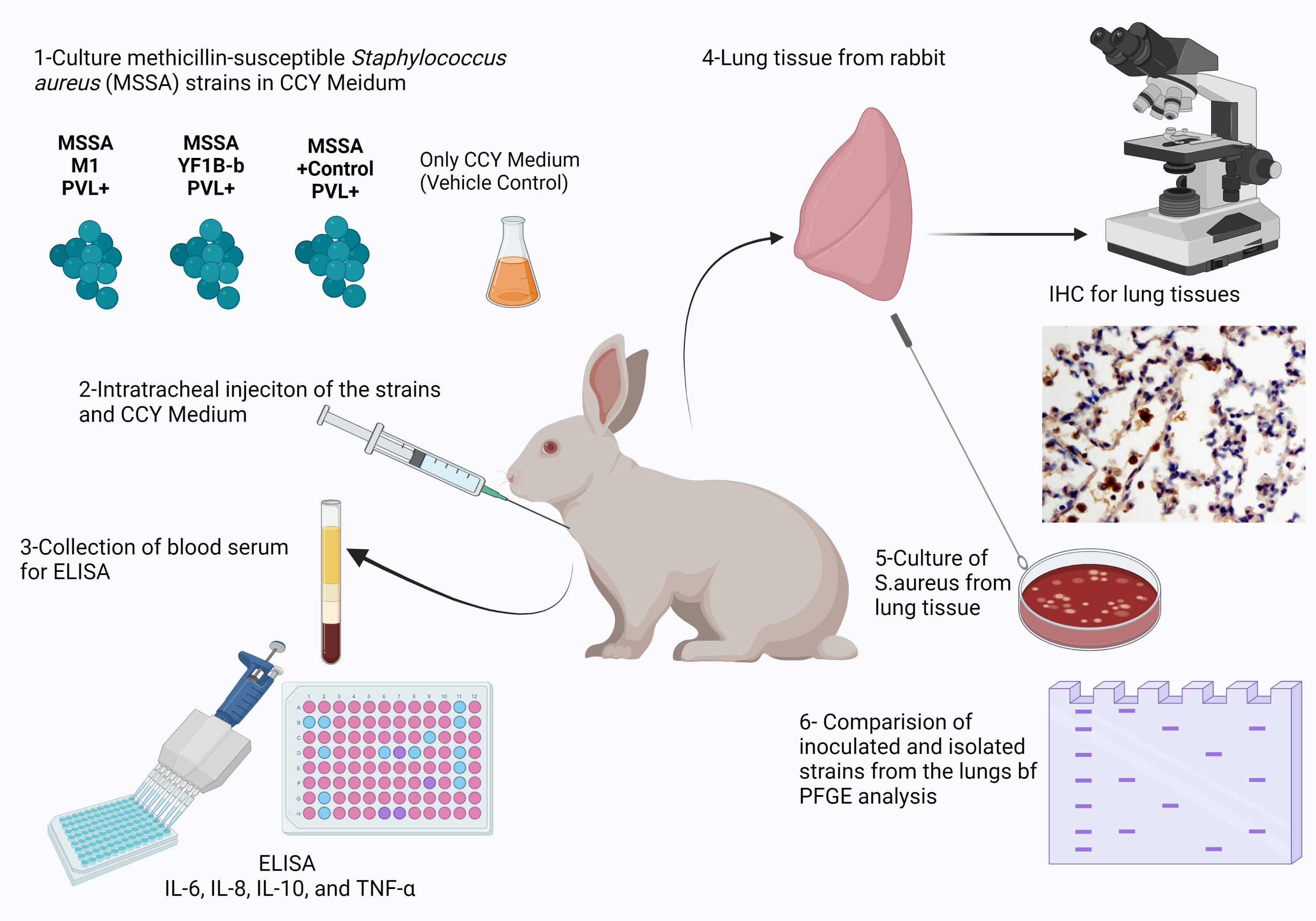
1. Introduction
2. Materials and Methods
2.1. Ethical Aspects
2.2. Test and Control Strains
2.3. Preparation of Culture Medium
2.4. Animal Model
2.5. Histopathology and Immunohistochemistry
2.6. Cytokine Levels
2.7. Isolation of Inoculated Strains
2.8. Pulsed-Field Gel Electrophoresis
2.9. Statistical Analysis
3. Results
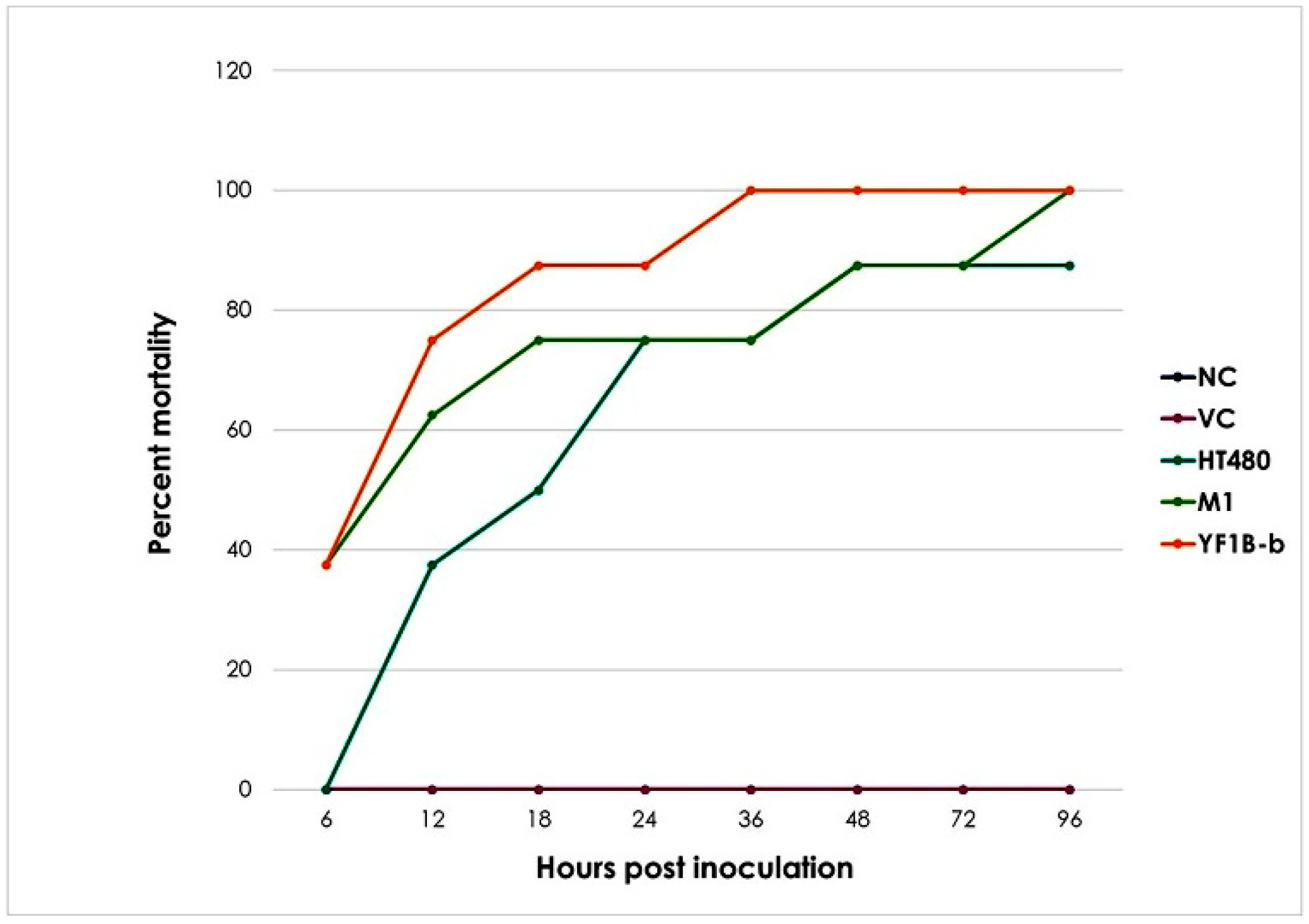
3.1. Animal Response
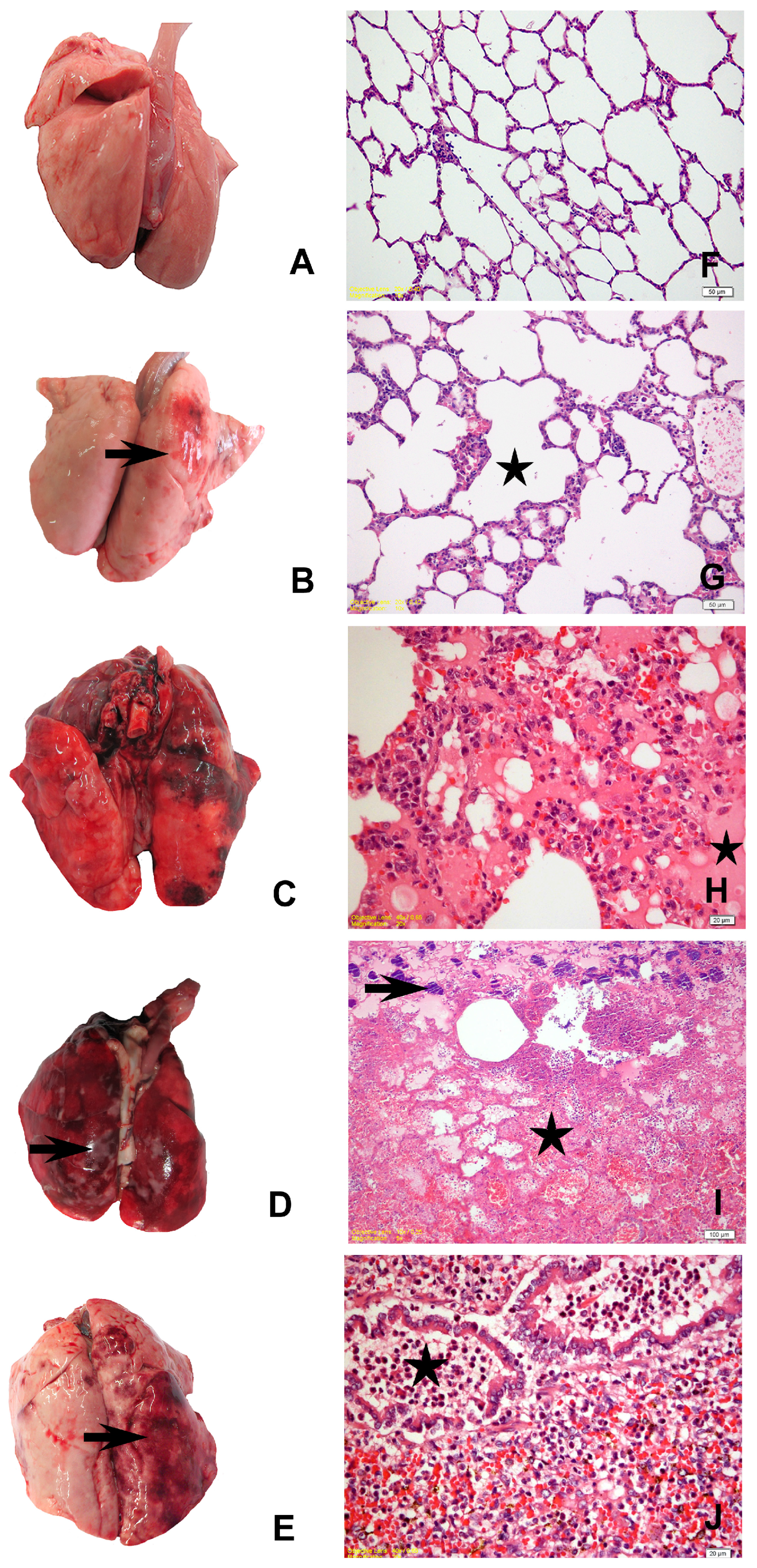
3.2. Gross Pathological Findings
3.3. Histopathological Findings
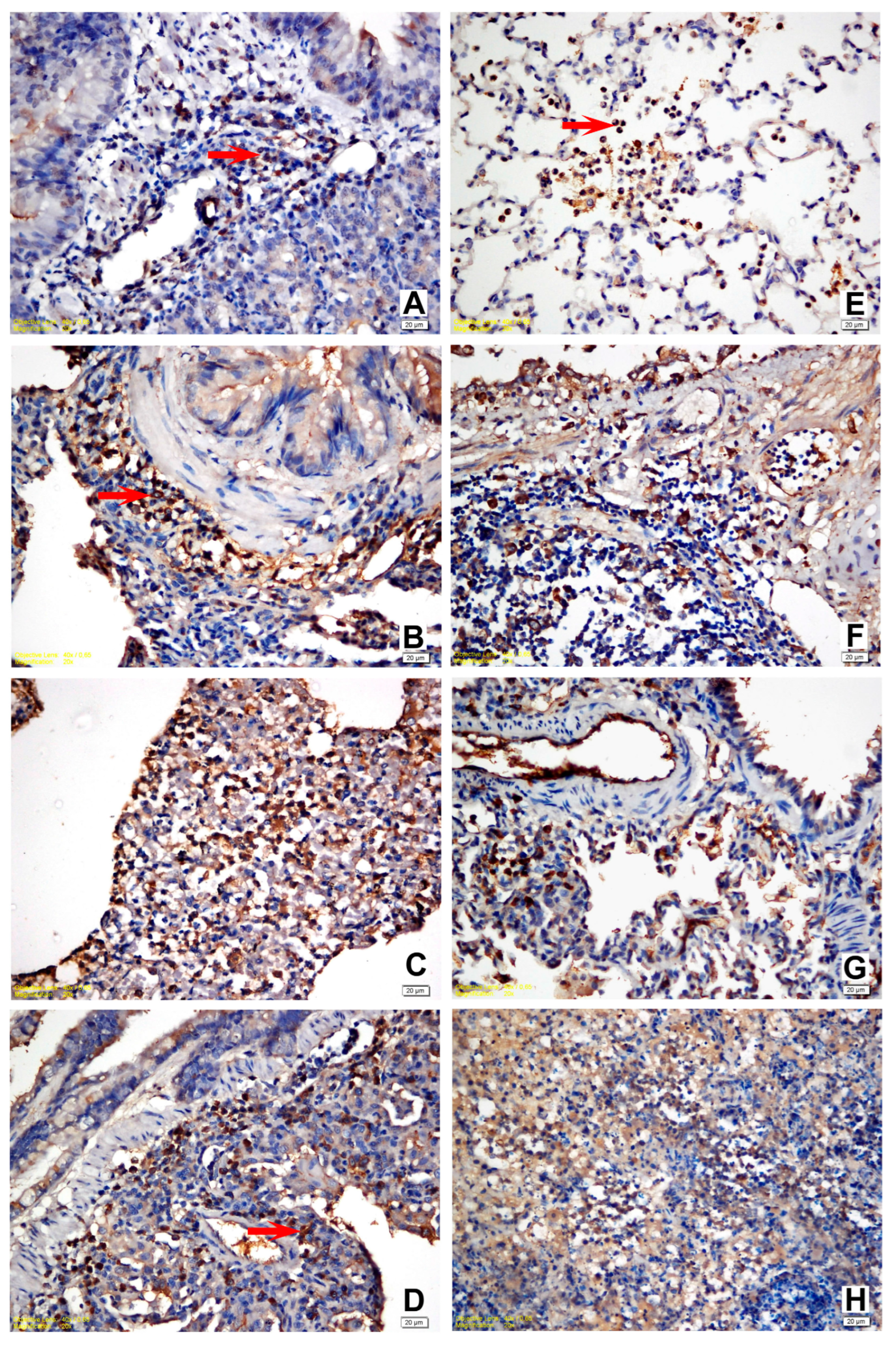
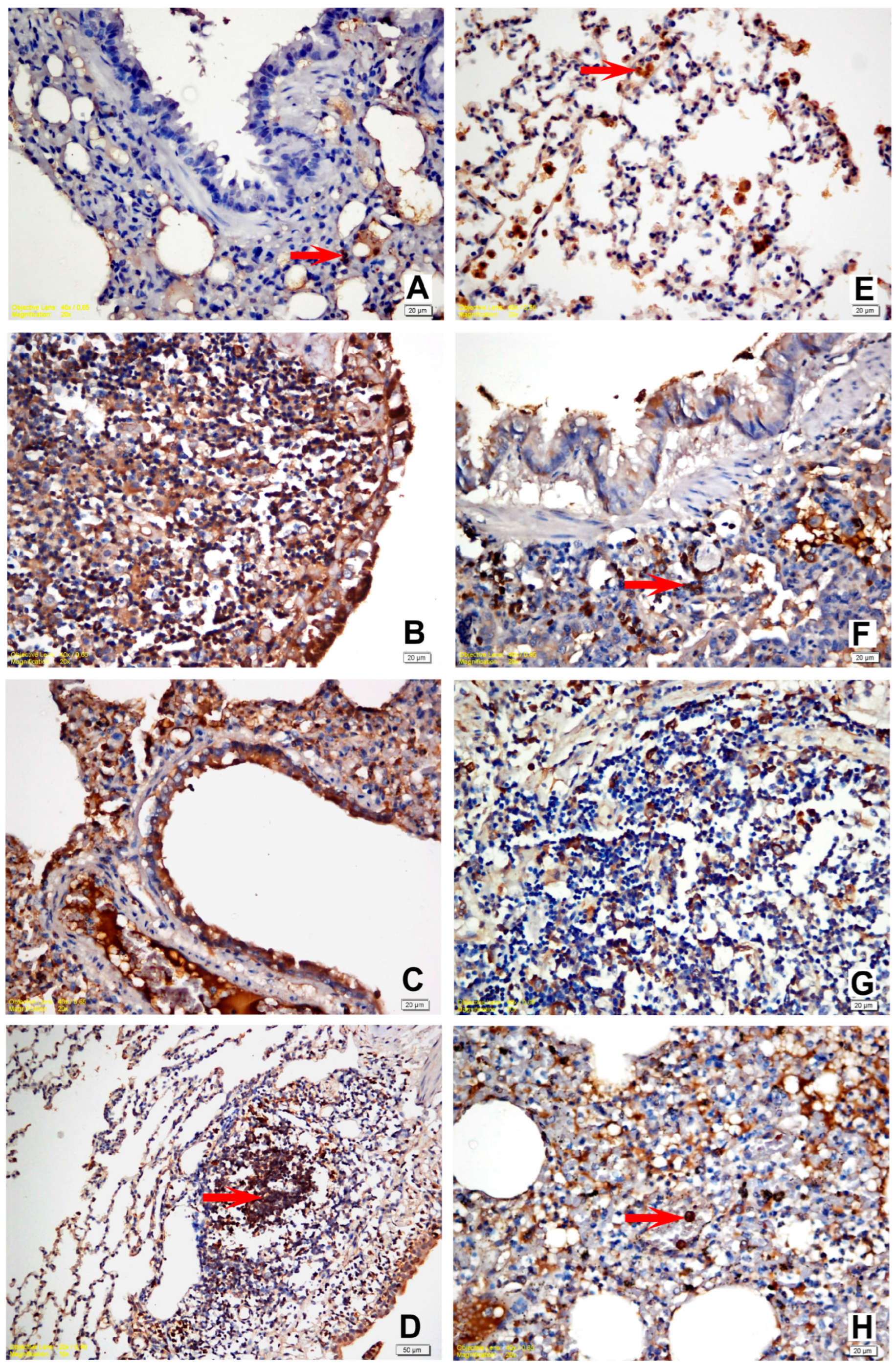
3.4. Immunohistochemical Findings
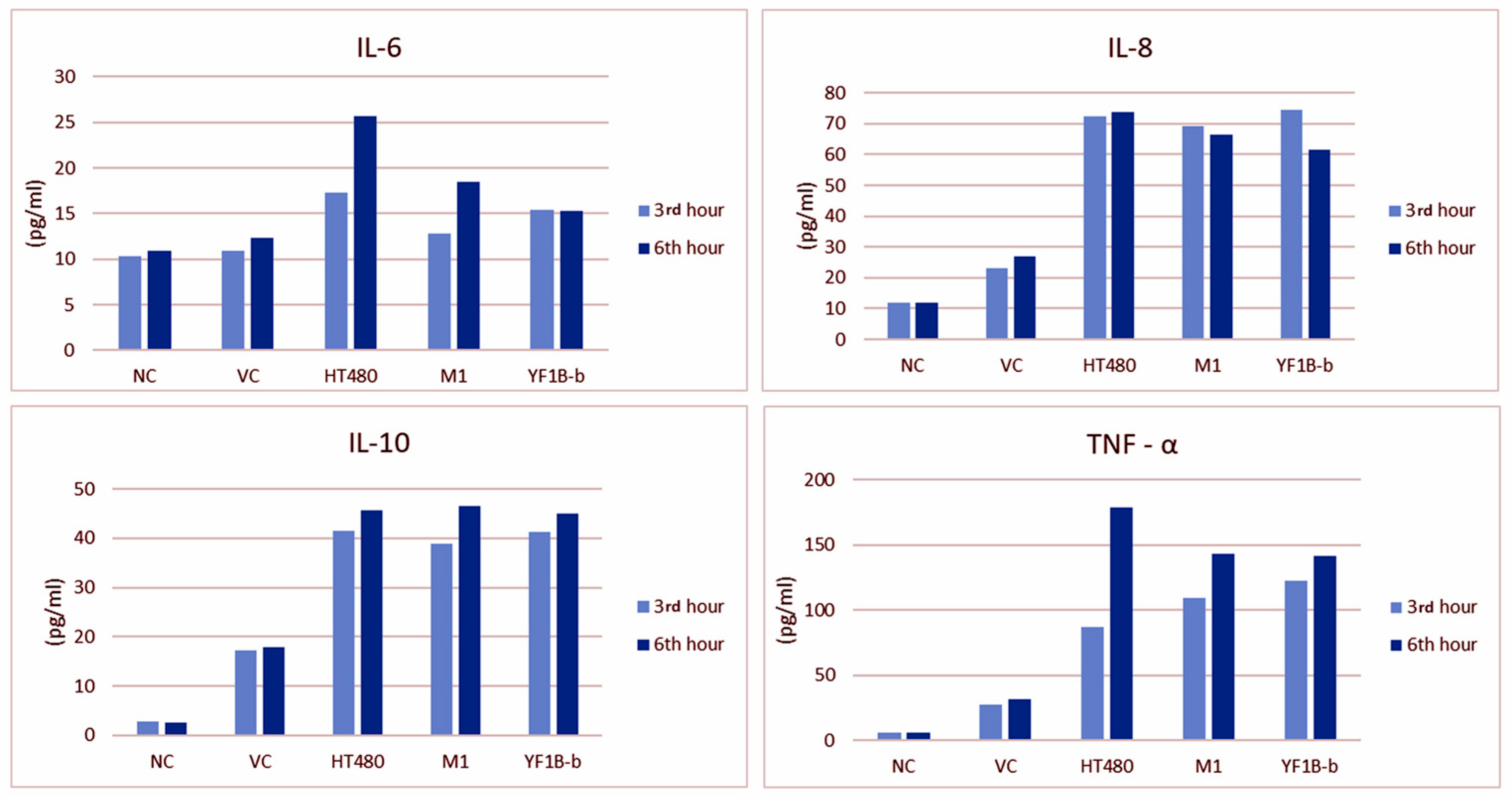
3.5. ELISA Results
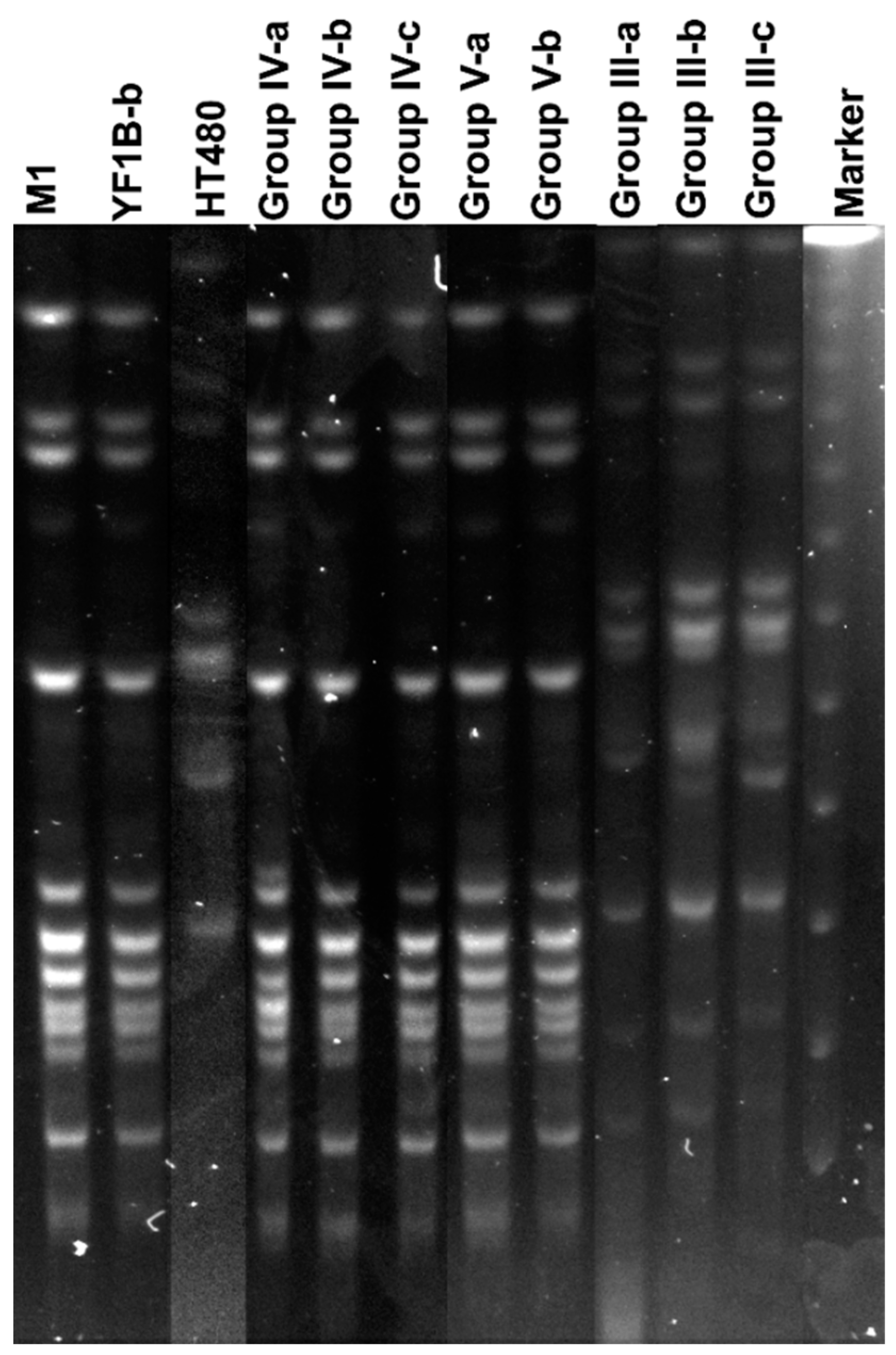
3.6. Isolation of Inoculated Strains and PFGE Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jablonski, L.M.; Bohach, G.A. Staphylococcus aureus. In Food Microbiology Fundamentals and Frontiers, 1st ed.; Doyle, M.P., Beuchat, L.R., Eds.; ASM Pres: Washington, DC, USA, 2001; pp. 411–434. [Google Scholar]
- Sutherland, J.; Varnam, A. Enterotoksin producing Staphylococcus, Shigella, Yersinia, Vibrio, Aeromonas and Plesiomonas. In Foodborne Pathogens; Blackburn, C.W., McClure, P.J., Eds.; CRC Press: Washington, DC, USA, 2002; pp. 384–415. [Google Scholar]
- Ramos, A.; Ley, L.; Muñez, E.; Videl, A.; Sánchez, I. Brain abscess due to Panton–Valentine leukocidin-positive Staphylococcus aureus. Infection 2009, 37, 365–367. [Google Scholar] [CrossRef] [PubMed]
- Kadariya, J.; Smith, T.C.; Thapaliya, D. Staphylococcus aureus and staphylococcal food-borne disease: An ongoing challenge in public health. BioMed Res. Int. 2014, 2014, 827965. [Google Scholar] [CrossRef] [PubMed]
- Fetsch, S.; Johler, S. Staphylococcus aureus as a foodborne pathogen. Curr. Clin. Microbiol. Rep. 2018, 5, 88–96. [Google Scholar] [CrossRef]
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati, M.; Holland, T.L.; Fowler, V.G. Methicillin-resistant Staphylococcus aureus: An overview of basic and clinical research. Nat. Rev. Microbiol. 2019, 17, 203–218. [Google Scholar] [CrossRef]
- Reyes-Robles, T.; Torres, V.J. Staphylococcus aureus Pore-Forming Toxins. Curr. Top. Microbiol. Immunol. 2017, 409, 121–144. [Google Scholar] [CrossRef]
- Shumba, P.; Mairpady Shambat, S.; Siemens, N. The role of streptococcal and staphylococcal exotoxins and proteases in human necrotizing soft tissue infections. Toxins 2016, 11, 332. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, T.; Zou, X.; Wu, S.; Zhu, J. Panton-valentine leucocidin carrying Staphylococcus aureus causing necrotizing pneumonia inactivates the JAK/STAT signaling pathway and increases the expression of inflammatory cytokines. Infect. Genet. Evol. 2020, 86, 104582. [Google Scholar] [CrossRef]
- Zhao, H.; Hu, F.; Jin, S.; Xu, X.; Zou, Y.; Ding, B.; He, C.; Gong, F.; Liu, Q. Typing of Panton-Valentine leukocidin-encoding phages and lukSF-PV gene sequence variation in Staphylococcus aureus from China. Front. Microbiol. 2016, 7, 1200. [Google Scholar] [CrossRef]
- Tong, A.; Tong, S.Y.C.; Zhang, Y.; Lamlertthon, S.; Sharma-Kuinkel, B.K.; Rude, T.; Ahn, S.H.; Ruffin, F.; Llorens, L.; Tamarana, G.; et al. Panton-Valentine leukocidin is not the primary determinant of outcome for Staphylococcus aureus skin infections: Evaluation from the CANVAS studies. PLoS ONE 2012, 7, e37212. [Google Scholar] [CrossRef]
- Chen, J.; Luo, Y.; Zhang, S.; Liang, Z.; Wang, Y.; Zhang, Y.; Zhou, G.; Jia, Y.; Chen, L.; She, D. Community-acquired necrotizing pneumonia caused by methicillin-resistant Staphylococcus aureus producing Panton–Valentine leukocidin in a Chinese teenager: Case report and literature review. Int. J. Infect. Dis. 2014, 26, 17–21. [Google Scholar] [CrossRef]
- Rajova, J.; Pantůček, R.; Petraš, P.; Varbanovova, I.; Mašlaňova, I.; Beneš, J. Necrotizing pneumonia due to clonally diverse Staphylococcus aureus strains producing Panton-Valentine leukocidin: The Czech experience. Epidemiol. Infect. 2016, 144, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Takigawa, Y.; Fujiwara, K.; Saito, T.; Nakasuka, T.; Ozeki, T.; Okawa, S.; Takada, K.; Iwamoto, Y.; Kayatani, H.; Minami, D.; et al. Rapidly progressive multiple cavity formation in necrotizing pneumonia caused by community-acquired methicillin-resistant Staphylococcus aureus positive for the Panton-Valentine leucocidin gene. Intern. Med. 2019, 58, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Naas, T.; Fortineau, N.; Spicq, C.; Robert, J.; Jarlier, V.; Nordmann, P. Three-year survey of community-acquired methicillin-resistant Staphylococcus aureus producing Panton-Valentine leukocidin in a French university hospital. J. Hosp. Infect. 2005, 61, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Hatfull, G.F.; Hendrix, R.W. Bacteriophages and their genomes. Curr. Opin. Virol. 2011, 1, 298–303. [Google Scholar] [CrossRef]
- Waldron, D.E.; Lindsay, J.A. Sau1: A novel lineage-specific type I restriction-modification system that blocks horizontal gene transfer into Staphylococcus aureus and between S. aureus isolates of different lineages. J. Bacteriol. 2006, 188, 5578–5585. [Google Scholar] [CrossRef]
- McCarthy, A.J.; Loeffler, A.; Witney, A.A.; Gould, K.A.; Lloyd, D.H.; Lindsay, J.A. Extensive horizontal gene transfer during Staphylococcus aureus co-colonization in vivo. Genome Biol. Evol. 2014, 6, 2697–2708. [Google Scholar] [CrossRef]
- O'Brien, F.G.; Coombs, G.W.; Pearman, J.W.; Gracey, M.; Moss, F.; Christiansen, K.J.; Grubb, W.B. Population dynamics of methicillin-susceptible and -resistant Staphylococcus aureus in remote communities. J. Antimicrob. Chemother. 2009, 64, 684–693. [Google Scholar] [CrossRef]
- Jones, T.F.; Kellum, M.E.; Porter, S.S.; Bell, M.; Schaffner, W. An outbreak of community-acquired foodborne illness caused by methicillin-resistant Staphylococcus aureus. Emerg. Infect. Dis. 2002, 8, 82–84. [Google Scholar] [CrossRef]
- European Food Safety Authority. Assessment of the Public Health significance of methicillin resistant Staphylococcus aureus (MRSA) in animals and foods. EFSA J. 2009, 99, 1–73. [Google Scholar] [CrossRef]
- Issa, G.; Aydin, A. Detection of methicillin resistant Staphylococcus aureus strains and typing of staphylococcal cassette chromosome mec from various foods originated in different region from Turkey. Kafkas Univ. Veter Fak. Derg. 2021, 27, 323–329. [Google Scholar] [CrossRef]
- Varshney, A.K.; Martinez, L.R.; Hamilton, S.M.; Bryant, A.E.; Levi, M.H.; Gialanella, P.; Stevens, D.L.; Fries, B.C. Augmented Production of Panton-Valentine Leukocidin Toxin in Methicillin-Resistant and Methicillin-Susceptible Staphylococcus aureus Is Associated with Worse Outcome in a Murine Skin Infection Model. J. Infect. Dis. 2010, 201, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Daskalaki, M.; Rojo, P.; Marin-Ferrer, M.; Barrios, M.; Otero, J.R.; Chaves, F. Panton-Valentine leukocidin-positive Staphylococcus aureus skin and soft tissue infections among children in an emergency department in Madrid, Spain. Clin. Microbiol. Infect. 2010, 16, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Issartel, B.; Tristan, A.; Lechevallier, S.; Bruyere, F.; Lina, G.; Garin, B.; Lacassin, F.; Bes, M.; Vandenesch, F.; Etienne, J. Frequent carriage of Panton-Valentine leucocidin genes by Staphylococcus aureus isolates from surgically drained abscesses. J. Clin. Microbiol. 2005, 43, 3203–3207. [Google Scholar] [CrossRef] [PubMed]
- Sudagidan, M.; Aydin, A. Virulence properties of methicillin-susceptible Staphylococcus aureus food isolates encoding Panton-Valentine Leukocidin gene. Int. J. Food Microbiol. 2010, 138, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Aydin, A.; Sudagidan, M.; Muratoglu, K. Prevalence of staphylococcal enterotoxins, toxin genes and genetic-relatedness of foodborne Staphylococcus aureus strains isolated in Marmara Region of Turkey. Int. J. Food Microbiol. 2011, 148, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Aydin, A.; Muratoglu, K.; Sudagidan, M.; Bostan, K.; Okuklu, B.; Harsa, S. Prevalence and antibiotic resistance of foodborne Staphylococcus aureus isolates in Turkey. Foodborne Pathog. Dis. 2011, 8, 63–69. [Google Scholar] [CrossRef]
- Holzinger, D.; Gieldon, L.; Mysore, V.; Nippe, N.; Taxman, D.J.; Duncan, J.A.; Broglie, P.M.; Marketon, K.; Austermann, J.; Vogl, T.; et al. Staphylococcus aureus Panton-Valentine leukocidin induces an inflammatory response in human phagocytes via the NLRP3 inflammasome. J. Leukoc. Biol. 2012, 92, 1069–1081. [Google Scholar] [CrossRef]
- Yoong, P.; Pier, G.B. Immune-Activating properties of Panton Valentine Leukocidin improve the outcome in a model of methicillin-resistant Staphylococcus aureus pneumonia. Infect. Immun. 2012, 8, 2894–2904. [Google Scholar] [CrossRef]
- Ma, X.; Chang, W.; Zhang, C.; Zhou, X.; Yu, F. Staphylococcal Panton-Valentine Leukocidin Induces Pro-Inflammatory Cytokine Production and Nuclear Factor-Kappa B Activation in Neutrophils. PLoS ONE 2012, 7, e34970. [Google Scholar] [CrossRef]
- Atlas, R.M. Handbook of Microbiological Media, 4th ed.; ASM Press& CRC Press: Washington, DC, USA, 2010; p. 332. [Google Scholar]
- Diep, B.A.; Chan, L.; Tattevin, P.; Kajikawa, O.; Martin, T.R.; Basuino, L.; Mai, T.T.; Marbach, H.; Braughton, K.R.; Whitney, A.R.; et al. Polymorphonuclear leukocytes mediate Staphylococcus aureus Panton–Valentine leukocidin-induced lung inflammation and injury. Proc. Natl. Acad. Sci. USA 2010, 107, 5587–5592. [Google Scholar] [CrossRef]
- Klein, M.; Vignaud, J.M.; Hennequin, V.; Toussaint, B.; Bresler, L.; Plenat, F.; Ledere, J.; Duprez, A.; Weryha, G. Increased expression of the vascular endothelial growth factor is a pejorative prognosis marker in papillary thyroid carcinoma. J. Clin. Endocrinol. Metab. 2001, 86, 656–658. [Google Scholar] [CrossRef] [PubMed]
- Sudagidan, M.; Cavusoglu, C.; Bacakoglu, F. Investigation of the virulence genes in methicillin-resistant Staphylococcus aureus strains isolated from biomaterial surfaces. Mikrobiyoloji Bul. 2008, 42, 29–39. [Google Scholar]
- Sudagidan, M.; Aydin, A. Screening virulence properties of staphylococci isolated from meat and meat products. Vet. Med. Austria/Wien. Tierarztl. Mon. 2008, 96, 128–134. [Google Scholar]
- Hookey, J.V.; Richardson, J.F.; Cookson, B.D. Molecular typing of Staphylococcus aureus based on PCR restriction fragment length polymorphism and DNA sequence analysis of the coagulase gene. J. Clin. Microbiol. 1998, 36, 1083–1089. [Google Scholar] [CrossRef]
- Aires-de-Sousa, M.; Boye, K.; de Lencastre, H.; Deplano, A.; Enright, M.C.; Etienne, J.; Friedrich, A.; Harmsen, D.; Holmes, A.; Huijsden, X.W.; et al. High interlaboratory reproducibility of DNA sequence-based typing of bacteria in a multicenter study. J. Clin. Microbiol. 2006, 44, 619–621. [Google Scholar] [CrossRef]
- Lina, G.; Piemont, Y.; Godail-Gamot, F.; Bes, M.; Peter, M.O.; Gauduchon, V.; Vandenesh, F.; Etienne, J. Involvement of Panton–Valentine leukocidin-producing Staphylococcus aureus in primary skin infections and pneumonia. Clin. Infect. Dis. 1999, 29, 1128–1132. [Google Scholar] [CrossRef]
- Durmaz, R.; Otlu, B.; Çaliskan, A.; Gursoy, N. A Rapid pulsed-field gel electrophoresis (PFGE) protocol developed for subtyping Acinetobacter baumannii, Escherichia coli, and Klebsiella spp. ANKEM Derg. 2007, 21, 113–117. [Google Scholar]
- Tomita, T.; Kamio, Y. Molecular biology of the pore-forming cytolysins from Staphylococcus aureus, α-and γ-hemolysins and leucocidin. Biosci. Biotechnol. Biochem. 1997, 61, 565–572. [Google Scholar] [CrossRef]
- Löffler, B.; Niemann, S.; Ehrhardt, C.; Horn, D.; Lanckohr, C.; Lina, G.; Ludwig, S.; Peters, G. Pathogenesis of Staphylococcus aureus necrotizing pneumonia: The role of PVL and an influenza coinfection. Expert Rev. Anti Infect. Ther. 2013, 11, 1041–1051. [Google Scholar] [CrossRef]
- Gillet, Y.; Issartel, B.; Vanhems, P.; Fournet, J.C.; Lina, G.; Bes, M.; Vandenesch, F.; Piémont, Y.; Brousse, N.; Floret, D.; et al. Association between Staphylococcus aureus strains carrying gene for Panton-Valentine leukocidin and highly lethal necrotizing pneumonia in young immunocompetent patients. Lancet 2002, 359, 753–759. [Google Scholar] [CrossRef]
- Rao, G.G.; Batura, R.; Nicholl, R.; Coogan, F.; Patel, B.; Bassett, P.; Kearns, A.M. Outbreak report of investigation and control of an outbreak of Panton-Valentine Leukocidin-positive methicillin-sensitive Staphylococcus aureus (PVL-MSSA) infection in neonates and mothers. BMC Infect. Dis. 2019, 19, 178. [Google Scholar] [CrossRef]
- Rhee, C.H.; Woo, G.J. Emergence and characterization of foodborne methicillin-resistant Staphylococcus aureus in Korea. J. Food Prot. 2010, 73, 2285–2290. [Google Scholar] [CrossRef] [PubMed]
- Witte, W.; Strommenger, B.; Stanek, C.; Cuny, C. Methicillin-resistant Staphylococcus aureus ST398 in humans and animals in central Europe. Emerg. Infect. Dis. 2007, 13, 255–258. [Google Scholar] [CrossRef] [PubMed]
- Clarridge, J.E., III; Harrington, A.T.; Roberts, M.C.; Soge, O.O.; Maquelin, K. Impact of strain typing methods on assessment of relationship between paired nares and wound isolates of methicillin-resistant Staphylococcus aureus. J. Clin. Microbiol. 2013, 51, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Eells, S.J.; David, M.Z.; Taylor, A.; Ortiz, N.; Kumar, N.; Sieth, J.; Boyle-Vavra, S.; Daum, R.S.; Miller, L.G. Persistent environmental contamination with USA300 methicillin-resistant Staphylococcus aureus and other pathogenic strain types in households with S. aureus skin infections. Infect. Control. Hosp. Epidemiol. 2014, 35, 1373–1382. [Google Scholar] [CrossRef]
- Desai, R.; Pannaraj, P.S.; Agopian, J.; Sugar, C.A.; Liu, G.Y.; Miller, L.G. Survival and transmission of community-associated methicillin-resistant Staphylococcus aureus from fomites. Am. J. Infect. Control 2011, 39, 219–225. [Google Scholar] [CrossRef]
- Argudín, M.Á.; Mendoza, M.C.; Rodicio, M.R. Food poisoning and Staphylococcus aureus enterotoxins. Toxins 2010, 2, 1751–1773. [Google Scholar] [CrossRef]
- Lipinska, U.; Hermans, K.; Meulemans, L.; Dumitrescu, O.; Badiou, C.; Duchateau, L.; Haesebrouck, F.; Etienne, J.; Lina, G. Panton-Valentine leukocidin does play a role in the early stage of Staphylococcus aureus skin infections: A rabbit model. PLoS ONE 2011, 6, e22864. [Google Scholar] [CrossRef]
- Montgomery, C.P.; Daum, R.S. Transcription of Inflammatory Genes in the Lung after Infection with Community-Associated Methicillin-Resistant Staphylococcus aureus: A Role for Panton-Valentine Leukocidin. Infect. Immun. 2009, 77, 2159–2167. [Google Scholar] [CrossRef]
- Labandeira-Rey, M.; Couzon, F.; Boisset, S.; Brown, E.L.; Bes, M.; Benito, Y.; Barbu, E.M.; Vazquez, V.; Höök, M.; Etienne, J.; et al. Staphylococcus aureus Panton-Valentine Leukocidin Causes Necrotizing Pneumonia. Science 2007, 315, 1130–1133. [Google Scholar] [CrossRef]
- Diep, B.A.; Palazzolo-Balance, A.M.; Tattevin, P.; Basuino, L.; Braughton, K.R.; Whitney, A.R.; Chen, L.; Kreiswirth, B.N.; Otto, M.; DeLeo, F.R.; et al. Contribution of Panton-Valentine leukocidin in community-associated methicillin-resistant Staphylococcus aureus pathogenesis. PLoS ONE 2008, 3, e3198. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.L.; Dumitrescu, O.; Thomas, D.; Badiou, C.; Koers, E.M.; Choudhury, P.; Vasquez, V.; Etienne, J.; Lina, G.; Vandenesch, F.; et al. The Panton–Valentine leukocidin vaccine protects mice against lung and skin infections caused by Staphylococcus aureus USA300. Clin. Microbiol. Infect. 2009, 15, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Löffler, B.; Hussain, M.; Grundmeier, M.; Brück, M.; Holzinger, D.; Varga, G.; Roth, J.; Kahl, B.C.; Proctor, R.A.; Peters, G. Staphylococcus aureus Panton-Valentine Leukocidin Is a Very Potent Cytotoxic Factor for Human Neutrophils. PloS Pathog. 2010, 6, e1000715. [Google Scholar] [CrossRef]
- Tseng, C.W.; Kyme, P.; Low, J.; Rocha, M.A.; Alsabeh, R.; Miller, L.G.; Otto, M.; Arditi, M.; Diep, B.A.; Nizet, V.; et al. Staphylococcus aureus Panton-Valentine leukocidin contributes to inflammation and muscle tissue injury. PLoS ONE 2009, 4, e6387. [Google Scholar] [CrossRef]
- Stulik, L.; Rouha, H.; Labrousse, D.; Visram, Z.C.; Badarau, A.; Maierhofer, B.; Groß, K.; Weber, S.; Kramarić, M.D.; Glojnarić, I.; et al. Preventing lung pathology and mortality in rabbit Staphylococcus aureus pneumonia models with cytotoxin-neutralizing monoclonal IgGs penetrating the epithelial lining fluid. Sci. Rep. 2019, 9, 5339. [Google Scholar] [CrossRef]
- Montgomery, C.P.; Boyle-Vavra, S.; Adem, P.V.; Lee, J.C.; Husain, A.N.; Clasen, J.; Daum, R.S. Comparison of virulence in community-associated methicillin-resistant Staphylococcus aureus pulsotypes USA300 and USA400 in a rat model of pneumonia. J. Infect. Dis. 2008, 198, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Zhang, W.; Huang, J.; Liu, H.; Zhang, T. Effect of recombinant Panton Valentine Leukocidin in vitro on apoptosis and cytokine production of human alveolar macrophages. Can. J. Microbiol. 2010, 56, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Laventie, B.-J.; Rademaker, H.J.; Saleh, M.; de Boer, E.; Janssens, R.; Bourcier, T.; Subilia, A.; Marcellin, L.; van Haperen, R.; Lebbink, J.H.G.; et al. Heavy chain-only antibodies and tetravalent bispecific antibody neutralizing Staphylococcus aureus leukotoxins. Proc. Natl. Acad. Sci. USA 2011, 108, 16404–16409. [Google Scholar] [CrossRef] [PubMed]
- Goodman, R.B.; Pugin, J.; Lee, J.S.; Matthay, M.A. Cytokine-mediated inflammation in acute lung injury. Cytokine Growth Factor Rev. 2003, 14, 523–535. [Google Scholar] [CrossRef]
- König, B.; Prévost, G.; Piemont, Y.; König, W. Effects of Staphylococcus aureus leukocidins on inflammatory mediator release from human granulocytes. J. Infect. Dis. 1995, 171, 607–613. [Google Scholar] [CrossRef]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in inflammatory disease. Int. J. Mol. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Group-I | Group-II | Group-III | Group-IV | Group-V | p Value |
|---|---|---|---|---|---|---|
| Oedema | 0 (0–0) | 0 (0–2) | 2 (0–3) | 2 (0–3) | 1 (0–3) | 0.007 |
| Haemorrhage | 0 (0–0) | 0 (0–0) | 2 (0–3) | 1.5 (0–3) | 1 (0–3) | <0.001 |
| Exudate/Fibrin | 0 (0–0) | 0 (0–0) | 1 (0–2) | 1 (0–3) | 0.5 (0–2) | 0.003 |
| PMN infiltration | 0 (0–0) | 0 (0–1) | 2 (0–3) | 2 (0–3) | 1 (0–3) | <0.001 |
| Necrosis | 0 (0–0) | 0 (0–0) | 0.5 (0–3) | 1 (0–3) | 0.5 (0–3) | 0.016 |
| Parameters | Group I/Group III (NC/HT480) | Group I/Group IV (NC/M1) | Group I/Group V (NC/YF1B-b) | Group II/Group III (VC/HT480) | Group II/Group IV (VC/M1) | Group II/Group V (VC/YF1B-b) |
|---|---|---|---|---|---|---|
| Oedema | 0.004 | 0.004 | 0.004 | 0.071 | 0.043 | 0.183 (NS) |
| Haemorrhage | 0.001 | 0.004 | 0.003 | 0.001 | 0.004 | 0.003 |
| Exudate/Fibrin | 0.010 | 0.003 | 0.027 | 0.010 | 0.003 | 0.027 |
| PMN infiltration | 0.001 | 0.001 | 0.003 | 0.006 | 0.006 | 0.085 (NS) |
| Necrosis | 0.027 | 0.010 | 0.027 | 0.027 | 0.010 | 0.027 |
| Cytokine | Group-I | Group-II | Group-III | Group-IV | Group-V | p Value |
|---|---|---|---|---|---|---|
| IL-6 | 0 (0–1) | 1 (1–2) | 2 (1–3) | 2 (1–3) | 1 (1–2) | 0.001 |
| IL-8 | 0.5 (0–1) | 1.5 (1–2) | 2 (1–3) | 1.5 (1–2) | 2 (1–3) | 0.005 |
| IL-10 | 0 (0–1) | 1 (1–2) | 1.5 (1–3) | 2 (1–3) | 1.5 (1–2) | <0.001 |
| TNF-α | 0 (0–1) | 1 (1–2) | 2 (1–3) | 2 (1–3) | 1.5 (1–2) | 0.001 |
| Cytokine | Group I/Group II (NC/VC) | Group I/Group III (NC/HT480) | Group I/Group IV (NC/M1) | Group I/Group V (NC/YF1B-b) |
|---|---|---|---|---|
| IL-6 | 0.002 | 0.001 | 0.001 | 0.003 |
| IL-8 | 0.010 | 0.004 | 0.006 | 0.004 |
| IL-10 | 0.001 | 0.001 | <0.001 | 0.001 |
| TNF-α | 0.005 | 0.001 | 0.002 | 0.003 |
| Group I (NC) | Group II (VC) | Group III (HT480) | Group IV (M1) | Group V (YF1B-b) | p Values in Repeated ANOVA Statistics | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Cytokine | Time Points | Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | p-Values in One-Way ANOVA | Group | Measurement Time | Group × Measurement Time |
| IL-6 | 1 (3 h) | 10.30 ± 0.63 | 10.88 ± 1.52 | 17.24 ± 2.04 | 12.83 ± 1.58 | 15.41 ± 1.65 | 0.014 | 0.004 | 0.025 | 0.233 |
| 2 (6 h) | 10.94 ± 0.77 | 12.27 ± 0.87 | 25.69 ± 6.44 | 18.44 ± 2.29 | 15.25 ± 1.25 | 0.017 | ||||
| Significance | 0.108 | 0.420 | 0.162 | 0.071 | 0.955 | |||||
| IL-8 | 1 (3 h) | 11.83 ± 2.22 | 23.22 ± 13.24 | 72.24 ± 3.81 | 69.14 ± 4.71 | 74.67 ± 4.39 | <0.001 | <0.001 | 0.297 | 0.085 |
| 2 (6 h) | 11.76 ± 2.37 | 26.85 ± 3.26 | 73.90 ± 3.72 | 66.59 ± 2.54 | 61.72 ± 3.03 | <0.001 | ||||
| Significance | 0.808 | 0.205 | 0.637 | 0.686 | 0.079 | |||||
| IL-10 | 1 (3 h) | 2.69 ± 0.48 | 17.18 ± 3.36 | 41.43 ± 5.95 | 38.77 ± 8.66 | 41.27 ± 6.42 | <0.001 | <0.001 | 0.411 | 0.970 |
| 2 (6 h) | 2.49 ± 0.44 | 17.81 ± 3.95 | 45.71 ± 8.64 | 46.45 ± 5.43 | 45.01 ± 8.44 | <0.001 | ||||
| Significance | 0.100 | 0.739 | 0.656 | 0.487 | 0.787 | |||||
| TNF-α | 1 (3 h) | 6.22 ± 1.09 | 27.61 ± 5.32 | 86.81 ± 10.31 | 109.63 ± 18.17 | 122.77 ± 36.67 | <0.001 | <0.001 | 0.004 | 0.033 |
| 2 (6 h) | 6.45 ± 1.02 | 31.91 ± 5.49 | 178.55 ± 29.66 | 143.08 ± 14.72 | 141.22 ± 10.98 | <0.001 | ||||
| Significance | 0.558 | 0.193 | 0.011 | 0.010 | 0.650 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yildirim, F.; Sudagidan, M.; Aydin, A.; Akyazi, I.; Bayrakal, G.M.; Yavuz, O.; Gurel, A. In Vivo Pathogenicity of Methicillin-Susceptible Staphylococcus aureus Strains Carrying Panton–Valentine Leukocidin Gene. Life 2022, 12, 2126. https://doi.org/10.3390/life12122126
Yildirim F, Sudagidan M, Aydin A, Akyazi I, Bayrakal GM, Yavuz O, Gurel A. In Vivo Pathogenicity of Methicillin-Susceptible Staphylococcus aureus Strains Carrying Panton–Valentine Leukocidin Gene. Life. 2022; 12(12):2126. https://doi.org/10.3390/life12122126
Chicago/Turabian StyleYildirim, Funda, Mert Sudagidan, Ali Aydin, Ibrahim Akyazi, Gulay Merve Bayrakal, Orhan Yavuz, and Aydin Gurel. 2022. "In Vivo Pathogenicity of Methicillin-Susceptible Staphylococcus aureus Strains Carrying Panton–Valentine Leukocidin Gene" Life 12, no. 12: 2126. https://doi.org/10.3390/life12122126
APA StyleYildirim, F., Sudagidan, M., Aydin, A., Akyazi, I., Bayrakal, G. M., Yavuz, O., & Gurel, A. (2022). In Vivo Pathogenicity of Methicillin-Susceptible Staphylococcus aureus Strains Carrying Panton–Valentine Leukocidin Gene. Life, 12(12), 2126. https://doi.org/10.3390/life12122126