1. Introduction
Brain disease, including brain cancers and central nervous system (CNS) disorders, are among the most prevalent, devastating and inadequately treated conditions. Developing safe and effective therapeutic strategies for CNS malignancies presents significant challenges, including high failure rates and extended timelines to reach the market compared to non-CNS therapies. Notably, the U.S Food and Drug Administration (FDA) has approved several drugs for utilization for the treatment of brain tumors, such as everolimus, bevacizumab, carmustine, dabrafenib mesylate, temozolomide, trametinib dimethyl sulfoxide, and vorasidenib citrate [
1]. A critical obstacle in the development of CNS therapies is the blood–brain barrier (BBB), initially identified by Paul Ehrlich in the late 19th and early 20th centuries. Ehrlich’s observation that dyes injected into the bloodstream failed to stain brain tissue underscored the barrier’s selective permeability [
2].
The BBB is a highly specialized and semipermeable structure that separates the CNS from the systemic circulation. It plays a crucial role in maintaining the brain microenvironment. The BBB acts as both a protective shield and a regulator, ensuring the CNS is isolated from toxins, pathogens, and potentially harmful fluctuations in blood composition, such as hormones or metabolites that could disrupt normal function [
3]. By maintaining a tightly controlled environment, the BBB is essential for preserving neuronal activity and overall brain homeostasis.
Structurally, the BBB is composed of tightly bound endothelial cells (ECs) that line the brain’s capillaries. These ECs are reinforced by surrounding pericytes, astrocytes, and other supporting elements in the extracellular matrix (ECM) [
4]. Pericytes and astrocytes not only provide structural support, but also contribute to the regulation of BBB permeability and respond to physiological or pathological changes in the brain. They are crucial for the development of accurate in vitro models of the BBB [
5]. Other cellular and molecular components, such as enzymes and efflux transporters like P-glycoprotein (P-gp) and breast cancer resistance protein (BCRP), further enhance the barrier’s protective function by actively removing harmful substances [
6]. Efflux transporters are proteins embedded in cell membranes that actively transport molecules out of the cells, essentially acting like “pumps” to remove unwanted substances. The BBB’s complex architecture and stringent selective permeability significantly hinder drug delivery to the CNS, excluding over 95% of drug candidates. The BBB allows the passive diffusion of only lipophilic molecules with a molecular weight under 400 Da and specific drugs with favorable physicochemical properties. Additionally, active transport systems facilitate the controlled entry of essential nutrients, such as glucose, amino acids, and ions required for neural metabolism [
7,
8]. However, large molecules, hydrophilic compounds, and many therapeutic agents, such as antibody–drug conjugate (ADC), are excluded, limiting the ability of systemic treatment to reach the brain. This exclusion becomes even more challenging in pathological conditions like CNS tumors. The progression of disease leads to the formation of a blood–tumor barrier (BTB), which imposes additional constraints on achieving effective drug concentrations within the tumor microenvironment [
9].
In recent years, drug development for brain diseases has advanced substantially, with a particular focus on the utilization of polymeric and lipid-based nanoparticles (NPs) to enhance drug delivery to the brain. Nanocarriers can be modified to optimize brain targeting, enhance stability, and modify drug-release patterns. Despite these efforts, the success rate of CNS drug development remains among the lowest in the pharmaceutical industry, primarily due to the challenges posed by the BBB and BTB [
7,
8]. Effective and efficient drug delivery systems capable of traversing this protective barrier are indispensable for the development of therapies targeting CNS disorders. These systems must not only transport therapeutic agents across the BBB, but also ensure targeted delivery to specific brain regions, minimize systemic toxicity, and optimize pharmacokinetics and pharmacodynamics [
10]. Lipid-based NPs, such as liposomes, lipid nanoparticles (LNPs), nanostructured lipid carriers (NLCs), solid lipid nanoparticles (SLNs), and emulsions, have emerged as promising carriers for drug delivery [
11]. Their structural properties, including a lipid bilayer or lipid matrix, mimic biological membranes, facilitating the crossing of the BBB. Lipid-based NPs can encapsulate a wide range of drugs, including small molecules, peptides, and nucleic acids, protecting them from degradation and enabling sustained release, making them more attractive [
12]. However, the potential immune reactions to NPs must be carefully considered and managed [
13]. Polymeric NPs, including micelles, polyplexes, and polymeric hydrogels, offer additional advantages, such as a high drug-loading capacity, tunable release profiles, and structural versatility [
14]. These NPs have been successfully used in preclinical and clinical research, delivering drugs for CNS diseases including medulloblastoma (MB), Alzheimer’s disease (AD), Parkinson’s disease, multiple sclerosis, and stroke [
15]. One of the most notable advantages of NP-based systems is their ability to encapsulate drugs into nanocarriers, thereby facilitating sustained drug release. This capability not only reduces the dosing frequency, but also enhances patient compliance and optimizes therapeutic outcomes by ensuring that drug concentrations remain within the therapeutic window [
16,
17].
Surface modifications of NPs further enhance their functionality and ability to cross the BBB. Functionalization with targeting ligands, such as transferrin, lactoferrin, insulin, or low-density lipoprotein (LDL) receptors, facilitates receptor-mediated transcytosis (RMT) across the BBB [
18]. Additionally, modifying NPs with cell-penetrating peptides or specific antibodies can improve their interaction with BBB ECs and promote selective delivery to the brain [
19]. Transporter-mediated transcytosis (TMT), another promising strategy, exploits endogenous transport systems, such as glucose or amino acid transporters, to deliver drugs effectively into the brain [
20]. TMT routes across the BBB include multiple mechanisms, each with unique challenges and opportunities for therapeutic delivery. For the BBB, the paracellular route allows water-soluble drugs to pass through tight junctions between ECs, though this pathway is highly restricted due to the barrier’s tightness [
4]. The transcellular lipophilic pathway enables small, lipophilic molecules to diffuse across ECs, but limits the range of molecules that can be delivered [
21]. Advanced strategies focus on exploiting active transport mechanisms such as RMT and TMT. RMT uses ligand–receptor interactions to transport macromolecules such as NPs, proteins, or peptides across the BBB, while TMT leverages transport proteins to ferry small molecules and nutrients [
22]. Furthermore, adsorption-mediated transcytosis (AMT), driven by electrostatic interactions between positively charged NPs and negatively charged cell membranes, offers an alternative route for delivering therapeutics to the brain. However, AMT remains less specific than RMT or TMT, which limits its utility [
23].
Ongoing research into the anatomical and pathological characteristics of the BBB is crucial for enhancing these strategies. Advancements in understanding how BBB integrity alters under pathological conditions provide insights into developing targeted therapies. Current research focuses on developing NPs with ligands or other modifications that can specifically bind to receptors on the BBB or/and the targeted cells to facilitate CNS drug delivery. While promising in preclinical studies, the translation of NP-based CNS drug delivery to clinical trials can be challenging due to safety concerns and complex formulation considerations. In this review, the structure and physiology of the BBB will be summarized, accompanied by an in-depth discussion of various RMT and TMT mechanisms for crossing the BBB. Particular attention will be given to the emerging roles of RMT and TMT as innovative strategies for overcoming the BBB, as well as recent advancements in drug delivery over the past five years in the treatment of CNS disorders. Through such focused research, novel approaches for crossing the BBB and achieving effective brain accumulation of therapeutics can be realized, ultimately addressing the unmet medical needs of CNS diseases.
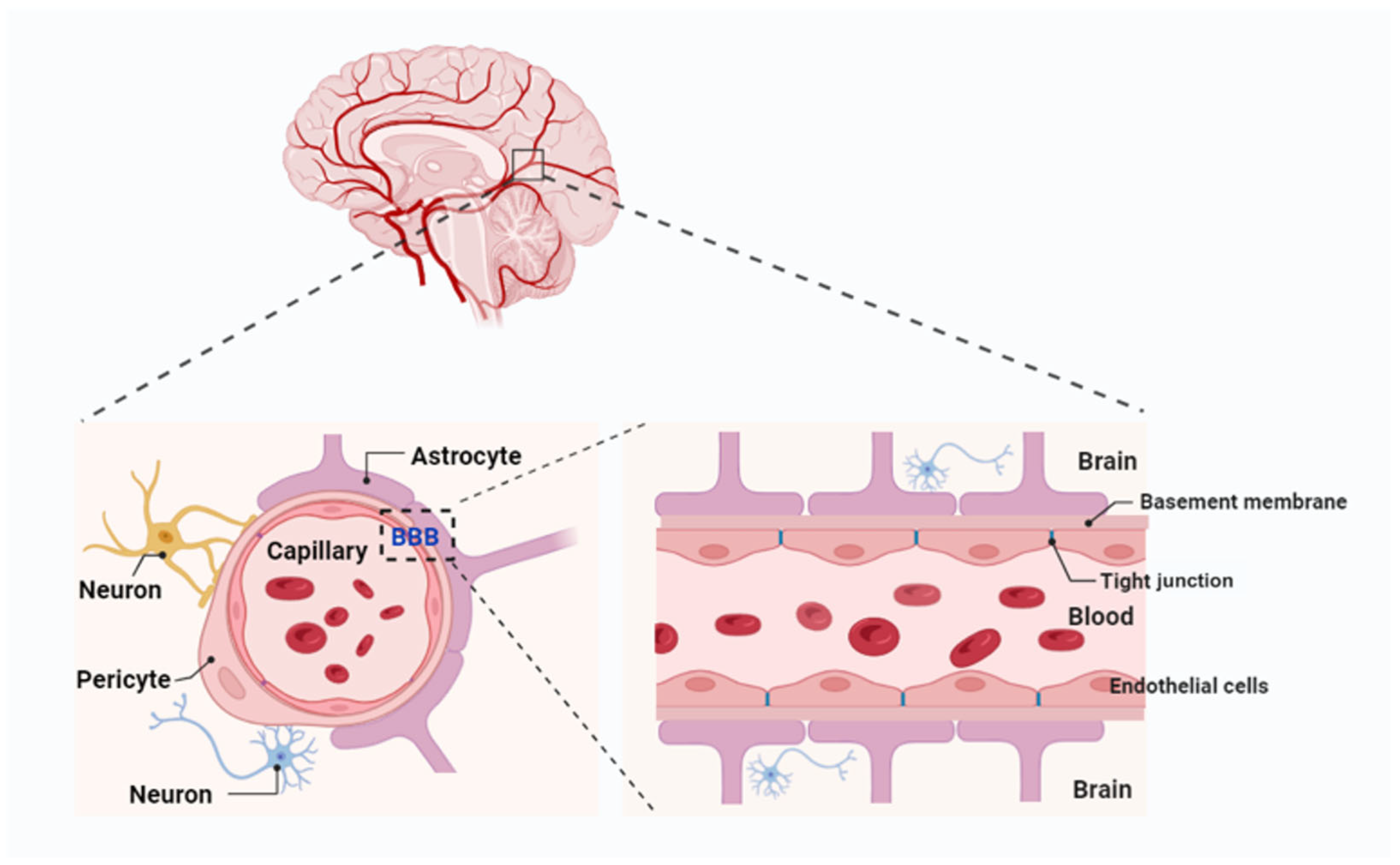
2. BBB Structure and Physiology
The BBB is a complex and dynamic structure, primarily composed of ECs with tight junctions, supported by a basement membrane, astrocytes, pericytes, and extracellular matrix (ECM) components. Its major functional site is located in the capillaries (
Figure 1) [
4]. The BBB is considered the largest interface for blood–brain exchange, as it has a combined surface area of 12 and 18 m
2 per average adult, based on an average microvessel surface area of 150 and 200 cm
2 per gram of tissue [
24]. Additionally, the blood–cerebrospinal fluid barrier (BCSFB) [
25] and the arachnoid barrier represent the second and third barriers in the brain. However, the arachnoid barrier’s contribution to the exchange between the blood and the brain is insignificant, due to its avascular nature and relatively small surface area compared to the BBB and BCSFB [
26].
The function of the BBB is contingent upon the coordinated contributions of multiple cell types within the neurovascular unit (NVU). This unit comprises astrocytes, pericytes, neurons, and microglia, all of which work together to maintain the integrity and selective permeability of the BBB [
27]. Astrocytes, with their extensive “endfeet” covering 99% of the basal capillary membrane, play a vital role in the development and maintenance of the BBB by secreting factors such as GDNF, angiopoietin-1, and angiotensin II. These factors support EC integrity and regulate barrier function. Pericytes, which encircle ECs, provide structural support, regulate blood flow, and limit barrier permeability. Recent research highlights the location-dependent diversity of pericytes: those adjacent to arterioles regulate blood flow through contractile properties, while venous capillary pericytes maintain barrier integrity and modulate immune responses. These interactions between pericytes, astrocytes, and ECs are critical for preserving BBB integrity. Studies have demonstrated that pericyte deficiency increases BBB permeability, contributing to neurovascular dysfunction in diseases such as Alzheimer’s, stroke, and multiple sclerosis [
28]. Consequently, these cell types form a highly coordinated system that safeguards the brain while facilitating the regulated exchange of nutrients and waste products.
The ECs that form the BBB are tightly joined by specialized junctions known as tight junctions (TJs), composed of transmembrane proteins such as claudins, occludins, and junctional adhesion molecules (JAMs) [
29]. These proteins create a high-resistance paracellular barrier that limits the crossing of ions and molecules while ensuring selective permeability to essential substances such as oxygen and glucose. Claudins, particularly claudin-5, are crucial for maintaining BBB integrity; their disruption has been linked to various neurological disorders [
30]. Additionally, ECs exhibit limited pinocytotic activity and express efflux transporters [
31]. Efflux transporters in brain capillary ECs further enhance the BBB’s protective function by actively removing undesirable substances and metabolic byproducts from the brain into the systemic circulation. Key players include multidrug resistance transporters like P-glycoprotein, multidrug resistance proteins (MRPs), and BCRP [
32]. While these systems safeguard the CNS, they also hinder the delivery of many therapeutic agents. Drugs designed to treat neurological disorders are often recognized as substrates and removed from the brain before achieving therapeutic levels. To address this, researchers are exploring strategies such as efflux transporter inhibitors, drug designs that evade transporter recognition, and advanced delivery systems. These approaches aim to balance the dual role of efflux transporters—protecting the brain while enabling effective drug delivery—without compromising the integrity or function of the BBB.
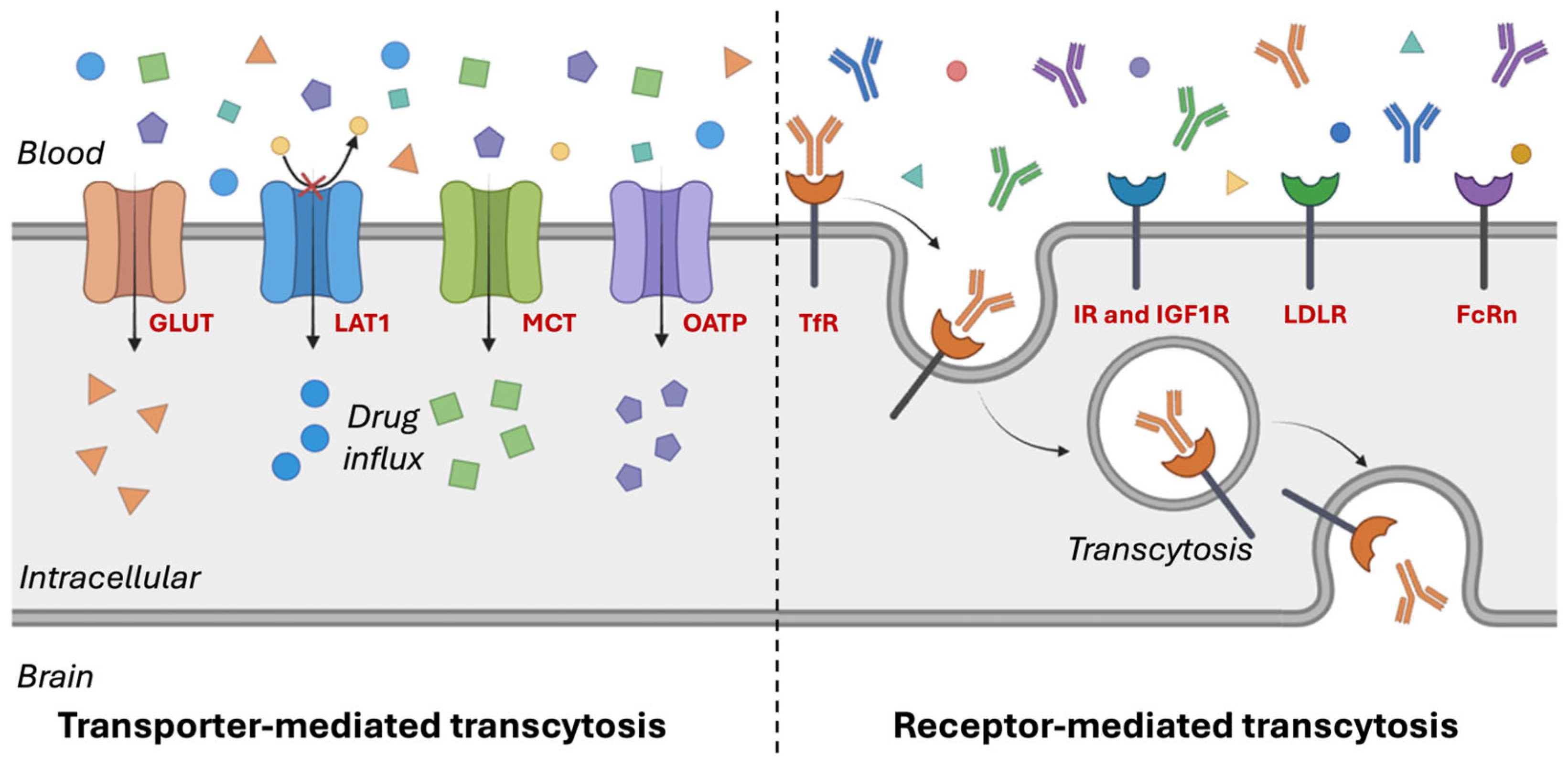
4. Mechanisms of BBB Crossing by Nanomaterials
Nanomaterials employ various mechanisms to cross the BBB, facilitating the targeted delivery of therapeutics to the brain. These mechanisms can be broadly categorized into passive and active transport pathways. Passive diffusion occurs when small, lipophilic nanomaterials cross the BBB without the involvement of specific transporters. In contrast, active transport mechanisms encompass TMT or carrier-mediated transport (CMT), RMT, and AMT. TMT exploits endogenous transporters expressed on brain ECs to actively shuttle molecules across the BBB, leveraging physiological pathways responsible for the transport of essential nutrients, peptides, and ions between the bloodstream and brain [
3]. RMT relies on ligands, peptides, or antibodies conjugated to nanomaterials that engage specific receptors, such as transferrin receptors (TfRs) or low-density lipoprotein receptor-related proteins (LRP1 and LRP2), to facilitate endocytosis and transcytosis across the BBB [
34]. AMT is mediated by electrostatic interactions between positively charged nanomaterials with negatively charged membrane components, promoting cellular uptake [
12]. Beyond these traditional approaches, emerging strategies, such as cell-mediated transport via monocytes or exosomes, and stimuli-responsive nanomaterials that react to pH, temperature, or enzymatic activity, are being explored to enhance BBB permeability and improve therapeutic efficacy. These mechanisms can be further classified into invasive or non-invasive approaches. Invasive approaches involve the temporary disruption of the BBB through physical means, enabling nanomaterials to cross the BBB via the paracellular pathway. Techniques such as focused ultrasound-mediated BBB opening and local delivery strategies fall under this category, which is also referred to as the paracellular pathway [
35,
36]. Conversely, non-invasive strategies maintain BBB integrity during drug delivery and rely on the transcellular pathway to improve nanomaterial transport. TMT, RMT, and AMT, as well as emerging cell-mediated and stimuli-responsive nanomaterials, fall within this category, and can be collectively to referred to as transcellular mechanisms. These diverse transport mechanisms offer promising strategies for overcoming the restrictive nature of the BBB, thereby offering novel opportunities for the treatment of neurological disorders.
5. Transporter-Mediated Transcytosis (TMT) for Crossing the BBB
Among the various strategies to overcome the BBB, TMT has gained significant attention due to its potential to facilitate efficient drug delivery to the CNS. TMT exploits specific endogenous transporters expressed on the ECs of the BBB to actively transport molecules across this tightly regulated barrier. By leveraging the natural mechanisms that transport essential nutrients, peptides, and ions between the bloodstream and brain, TMT enables efficient and targeted drug delivery to the brain (
Figure 2) [
37,
38]. These TMT transporters are encoded by genes within the solute carrier (SLC) transporter gene family, which comprises over 300 genes that regulate the translocation of various substrates across the BBB [
39]. Key transporters involved in TMT include glucose transporters (GLUTs), which mediate glucose and mannose uptake; neutral amino acid transporters (LAT1 and LAT2), responsible for transporting large neutral amino acids and certain drugs like L-dopa; and anionic/cationic amino acid transporters, which facilitate the transport of aspartate, glutamate, L-lysine, L-arginine, and L-ornithine [
38]. Additionally, monocarboxylate transporters (MCTs) play a critical role in lactate transport, while lipid transporters, such as LRP1, facilitate the translocation of lipoproteins and other macromolecules. Dipeptide transporters such as PEPT2 also contribute to TMT by shuttling di- and tripeptides across the BBB [
40,
41,
42].
The polarized distribution of transporters across the BBB contributes to its functional asymmetry. Certain transport proteins are exclusively localized to either the luminal or abluminal membrane, while others are expressed on both membranes of the ECs [
43]. By harnessing these transport pathways, targeted drug delivery strategies can be developed to facilitate the efficient transport of therapeutic agents that would otherwise be excluded from the brain. Recent advancements in TMT-based drug delivery have focused on two primary approaches: (1) designing drug molecules that structurally mimic endogenous substrates of specific transporters, and (2) conjugating therapeutic compounds or nanocarriers with ligands that selectively bind to and engage transporter-mediated pathways [
44]. TMT-based drug delivery systems have demonstrated significant potential in enhancing brain uptake of poorly permeable therapeutics, particularly through strategies such as LAT1-targeted prodrugs and GLUT1-based delivery platforms [
45,
46]. These innovative strategies improve the selective passage of therapeutics across the restrictive BBB, while preserving its structural integrity.
5.1. Glucose Transporters (GLUTs)
GLUTs, encoded by SLC2A1, play a crucial role in facilitating glucose transport across the BBB to meet the brain’s high energy demands [
47,
48]. Among the 14 identified human GLUT isoforms, GLUT1 to GLUT5 are the most extensively characterized, primarily transporting hexoses, notably D-glucose, while also facilitating the uptake of other hexoses, such as D-mannose and D-galactose, albeit with lower efficiency [
48,
49,
50,
51]. As a facilitative transporter, GLUT1 mediates passive glucose diffusion and is highly expressed on both the luminal (blood-facing) and abluminal (brain-facing) membranes of BBB ECs, ensuring a continuous glucose supply to the brain [
52]. Unlike sodium-dependent glucose transporters (SGLTs), which require ion co-transport, GLUT1 functions independently of sodium gradients, allowing for energy-independent glucose uptake [
53]. Beyond its fundamental physiological role, GLUT1 expression is tightly regulated in response to metabolic demands, and can be altered under pathological conditions [
54]. In neurodegenerative disorders such as AD, decreased GLUT1 expression impairs glucose transport to the brain, contributing to cognitive decline [
55]. Conversely, in glioblastoma and other malignancies, GLUT1 and GLUT3 cause rapid tumor proliferation by facilitating enhanced glucose uptake [
55,
56,
57,
58]. Furthermore, cerebrovascular conditions like ischemia can alter GLUT expression, thereby affecting glucose supply to the brain and further highlighting GLUTs as a potential therapeutic target [
58,
59].
Ligand-modified nanomaterials that mimic glucose structures or bind to GLUT1-binding domains have been developed to exploit GLUT1-mediated transport, enhancing CNS drug delivery. This understanding has led to an increasing interest in targeting GLUT1 for disease intervention and drug delivery. Recent studies have demonstrated that GLUT1-targeted NPs can improve the delivery of chemotherapeutic agents, neuroprotective peptides, and genetic material to the brain, leading to improved therapeutic efficacy [
60]. By harnessing GLUT1-mediated transport, researchers seek to enhance drug bioavailability and optimize treatment strategies for brain-related diseases [
61]. Given its critical role in BBB function and disease pathophysiology, GLUT1 remains a promising target for CNS drug delivery and therapeutic intervention [
62].
One approach involved modifying liposomal NPs with mannose, a known GLUT1 ligand, to facilitate targeted delivery in AD treatment [
60]. This strategy significantly enhanced brain-derived neurotrophic factor (BDNF) transport, achieving approximately 50% higher BBB permeability and a ~1.7-fold increase in BDNF levels in neuronal cells [
60]. Similarly, Xie et al. developed glucose-modified liposomes incorporating phospholipids, glucose-derived cholesterols, and polyethylene glycol (PEG) linkers to optimize brain-targeting efficiency [
63]. In one study, liposomes incorporating various glucose-conjugated cholesterols (GLU200-LIP, GLU1000-LIP, and GLU2000-LIP) demonstrated promising brain-targeting capabilities [
63]. In addition, novel multivalent glucoside-based ligands with strong affinity for GLUT1 have been developed for brain-targeted liposomal drug delivery. Liposomes functionalized with these glucosides and encapsulated with docetaxel achieved significantly higher brain accumulation of docetaxel compared to both free docetaxel and non-targeted liposome formulations [
64]. Xie et al. designed dual-sensitive nanomicelles functionalized with glucose molecules to enhance brain delivery of 3D6 antibody fragments (3D6-Fab) via GLUT1-mediated transport [
65]. This strategy achieved a 41-fold increase in brain accumulation of 3D6-Fab, representing a promising platform for efficient delivery of therapeutic antibodies to the CNS for the treatment of neurological diseases. Further advancing GLUT1-mediated transcytosis, Hao et al. introduced sequential targeting in crosslinking (STICK) NPs, a polymer-based delivery system designed to enhance both BBB penetration and tumor targeting [
66]. By incorporating maltobionic acid (a GLUT1-recognized glucose derivative) and 4-carboxyphenylboronic acid, this system improved drug stability, facilitated BBB transport, and significantly inhibited tumor growth in preclinical models of aggressive brain tumors [
66,
67]. Additionally, Zhou et al. developed glycosylated “triple-interaction” stabilized polymeric siRNA NPs (Gal-NP@siRNA) to target β-site amyloid precursor protein cleaving enzyme 1 (BACE1) in an amyloid precursor protein (APP)/PS1 transgenic AD mouse model [
68]. These NPs demonstrated superior blood stability and efficiently penetrated the BBB via glycemia-controlled GLUT1-mediated transport. Once in the brain, the siRNAs effectively reduced BACE1 expression, modulating key pathological pathways in AD progression (
Table 1) [
68].
These findings highlight the significant potential of GLUT1-targeted nanocarriers in CNS drug delivery. They demonstrate enhanced therapeutic efficacy through improved BBB transport, increased drug stability, and targeted disease modulation. However, challenges persist, particularly in studying human GLUTs in vivo, necessitating reliable in vitro models [
61]. Future research is needed to optimize ligand specificity, enhance transcytosis efficiency, and minimize off-target effects. Additionally, inter-individual variability in GLUT1 expression poses a challenge for clinical translation. Addressing these issues will be key to advancing GLUT1-mediated drug delivery for neurological disorders, brain tumors, and cerebrovascular diseases, paving the way for more effective CNS therapeutics.
5.2. L-Type Amino Acid Transporters (LATs)
L-type amino acid transporter 1 (LAT1) is a high-affinity, sodium-independent transporter that facilitates the uptake of large neutral amino acids, including leucine, phenylalanine, tryptophan, and tyrosine [
74]. Forming a heterodimeric complex with 4F2 heavy chain (SLC3A2), LAT1 is highly expressed in metabolically active tissues, such as the brain, placenta, testes, and various cancers, supporting protein synthesis, cell proliferation, and metabolic function [
74]. At the BBB, LAT1 is primarily located on the luminal membrane of ECs, ensuring the continuous influx of essential amino acids for neuronal and glial function [
75]. LAT1 is overexpressed in several cancers, including glioblastoma [
76], brain metastases [
77], and various solid tumors, as well as in neurological disorders, such as autism spectrum disorders [
78]. Additionally, LAT1 facilitates the transport of CNS-active drugs, such as melphalan [
79], levodopa (L-Dopa) [
80], gabapentin [
81], and pregabalin [
82], making it integral for developing prodrugs with enhanced BBB permeability [
83]. For example, LAT1 has been shown to exhibit a high maximal transport capacity (Vmax ≈ 40–60 nmol/min/g) and a substantial binding affinity (Km ≈ 10–200 µM), enabling the rapid exchange of high-affinity substrates across the BBB with half-times (half the time required to reach drug concentration equilibrium between the brain and blood) under 15 min [
83].
Recent advancements in LAT1-targeted drug delivery have focused on designing prodrugs and NPs that exploit LAT1-mediated transport. Montaser et al. developed LAT1-targeted prodrugs to enhance the delivery of nonsteroidal anti-inflammatory drugs (NSAIDs) to the brain. Salicylic acid derivatives exhibited a five-fold increase in brain uptake, but challenges such as plasma protein binding and premature bioconversion remain [
84]. Similarly, ferulic acid (FA)-based prodrugs have demonstrated LAT1-specific binding, enabling BBB penetration and cellular uptake. These amide-based prodrugs, incorporating an aromatic ring in the moiety, exhibited effective binding to LAT1, facilitating their cellular uptake in vitro and successfully crossing the BBB in mice [
85]. Beyond small molecules, LAT1-targeted NPs have shown promise. L-DOPA-functionalized gold nanoflowers (L-DOPA-AuNFs) exhibited superior BBB permeability and uptake by brain macrophages without inducing inflammation [
86]. In the context of glioma therapy, Zhang et al. developed LAT1-targeted liposomes co-loaded with temozolomide (TMZ) and sorafenib [
87]. The high expression of LAT1 on BBB and glioma cells enabled these liposomes to cross the BBB more efficiently, enhancing drug delivery to the tumor site and improving therapeutic outcomes (
Table 2) [
87].
Given its broad substrate specificity, high transport efficacy, and key role in both CNS and tumor metabolism, LAT1 represents a powerful avenue for brain-targeted drug delivery [
88]. Ongoing research into LAT1-mediated strategies, including optimized ligand conjugation, prodrug refinement, and NP engineering, holds promise for enhancing therapeutic outcomes in CNS diseases and brain tumors. By leveraging its strategic expression in key disease areas, LAT1 presents a critical opportunity for advancing more precise and effective treatments for complex neurological and oncological conditions. The following section delves into recent advancements based on LAT1-targeted strategies for overcoming the BBB.
Table 2.
Strategies to target L-type amino acid transporter 1 (LAT1) for brain-targeted drug delivery.
Table 2.
Strategies to target L-type amino acid transporter 1 (LAT1) for brain-targeted drug delivery.
| Drug | Malignancy | LAT1 Targeting Moiety | Delivery
Vehicle | Inference | Reference |
|---|
| Salicylic acid, ibuprofen, naproxen, and flurbiprofen | Neurodegenerative diseases | L-Phenylalanine | Prodrug | Significant (5-fold) increase in brain uptake across BBB | [84] |
| Ferulic acid | Alzheimer’s disease | L-Phenylalanine | Prodrug | Amide-based prodrug with aromatic ring effectively binds to LAT1, facilitating cellular uptake in vitro and crossing BBB | [84] |
| Levodopa | Brain malignancies like Parkinson’s disease | L-Phenylalanine | L-DOPA-functionalized gold NPs | Significantly higher penetration across BBB, increased internalization in brain macrophages, and inflammatory responses as compared to non-targeted NPs | [86] |
| Temozolomide and sorafenib | Glioblastoma | Tyrosine | Lipid NPs (carboxylated polyethylene glycol stearate and PLGA-PEG) | Enhanced drug delivery across BBB, increased tumor accumulation, and improved anti-tumor efficacy | [87] |
| Morin hydrate | Alzheimer’s disease | Phenylalanine-phenylalanine dipeptide | Dipeptide-functionalized PLGA NPs | Significantly improved brain retention with liver and lung accumulation | [89] |
| Antisense oligonucleotide | Neurodegenerative diseases | Phenylalanine | Phenylalanine-functionalized PEG-PLL NPs | Sixty-four-fold higher brain accumulation when compared to non-targeted NPs | [90] |
| Saxagliptin | Alzheimer’s disease | L-valine-conjugated chitosan | Chitosan-L-valine NPs | Enhanced BBB permeability with ~50-fold higher brain uptake than free drug and 3.4-fold lower plasma concentration, suggesting stability in plasma and release in brain tissue | [91] |
| Levodopa and dopamine | Parkinson’s disease | | Polyvinylpyrrolidone-functionalized selenium NPs | Facilitated BBB permeability and efficient internalization in brain endothelial cells | [92] |
| Phenylalanine analogs | Glioblastoma multiforme | Phenylalanine | Free drug | Iodine substitution at second position improved LAT1 affinity, but reduced velocity; reducing one carbon reduced LAT1 affinity, and bicyclic and α-methyl phenylalanine showed similar velocity, with preferential tumor accumulation of bicyclic phenylalanine | [93] |
5.3. Monocarboxylate Transporters (MCTs)
MCTs constitute a family of transmembrane proteins responsible for the transport of monocarboxylates, including lactate, pyruvate, ketone bodies (β-hydroxybutyrate and acetoacetate), and short-chain fatty acids, across biological membranes. These proton-coupled symporters facilitate energy-independent transport by co-transporting protons (H
+) along substrate concentration gradients [
40]. Among the MCT family, MCT1 (SLC16A1), MCT2 (SLC16A7), and MCT4 (SLC16A3) hold particular significance in brain metabolism and BBB function.
At the BBB, MCT1 is predominantly expressed on the luminal membrane of endothelial cells, facilitating the uptake of ketone bodies during periods of low glucose availability, such as fasting, exercise, or ketogenic diets [
44,
94,
95]. Additionally, MCT1 plays a crucial role in transporting lactate from the bloodstream into the brain, where it serves as an energy substrate for neurons and glial cells [
96]. MCT2, primarily expressed in neurons, supports lactate uptake for metabolic processes, whereas MCT4, found in astrocytes, facilitates lactate export to sustain neuronal energy demands [
97]. Given their essential role in brain metabolism, MCTs have emerged as promising targets for drug delivery, particularly in conditions associated with metabolic dysfunction and neurodegeneration.
Taking advantage of the natural substrate affinity of MCT1, researchers have developed MCT1-mediated drug delivery systems, including NPs, prodrugs, and inhibitors, to enhance drug penetration into the brain. For instance, Venishetty et al. designed β-hydroxybutyric acid (HBA)-grafted docetaxel-loaded solid lipid NPs (HD-SLNs) to exploit MCT1-mediated transport, achieving significantly higher brain concentrations of docetaxel than Taxotere
® [
98]. This system also demonstrated enhanced cellular uptake and controlled drug release, highlighting its potential for CNS drug delivery [
98]. Similarly, Güliz Ak et al. developed HBA-conjugated solid lipid NPs (SLNs) for glioblastoma therapy, facilitating the transport of carmustine (BCNU) and TMZ across the BBB [
99]. These dual drug-loaded SLNs exhibited controlled release, increased cytotoxicity against glioblastoma cells, and reduced toxicity to healthy cells, making them a promising targeted therapy for glioblastoma multiforme (GBM) [
99]. Beyond drug-loaded NPs, inhibitors targeting MCT1 have shown potential in modulating tumor metabolism. Pluronic P85 (P85) NPs were found to inhibit MCT1-mediated lactate transport across brain microvascular ECs without significantly affecting GLUT1 function. This demonstrated a concentration-dependent impact on brain monocarboxylate metabolism, while maintaining safety [
100]. In another approach, Huang et al. developed ultra-pH-sensitive NPs encapsulating AZD3965, a selective MCT1 inhibitor [
101]. This formulation effectively blocked MCT1 activity in tumors, reversing lactic acid-induced immunosuppression and enhancing the effectiveness of cancer immunotherapy, while reducing systemic toxicity [
101]. AZD3965 exhibited rapid oral absorption, high bioavailability, and target engagement with minimal systemic toxicity. However, its potential for brain cancer therapy remains underexplored [
102]. The study also revealed nonlinear pharmacokinetics, with dose-dependent increases in exposure and changes in clearance, indicating potential target-mediated drug disposition (TMDD) for AZD3965 [
102]. Given its ability to inhibit MCT1 in tumors expressing high MCT1 levels, AZD3965 could enhance drug penetration across the BBB and modulate the tumor microenvironment, warranting further investigation in glioblastoma models (
Table 3).
In conclusion, MCT1-targeted drug delivery has demonstrated significant potential for improving brain drug penetration, particularly in glioblastoma and other CNS disorders. The development of HBA-grafted NPs and MCT1 inhibitors such as AZD3965 offers promising avenues for overcoming the BBB limitations and enhancing drug efficacy. However, further research is necessary to optimize these formulations, assess their long-term safety, and expand their clinical applicability. Additionally, more specific studies focusing on MCT1’s role in brain tumors are crucial for refining these strategies and improving therapeutic outcomes for patients with CNS malignancies.
5.4. Organic Anion-Transporting Polypeptides (OATPs)
OATPs, a subfamily of the solute carrier (SLC) superfamily, play a critical role in the uptake and distribution of amphipathic molecules, including steroid hormones, bile acids, statins, antihypertensives, antibiotics, antifungals, and chemotherapeutic agents [
104]. Their broad tissue distribution and capacity to transport structurally diverse compounds make them key regulators of drug disposition in major organs, including the liver, kidney, intestine, and brain [
104,
105]. OATP isoforms such as OATP1B1, OATP1B3, OATP1A2, and OATP2B1 are particularly relevant to BBB transport, influencing CNS drug permeability and offering potential targets for enhancing drug delivery to the brain [
104,
105]. Given their role in modulating drug transport across the BBB, OATPs also represent promising targets for enhancing CNS drug delivery, particularly in neurological disorders, where drug penetration remains a significant challenge.
The expression and function of OATPs are influenced by genetic polymorphisms, age, gender, and dietary factors, contributing to inter-individual variability in drug absorption, distribution, efficacy, and toxicity. Genetic polymorphisms in OATP transporters also impact their functional capacity, leading to variations in pharmacokinetics and pharmacodynamics. Notably, OATP1A4 and OATP1A5 exhibit age- and gender-specific expression patterns, affecting brain drug-transport dynamics [
106,
107,
108]. A deeper understanding of these variations is crucial for optimizing CNS drug delivery strategies, particularly in neurological disorders. Advancements in nanotechnology have enabled the development of OATP-targeted drug delivery systems to enhance CNS drug uptake, offering novel insights into transporter-mediated brain drug delivery, improving BBB penetration and therapeutic efficacy [
109,
110]. For example, Yang et al. developed OATP2B1-targeted dendrigraft poly-l-lysine-polyethylene glycol NPs for siRNA delivery, demonstrating enhanced targeting precision and gene-silencing efficiency in the mouse brain [
109]. Similarly, Reichel et al. designed the HMC-FMX nanoprobe, which facilitated OATP-mediated BBB transport and glioblastoma accumulation, highlighting its potential for brain tumor therapy [
110]. Additionally, Thompson et al. found that hypoxia/reoxygenation stress upregulates OATP1A4 expression at the BBB, enhancing atorvastatin transport into the brain, suggesting its potential as a drug delivery target (
Table 4) [
111].






{kind=link}
{kind=link}