
Targeted Polymer–Peptide Conjugates for E-Selectin Blockade in Renal Injury


Abstract
1. Introduction
2. Materials and Methods
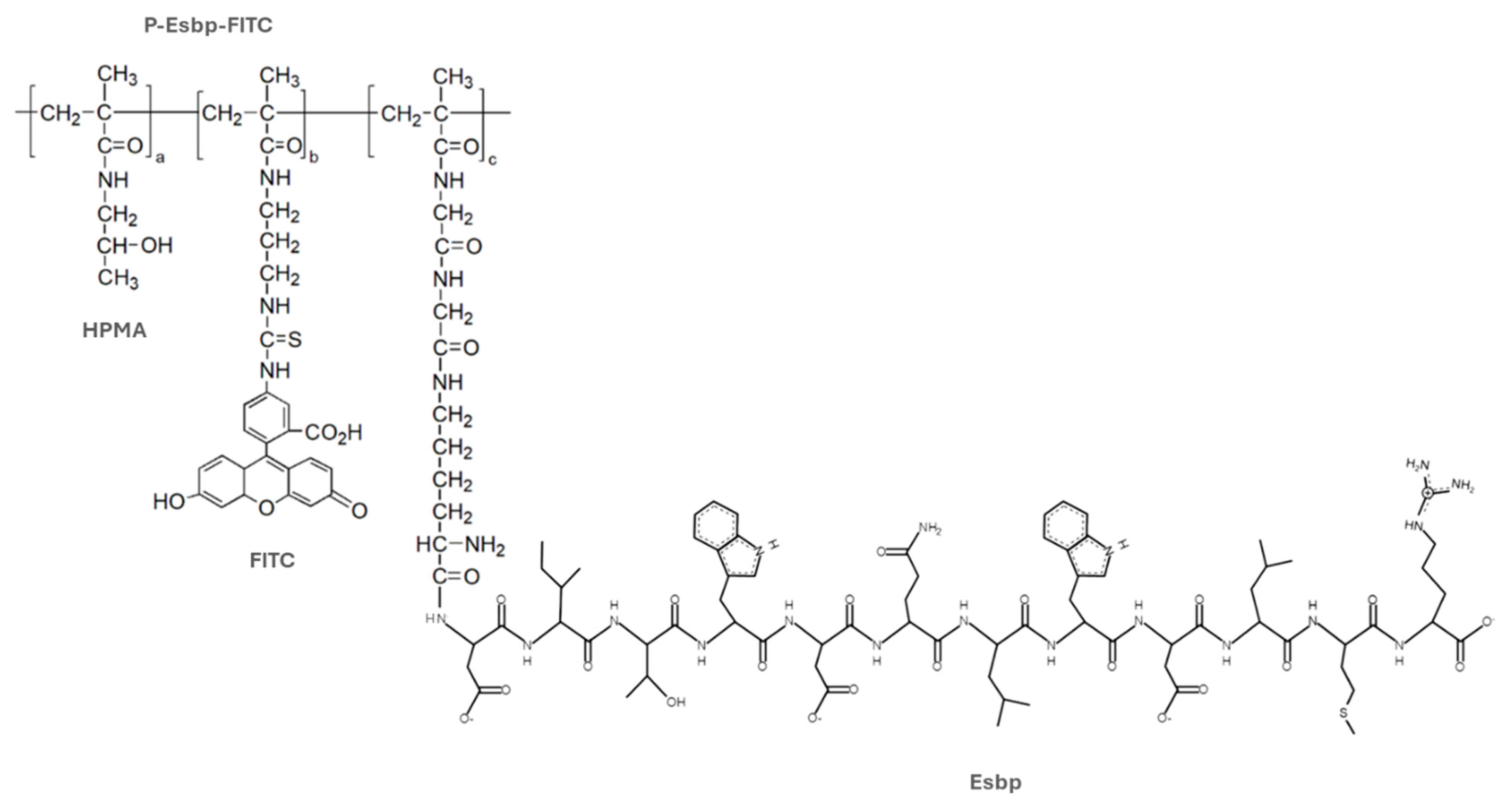
2.1. Chemical Synthesis and Characterization of Polymers P-Esbp
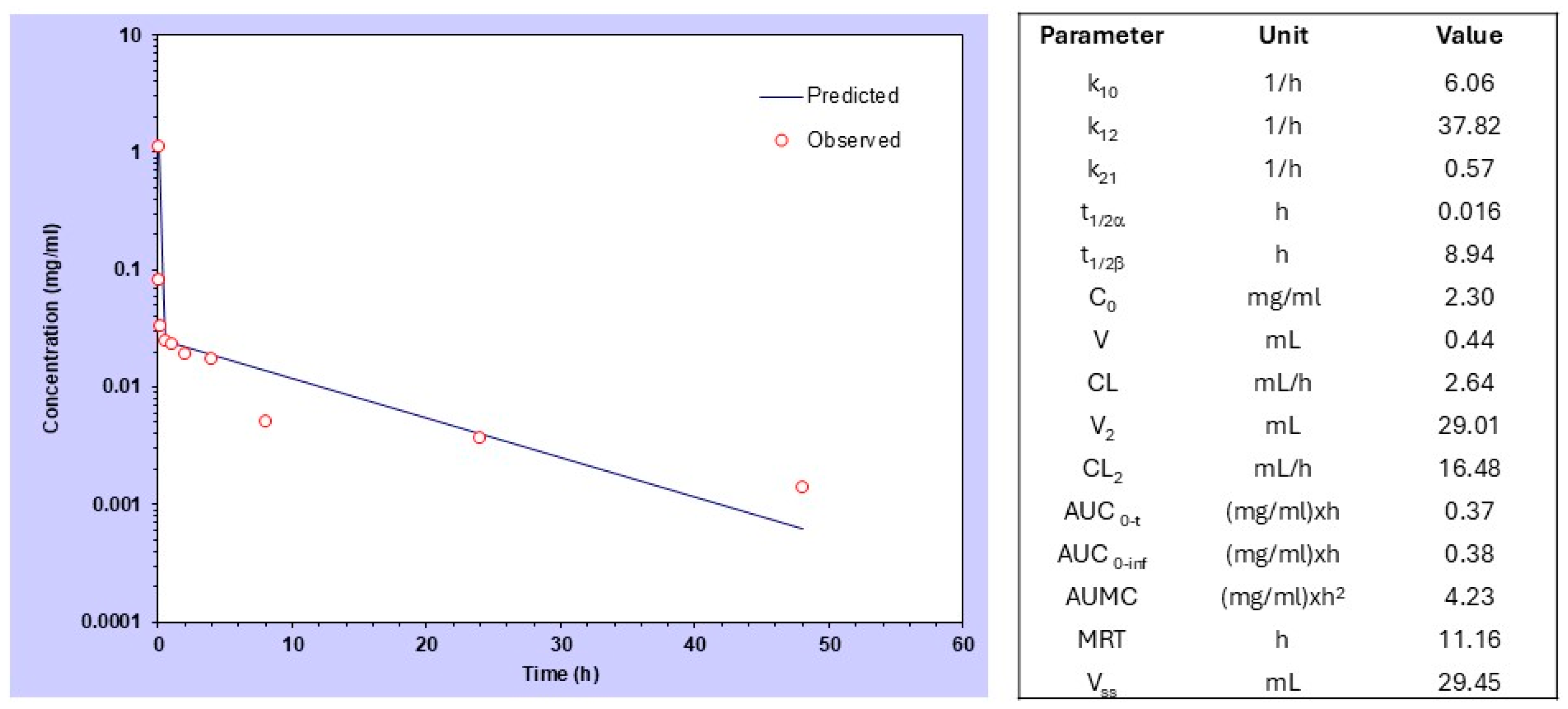
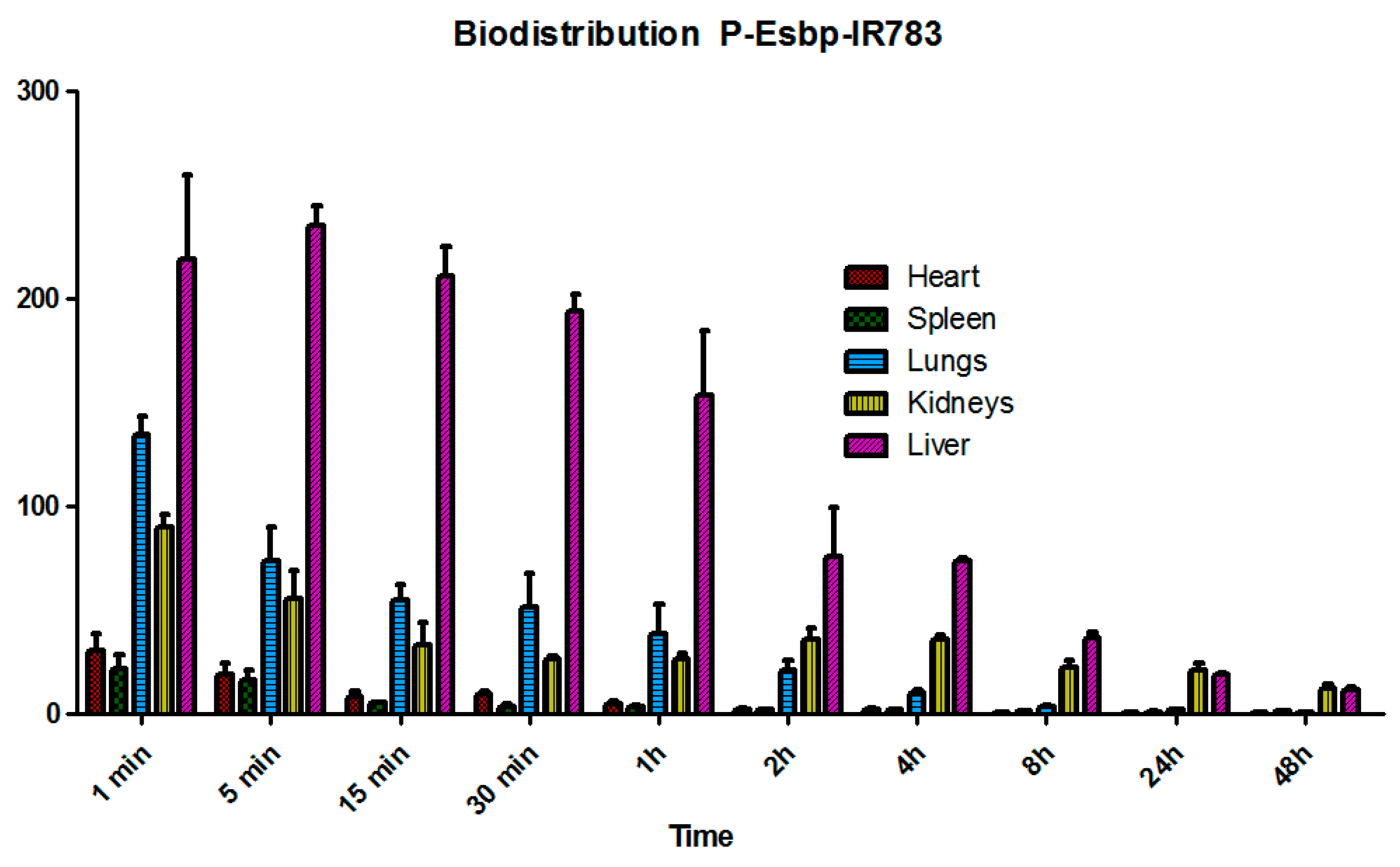
2.2. Pharmacokinetic and Biodistribution Analysis of P-(Esbp)-IR783
2.3. Animal Models of Kidney Inflammation
2.3.1. In Vivo Model of Acute Kidney Inflammation—Renal Ischemia–Reperfusion Injury Model (RIRI)
2.3.2. In Vivo Model of Adenine-Induced Chronic Kidney Disease (CKD)
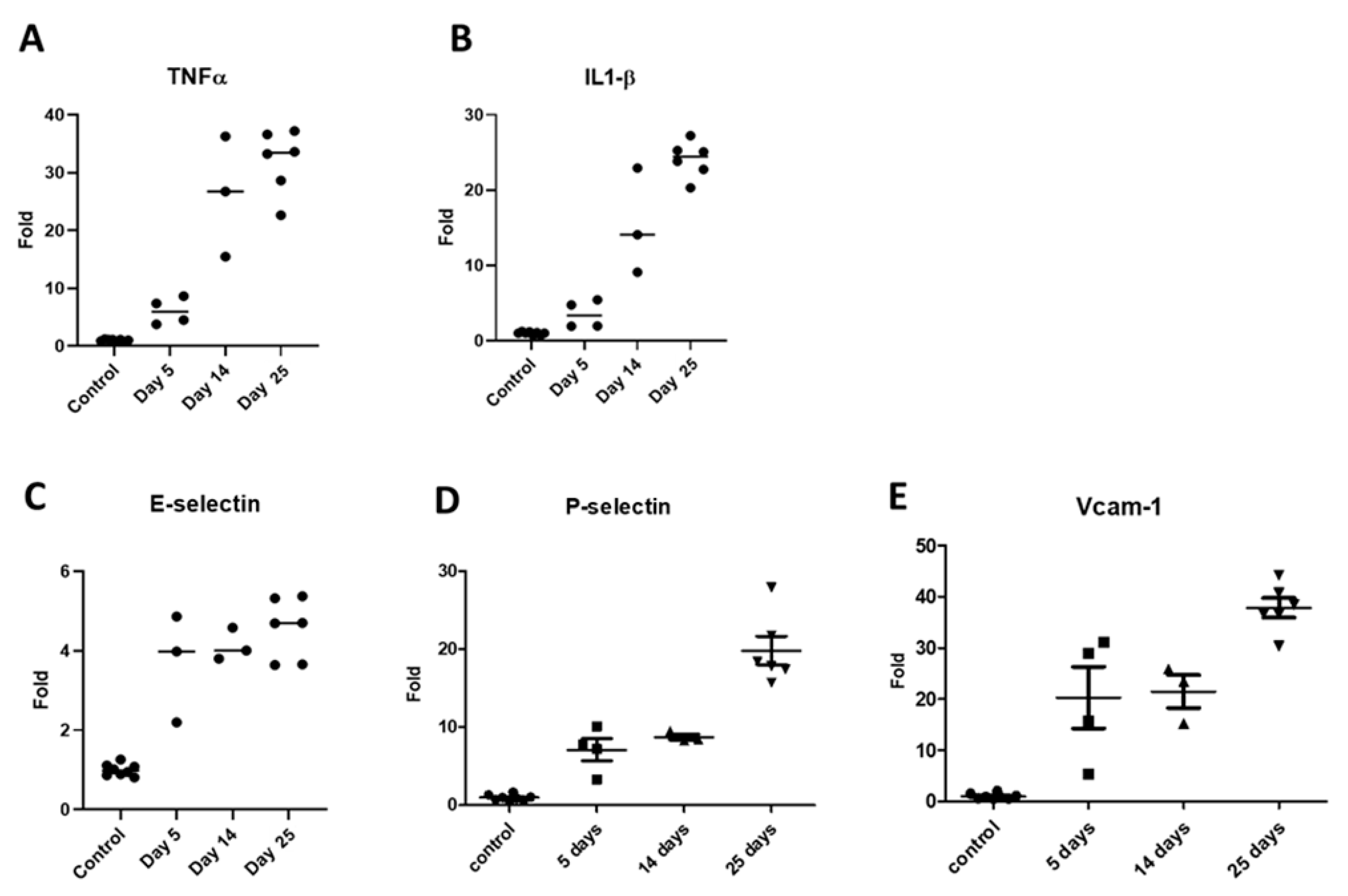
- Experimental Design of Adenine-Induced CKD Murine Model
- Tissue and Serum Processing and Analysis
3. Results
3.1. Targeted Polymer Synthesis
3.2. Pharmacokinetics (PK) and Biodistribution (BD) Analysis of P-(Esbp)-IR783
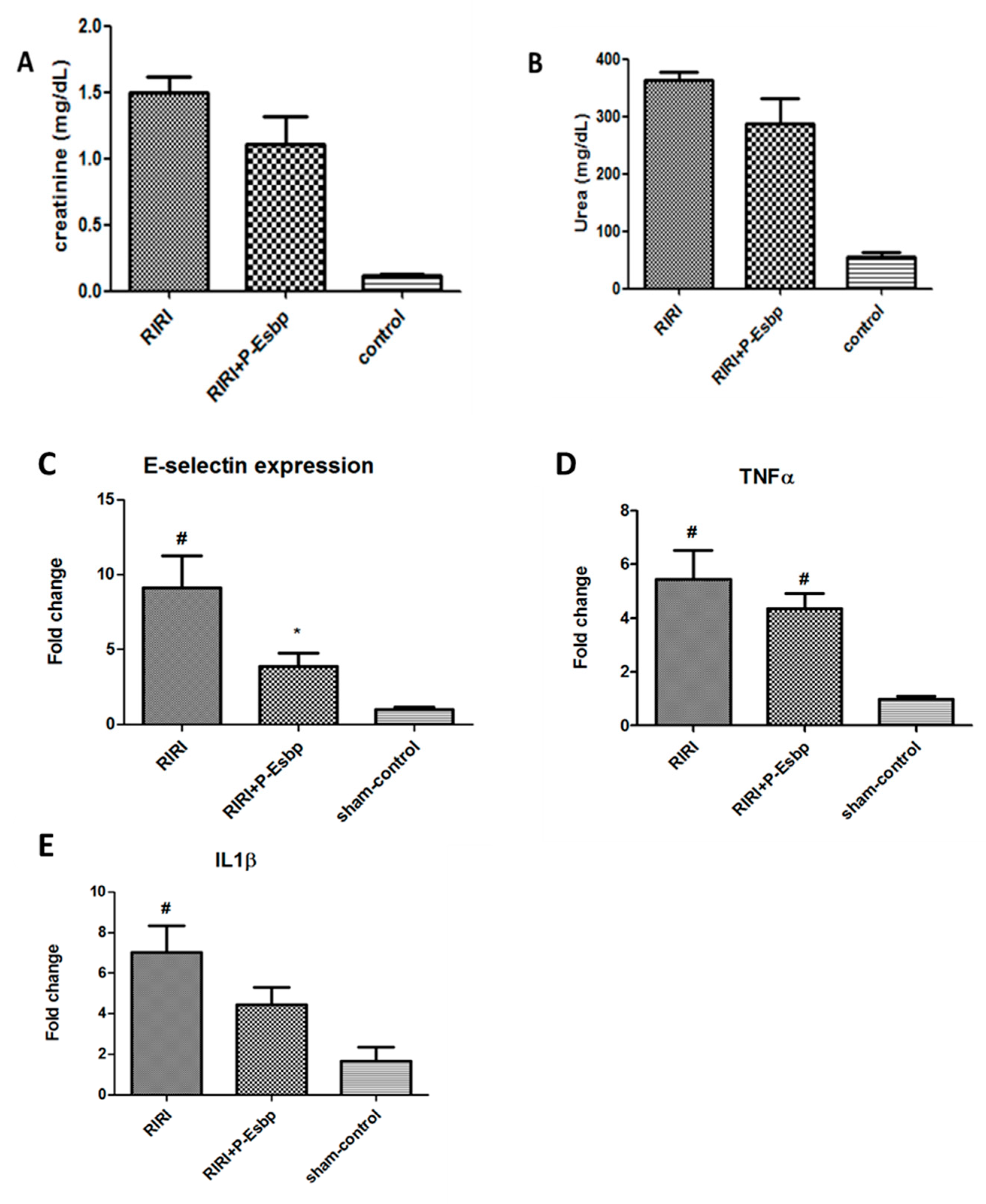
3.3. P-Esbp Reduced Kidney Damage in Acute Kidney Inflammation—Renal Ischemia–Reperfusion Injury (RIRI) Model
3.4. Continuous P-Esbp Treatment Did Not Affect Chronic Kidney Injury in Adenine-Induced CKD Model
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chawla, L.S.; Eggers, P.W.; Star, R.A.; Kimmel, P.L. Acute kidney injury and chronic kidney disease as interconnected syndromes. N. Engl. J. Med. 2014, 371, 58–66. [Google Scholar] [CrossRef]
- Pickkers, P.; Murray, P.T.; Ostermann, M. New drugs for acute kidney injury. Intensive Care Med. 2022, 48, 1796–1798. [Google Scholar] [CrossRef]
- Kinsey, G.R.; Okusa, M.D. Role of leukocytes in the pathogenesis of acute kidney injury. Crit. Care 2012, 16, 214. [Google Scholar] [CrossRef] [PubMed]
- Hopkin, S.J.; Lewis, J.W.; Krautter, F.; Chimen, M.; McGettrick, H.M. Triggering the resolution of immune mediated inflammatory diseases: Can targeting leukocyte migration be the answer? Front. Pharmacol. 2019, 10, 184. [Google Scholar] [CrossRef] [PubMed]
- Moreira, M.B.; Garcia-Cardeña, G.; Saffi, M.A.L.; Libby, P. Endothelium: A coordinator of acute and chronic inflammation. In Endothelium and Cardiovascular Diseases; Protasio, L.D.L., Libby, P., Chagas, A.C.P., Laurindo, F.R.M., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 485–491. [Google Scholar]
- Kreuger, J.; Phillipson, M. Targeting vascular and leukocyte communication in angiogenesis, inflammation and fibrosis. Nat. Rev. Drug Discov. 2016, 15, 125–142. [Google Scholar] [CrossRef]
- Smith, C.W. 3. Adhesion molecules and receptors. J. Allergy Clin. Immunol. 2008, 121, S375–S379. [Google Scholar] [CrossRef] [PubMed]
- Peres, R.S.; Menezes, G.B.; Teixeira, M.M.; Cunha, F.Q. Pharmacological opportunities to control inflammatory diseases through inhibition of the leukocyte recruitment. Pharmacol. Res. 2016, 112, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Rymer, J.A.; Newby, L.K. Failure to launch: Targeting inflammation in acute coronary syndromes. Basic. Transl. Sci. 2017, 2, 484–497. [Google Scholar]
- Seekamp, A.; van Griensven, M.; Rusu, C.; König, J.; Khan-Boluki, J.; Redl, H. The effect of anti-L-selectin (Aselizumab) on the posttraumatic inflammatory response in multiply traumatized patients. Eur. J. Trauma 2005, 31, 557–567. [Google Scholar] [CrossRef]
- Romano, S.J. Selectin antagonists. Treat. Respir. Med. 2005, 4, 85–94. [Google Scholar] [CrossRef]
- Beeh, K.M.; Beier, J.; Meyer, M.; Buhl, R.; Zahlten, R.; Wolff, G. Bimosiamose, an inhaled small-molecule pan-selectin antagonist, attenuates late asthmatic reactions following allergen challenge in mild asthmatics: A randomized, double-blind, placebo-controlled clinical cross-over-trial. Pulm. Pharmacol. Ther. 2006, 19, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M.J.; Billingsley, M.M.; Haley, R.M.; Wechsler, M.E.; Peppas, N.A.; Langer, R. Engineering precision nanoparticles for drug delivery. Nat. Rev. Drug Discov. 2021, 20, 101–124. [Google Scholar] [CrossRef] [PubMed]
- Rütter, M.; Milošević, N.; David, A. Say no to drugs: Bioactive macromolecular therapeutics without conventional drugs. J. Control. Release 2021, 330, 1191–1207. [Google Scholar] [CrossRef]
- Milošević, N.; Rütter, M.; David, A. Endothelial cell adhesion molecules-(un) Attainable targets for nanomedicines. Front. Med. Technol. 2022, 4, 846065. [Google Scholar] [CrossRef] [PubMed]
- Shamay, Y.; Paulin, D.; Ashkenasy, G.; David, A. E-selectin binding peptide–polymer–drug conjugates and their selective cytotoxicity against vascular endothelial cells. Biomaterials 2009, 30, 6460–6468. [Google Scholar] [CrossRef] [PubMed]
- Shamay, Y.; Raviv, L.; Golan, M.; Voronov, E.; Apte, R.N.; David, A. Inhibition of primary and metastatic tumors in mice by E-selectin-targeted polymer–drug conjugates. J. Control. Release 2015, 217, 102–112. [Google Scholar] [CrossRef]
- Rütter, M.; Milošević, N.; Ventura, Y.; Feinshtein, V.; David, A. E-selectin-targeted polymer-doxorubicin conjugate induces regression of established colorectal liver metastases and improves mice survival. Nano Today 2024, 55, 102182. [Google Scholar] [CrossRef]
- Tsoref, O.; Tyomkin, D.; Amit, U.; Landa, N.; Cohen-Rosenboim, O.; Kain, D.; Golan, M.; Naftali-Shani, N.; David, A.; Leor, J. E-selectin-targeted copolymer reduces atherosclerotic lesions, adverse cardiac remodeling, and dysfunction. J. Control. Release 2018, 288, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Milošević, N.; Rütter, M.; Ventura, Y.; Kezerle, Y.; Feinshtein, V.; David, A. Attenuation of neutrophil-mediated liver injury in mice by drug-free E-selectin binding polymer. J. Control. Release 2020, 319, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Kato, N.; Yuzawa, Y.; Kosugi, T.; Hobo, A.; Sato, W.; Miwa, Y.; Sakamoto, K.; Matsuo, S.; Kadomatsu, K. The E-selectin ligand basigin/CD147 is responsible for neutrophil recruitment in renal ischemia/reperfusion. J. Am. Soc. Nephrol. 2009, 20, 1565–1576. [Google Scholar] [CrossRef]
- Jia, T.; Olauson, H.; Lindberg, K.; Amin, R.; Edvardsson, K.; Lindholm, B.; Andersson, G.; Wernerson, A.; Sabbagh, Y.; Schiavi, S. A novel model of adenine-induced tubulointerstitial nephropathy in mice. BMC Nephrol. 2013, 14, 116. [Google Scholar] [CrossRef] [PubMed]
- Klinkhammer, B.M.; Djudjaj, S.; Kunter, U.; Palsson, R.; Edvardsson, V.O.; Wiech, T.; Thorsteinsdottir, M.; Hardarson, S.; Foresto-Neto, O.; Mulay, S.R.; et al. Cellular and molecular mechanisms of kidney injury in 2,8-dihydroxyadenine nephropathy. J. Am. Soc. Nephrol. JASN 2020, 31, 799. [Google Scholar] [CrossRef] [PubMed]
- Singbartl, K.; Ley, K. Protection from ischemia-reperfusion induced severe acute renal failure by blocking E-selectin. Crit. Care Med. 2000, 28, 2507–2514. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Dong, Z. Mouse model of ischemic acute kidney injury: Technical notes and tricks. Am. J. Physiol. Ren. Physiol. 2012, 303, F1487–F1494. [Google Scholar] [CrossRef] [PubMed]
- Hesketh, E.E.; Czopek, A.; Clay, M.; Borthwick, G.; Ferenbach, D.; Kluth, D.; Hughes, J. Renal ischaemia reperfusion injury: A mouse model of injury and regeneration. J. Vis. Exp. 2014, 88, e51816. [Google Scholar] [CrossRef]
- Shiah, J.-G.; Dvořák, M.; Kopečková, P.; Sun, Y.; Peterson, C.; Kopeček, J. Biodistribution and antitumour efficacy of long-circulating N-(2-hydroxypropyl) methacrylamide copolymer–doxorubicin conjugates in nude mice. Eur. J. Cancer 2001, 37, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Blanco, E.; Shen, H.; Ferrari, M. Principles of nanoparticle design for overcoming biological barriers to drug delivery. Nat. Biotechnol. 2015, 33, 941–951. [Google Scholar] [CrossRef]
- Santana, A.C.; Degaspari, S.; Catanozi, S.; Dellê, H.; de Sá Lima, L.; Silva, C.; Blanco, P.; Solez, K.; Scavone, C.; Noronha, I.L. Thalidomide suppresses inflammation in adenine-induced CKD with uraemia in mice. Nephrol. Dial. Transplant. 2013, 28, 1140–1149. [Google Scholar] [CrossRef] [PubMed]
- de Frutos, S.; Luengo, A.; Garcia-Jerez, A.; Hatem-Vaquero, M.; Griera, M.; O’Valle, F.; Rodríguez–Puyol, M.; Rodríguez–Puyol, D.; Calleros, L. Chronic kidney disease induced by an adenine rich diet upregulates integrin linked kinase (ILK) and its depletion prevents the disease progression. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2019, 1865, 1284–1297. [Google Scholar] [CrossRef]
- Ludwig-Portugall, I.; Bartok, E.; Dhana, E.; Evers, B.D.; Primiano, M.J.; Hall, J.P.; Franklin, B.S.; Knolle, P.A.; Hornung, V.; Hartmann, G.; et al. An NLRP3-specific inflammasome inhibitor attenuates crystal-induced kidney fibrosis in mice. Kidney Int. 2016, 90, 525–539. [Google Scholar] [CrossRef] [PubMed]
- Tamura, M.; Aizawa, R.; Hori, M.; Ozaki, H. Progressive renal dysfunction and macrophage infiltration in interstitial fibrosis in an adenine-induced tubulointerstitial nephritis mouse model. Histochem. Cell Biol. 2009, 131, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Herter, J.M.; Rossaint, J.; Spieker, T.; Zarbock, A. Adhesion molecules involved in neutrophil recruitment during sepsis-induced acute kidney injury. J. Innate Immun. 2014, 6, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Singbartl, K.; Green, S.A. LEYK Blocking P-selectin protects from ischemia/reperfusion-induced acute renal failure. FASEB J. 2000, 14, 48–54. [Google Scholar] [CrossRef]
- Nemoto, T.; Burne, M.J.; Daniels, F.; O’Donnell, M.P.; Crosson, J.; Berens, K.; Issekutz, A.; Kasiske, B.L.; Keane, W.F.; Rabb, H. Small molecule selectin ligand inhibition improves outcome in ischemic acute renal failure. Kidney Int. 2001, 60, 2205–2214. [Google Scholar] [CrossRef]
- Bertola, A.; Park, O.; Gao, B. Chronic plus binge ethanol feeding synergistically induces neutrophil infiltration and liver injury in mice: A critical role for E-selectin. Hepatology 2013, 58, 1814–1823. [Google Scholar] [CrossRef]
- Akhtar, A.M.; Schneider, J.E.; Chapman, S.J.; Jefferson, A.; Digby, J.E.; Mankia, K.; Chen, Y.; McAteer, M.A.; Wood, K.J.; Choudhury, R.P. In vivo quantification of VCAM-1 expression in renal ischemia reperfusion injury using non-invasive magnetic resonance molecular imaging. PLoS ONE 2010, 5, e12800. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Li, R.; Chen, X.; Yan, H.; Li, H.; Zhao, X.; Chen, Y.; McAteer, M.A.; Wood, K.J.; Choudhury, R.P. Renal Endothelial Cell-Targeted Extracellular Vesicles Protect the Kidney from Ischemic Injury. Adv. Sci. 2023, 10, e2204626. [Google Scholar] [CrossRef] [PubMed]
- Boesen, E.I.; Crislip, G.R.; Sullivan, J.C. Use of ultrasound to assess renal reperfusion and P-selectin expression following unilateral renal ischemia. Am. J. Physiol. Ren. Physiol. 2012, 303, F1333–F1340. [Google Scholar] [CrossRef] [PubMed]
- Mayer, C.L.; Koeck, K.; Hottmann, M.; Redfern, A.; Davis, M.; Barth, A.; Geng, X.; Hoppe, C.; Yue, P. A phase 1 study in healthy participants to characterize the safety and pharmacology of inclacumab, a fully human anti-P-selectin antibody, in development for treatment of sickle cell disease. Eur. J. Clin. Pharmacol. 2023, 79, 1219–1228. [Google Scholar] [CrossRef] [PubMed]
- Ailuno, G.; Baldassari, S.; Zuccari, G.; Schlich, M.; Caviglioli, G. Peptide-based nanosystems for vascular cell adhesion molecule-1 targeting: A real opportunity for therapeutic and diagnostic agents in inflammation associated disorders. J. Drug Deliv. Sci. Technol. 2020, 55, 101461. [Google Scholar] [CrossRef]
- Xu, J.; Zhou, J.; Zhong, Y.; Zhang, Y.; Ye, M.; Hou, J.; Wang, Z.; Ran, H.; Liu, J.; Guo, D. EWVDV-Mediated Platelet-Targeting Nanoparticles for the Multimodal Imaging of Thrombi at Different Blood Flow Velocities. Int. J. Nanomed. 2020, 15, 1759. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HPMA Copolymer | Mw [kDa] a | Polydispersity b | %mol FITC or IR783 c | %mol ONp/Peptide/Scrm d |
|---|---|---|---|---|
| P-(GGONp)-FITC | 34.0 | 1.42 | 1.8 | 7.50 |
| P-(Esbp)-FITC | 34.1 | 1.35 | 1.8 | 7.49 |
| P-(EsbpScrm)-FITC | 33.2 | 1.2 | 1.8 | 7.35 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milošević, N.; Rütter, M.; Ventura, Y.; Feinshtein, V.; David, A. Targeted Polymer–Peptide Conjugates for E-Selectin Blockade in Renal Injury. Pharmaceutics 2025, 17, 82. https://doi.org/10.3390/pharmaceutics17010082
Milošević N, Rütter M, Ventura Y, Feinshtein V, David A. Targeted Polymer–Peptide Conjugates for E-Selectin Blockade in Renal Injury. Pharmaceutics. 2025; 17(1):82. https://doi.org/10.3390/pharmaceutics17010082
Chicago/Turabian StyleMilošević, Nenad, Marie Rütter, Yvonne Ventura, Valeria Feinshtein, and Ayelet David. 2025. "Targeted Polymer–Peptide Conjugates for E-Selectin Blockade in Renal Injury" Pharmaceutics 17, no. 1: 82. https://doi.org/10.3390/pharmaceutics17010082
APA StyleMilošević, N., Rütter, M., Ventura, Y., Feinshtein, V., & David, A. (2025). Targeted Polymer–Peptide Conjugates for E-Selectin Blockade in Renal Injury. Pharmaceutics, 17(1), 82. https://doi.org/10.3390/pharmaceutics17010082