
Effects of Combined Treatment with Sodium Dichloroacetate and Sodium Valproate on the Genes in Inflammation- and Immune-Related Pathways in T Lymphocytes from Patients with SARS-CoV-2 Infection with Pneumonia: Sex-Related Differences

Abstract
1. Introduction
2. Materials and Methods
2.1. The Investigational Medicinal Product
2.2. Patients with SARS-CoV-2 Infection and Pneumonia
2.3. Isolation of T Lymphocytes from Human Venous Blood by Ficoll-Paque™ Density Gradient and Treatment
2.4. Total RNA Extraction and Next-Generation Sequencing
2.5. Bioinformatic Analysis
3. Results
3.1. Data on the Effect of the DCA–VPA Treatment on the Expression of Genes Related to Inflammation and Immune Response in T Lymphocytes from Men with SARS-CoV-2 Infection and Pneumonia
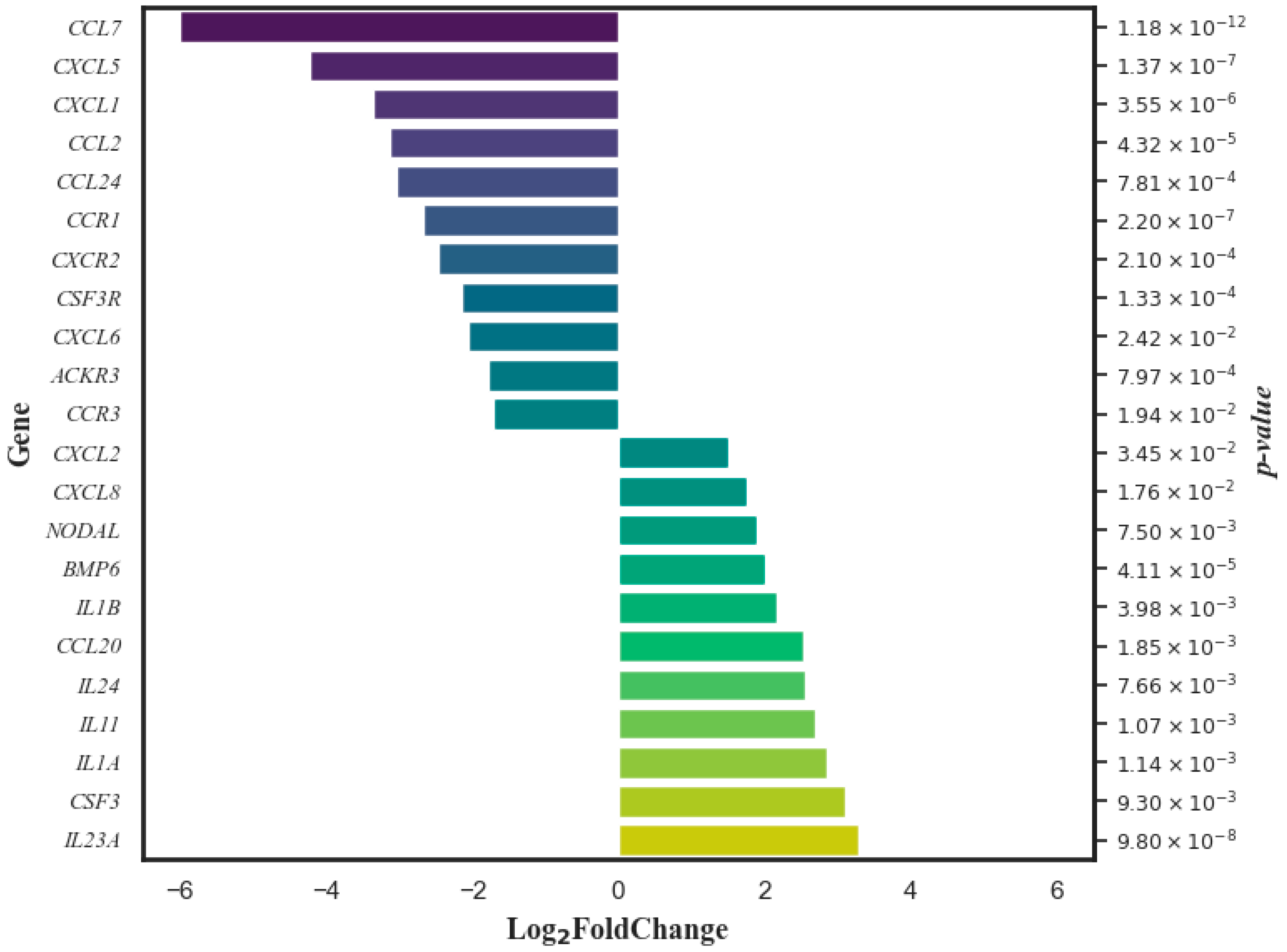
3.2. Data on the Effect of the DCA–VPA Treatment on the Expression of Genes Related to Inflammation and Immune Response in T Lymphocytes from Females with SARS-CoV-2 Infection with Pneumonia
3.3. Sex-Specific Gene Expression Responses to DCA-VPA Treatment in T Lymphocytes of SARS-CoV-2-Infected Patients with Pneumonia
4. Discussion
4.1. DCA–VPA Effects on Cytokine Activity and Sex-Related Differences
4.2. DCA–VPA Effects on Chemokine-Mediated Signaling, Neutrophil Chemotaxis, Lymphocyte Chemotaxis, and Regulation of T-Cell Proliferation Pathways and Sex-Related Differences
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stakišaitis, D.; Kapočius, L.; Kilimaitė, E.; Gečys, D.; Šlekienė, L.; Balnytė, I.; Palubinskienė, J.; Lesauskaitė, V. Preclinical Study in Mouse Thymus and Thymocytes: Effects of Treatment with a Combination of Sodium Dichloroacetate and Sodium Valproate on Infectious Inflammation Pathways. Pharmaceutics 2023, 15, 2715. [Google Scholar] [CrossRef] [PubMed]
- Brandenburg, K.; Ferrer-Espada, R.; Martinez-de-Tejada, G.; Nehls, C.; Fukuoka, S.; Mauss, K.; Weindl, G.; Garidel, P. A Comparison between SARS-CoV-2 and Gram-Negative Bacteria-Induced Hyperinflammation and Sepsis. Int. J. Mol. Sci. 2023, 24, 15169. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Qin, S.; Li, Z.; Gao, W.; Tang, M.; Dong, X. Early immune system alterations in patients with septic shock. Front. Immunol. 2023, 14, 1126874. [Google Scholar] [CrossRef] [PubMed]
- Fleischmann, C.; Scherag, A.; Adhikari, N.K.J.; Hartog, C.S.; Tsaganos, T.; Schlattmann, P.; Angus, D.C.; Reinhart, K. International Forum of Acute Care Trialists, Assessment of Global Incidence and Mortality of Hospital-treated Sepsis. Current Estimates and Limitations. Am. J. Respir. Crit. Care Med. 2016, 193, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Vincent, J.-L.; Bakker, J. Blood lactate levels in sepsis: In 8 questions. Curr. Opin. Crit. Care 2021, 27, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Shoraka, S.; Samarasinghe, A.E.; Ghaemi, A.; Mohebbi, S.R. Host mitochondria: More than an organelle in SARS-CoV-2 infection. Front. Cell Infect. Microbiol. 2023, 13, 1228275. [Google Scholar] [CrossRef] [PubMed]
- Langley, R.J.; Tsalik, E.L.; van Velkinburgh, J.C.; Glickman, S.W.; Rice, B.J.; Wang, C.; Chen, B.; Carin, L.; Suarez, A.; Mohney, R.P.; et al. An integrated clinico-metabolomic model improves prediction of death in sepsis. Sci. Transl. Med. 2013, 5, 195ra95. [Google Scholar] [CrossRef]
- Kotas, M.E.; Medzhitov, R. Homeostasis, inflammation, and disease susceptibility. Cell 2015, 816–827. [Google Scholar] [CrossRef]
- Soares, M.P.; Teixeira, L.; Moita, L.F. Disease tolerance and immunity in host protection against infection. Nat. Rev. Immunol. 2017, 17, 83–96. [Google Scholar] [CrossRef]
- Bhowal, C.; Ghosh, S.; Ghatak, D.; De, R. Pathophysiological involvement of host mitochondria in SARS-CoV-2 infection that causes COVID-19: A comprehensive evidential insight. Mol. Cell Biochem. 2023, 478, 1325–1343. [Google Scholar] [CrossRef]
- Vardavas, C.I.; Mathioudakis, A.G.; Nikitara, K.; Stamatelopoulos, K.; Georgiopoulos, G.; Phalkey, R.; Leonardi-Bee, J.; Fernandez, E.; Carnicer-Pont, D.; Vestbo, J.; et al. Prognostic factors for mortality, intensive care unit and hospital admission due to SARS-CoV-2: A systematic review and meta-analysis of cohort studies in Europe. Eur. Respir. Rev. 2022, 31, 220098. [Google Scholar] [CrossRef]
- Peckham, H.; de Gruijter, N.M.; Raine, C.; Radziszewska, A.; Ciurtin, C.; Wedderburn, L.R.; Rosser, E.C.; Webb, K.; Deakin, C.T. Male sex identified by global COVID-19 meta-analysis as a risk factor for death and ITU admission. Nat. Commun. 2020, 11, 6317. [Google Scholar] [CrossRef] [PubMed]
- Gebhard, C.; Regitz-Zagrosek, V.; Neuhauser, H.K.; Morgan, R.; Klein, S.L. Impact of sex and gender on COVID-19 outcomes in Europe. Biol. Sex. Differ. 2020, 11, 29. [Google Scholar] [CrossRef] [PubMed]
- Williamson, E.J.; Walker, A.J.; Bhaskaran, K.; Bacon, S.; Bates, C.; Morton, C.E.; Curtis, H.J.; Mehrkar, A.; Evans, D.; Inglesby, P.; et al. Factors associated with COVID-19-related death using OpenSAFELY. Nature 2020, 584, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Grasselli, G.; Zangrillo, A.; Zanella, A.; Antonelli, M.; Cabrini, L.; Castelli, A.; Cereda, D.; Coluccello, A.; Foti, G.; Fumagalli, R.; et al. Baseline Characteristics and Outcomes of 1591 Patients Infected With SARS-CoV-2 Admitted to ICUs of the Lombardy Region, Italy. JAMA 2020, 323, 1574–1581. [Google Scholar] [CrossRef] [PubMed]
- Stakišaitis, D.; Kapočius, L.; Valančiūtė, A.; Balnytė, I.; Tamošuitis, T.; Vaitkevičius, A.; Sužiedėlis, K.; Urbonienė, D.; Tatarūnas, V.; Kilimaitė, E.; et al. SARS-CoV-2 Infection; Differences, S.-R. and a Possible Personalized Treatment Approach with Valproic Acid: A Review. Biomedicines 2022, 10, 962. [Google Scholar] [CrossRef] [PubMed]
- Mainali, R.; Zabalawi, M.; Long, D.; Buechler, N.; Quillen, E.; Key, C.-C.; Zhu, X.; Parks, J.S.; Furdui, C.; Stacpoole, P.W.; et al. Dichloroacetate reverses sepsis-induced hepatic metabolic dysfunction. Elife 2021, 10, e64611. [Google Scholar] [CrossRef] [PubMed]
- Cavagnaro, J.A. Preclinical safety evaluation of biotechnology-derived pharmaceuticals. Nat. Rev. Drug Discov. 2002, 1, 469–475. [Google Scholar] [CrossRef]
- U.S. Food and Drug Administration. Guideline for the study and evaluation of gender differences in the clinical evaluation of drugs; notice. Fed. Regist. 1993, 58, 39406–39416. [Google Scholar]
- Clayton, J.A.; Collins, F.S. Policy: NIH to balance sex in cell and animal studies. Nature 2014, 509, 282–283. [Google Scholar] [CrossRef]
- de Vries, S.T.; Starokozhko, V.; Schellens, I.M.M.; Wijnans, L.; Enzmann, H.; Cavaleri, M.; Mol, P.G.M. Attention for sex in COVID-19 trials: A review of regulatory dossiers. BMJ Glob. Health 2022, 7, e008173. [Google Scholar] [CrossRef]
- Schoenmann, N.; Tannenbaum, N.; Hodgeman, R.M.; Raju, R.P. Regulating mitochondrial metabolism by targeting pyruvate dehydrogenase with dichloroacetate, a metabolic messenger. Biochim. Biophys. Acta Mol. Basis Dis. 2023, 1869, 166769. [Google Scholar] [CrossRef]
- Nuzzo, E.; Berg, K.M.; Andersen, L.W.; Balkema, J.; Montissol, S.; Cocchi, M.N.; Liu, X.; Donnino, M.W. Pyruvate Dehydrogenase Activity Is Decreased in the Peripheral Blood Mononuclear Cells of Patients with Sepsis. A Prospective Observational Trial. Ann. Am. Thorac. Soc. 2015, 12, 1662–1666. [Google Scholar] [CrossRef]
- Icard, P.; Lincet, H.; Wu, Z.; Coquerel, A.; Forgez, P.; Alifano, M.; Fournel, L. The key role of Warburg effect in SARS-CoV-2 replication and associated inflammatory response. Biochimie 2021, 180, 169–177. [Google Scholar] [CrossRef]
- Yang, K.; Fan, M.; Wang, X.; Xu, J.; Wang, Y.; Tu, F.; Gill, P.S.; Ha, T.; Liu, L.; Williams, D.L.; et al. Lactate promotes macrophage HMGB1 lactylation, acetylation, and exosomal release in polymicrobial sepsis. Cell Death Differ. 2022, 29, 133–146. [Google Scholar] [CrossRef]
- Colegio, O.R.; Chu, N.-Q.; Szabo, A.L.; Chu, T.; Rhebergen, A.M.; Jairam, V.; Cyrus, N.; Brokowski, C.E.; Eisenbarth, S.C.; Phillips, G.M.; et al. Functional polarization of tumour-associated macrophages by tumour-derived lactic acid. Nature 2014, 513, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Gupta, G.S. The Lactate and the Lactate Dehydrogenase in Inflammatory Diseases and Major Risk Factors in COVID-19 Patients. Inflammation 2022, 45, 2091–2123. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- Pitt, B.; Sutton, N.R.; Wang, Z.; Goonewardena, S.N.; Holinstat, M. Potential repurposing of the HDAC inhibitor valproic acid for patients with COVID-19. Eur. J. Pharmacol. 2021, 898, 173988. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, T.; Suzuki, Y.; Imai, J.; Sugano, S.; Hida, M.; Tanigami, A.; Muroi, S.; Yamada, Y.; Hanaoka, K. Molecular cloning, mRNA expression and chromosomal localization of mouse angiotensin-converting enzyme-related carboxypeptidase (mACE2). DNA Seq. 2002, 13, 217–220. [Google Scholar] [CrossRef]
- Tipnis, S.R.; Hooper, N.M.; Hyde, R.; Karran, E.; Christie, G.; Turner, A.J. A human homolog of angiotensin-converting enzyme. Cloning and functional expression as a captopril-insensitive carboxypeptidase. J. Biol. Chem. 2000, 275, 33238–33243. [Google Scholar] [CrossRef]
- Kloc, M.; Ghobrial, R.M.; Kubiak, J.Z. The Role of Genetic Sex and Mitochondria in Response to COVID-19 Infection. Int. Arch. Allergy Immunol. 2020, 181, 629–634. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhao, Z.; Wang, Y.; Zhou, Y.; Ma, Y.; Zuo, W. Single-Cell RNA Expression Profiling of ACE2, the Receptor of SARS-CoV-2. Am. J. Respir. Crit. Care Med. 2020, 202, 756–759. [Google Scholar] [CrossRef]
- Patel, S.K.; Velkoska, E.; Burrell, L.M. Emerging markers in cardiovascular disease: Where does angiotensin-converting enzyme 2 fit in? Clin. Exp. Pharmacol. Physiol. 2013, 40, 551–559. [Google Scholar] [CrossRef]
- Fernández-Atucha, A.; Izagirre, A.; Fraile-Bermúdez, A.B.; Kortajarena, M.; Larrinaga, G.; Martinez-Lage, P.; Echevarría, E.; Gil, J. Sex differences in the aging pattern of renin-angiotensin system serum peptidases. Biol. Sex. Differ. 2017, 8, 5. [Google Scholar] [CrossRef]
- Bukowska, A.; Spiller, L.; Wolke, C.; Lendeckel, U.; Weinert, S.; Hoffmann, J.; Bornfleth, P.; Kutschka, I.; Gardemann, A.; Isermann, B.; et al. Protective regulation of the ACE2/ACE gene expression by estrogen in human atrial tissue from elderly men. Exp. Biol. Med. 2017, 242, 1412–1423. [Google Scholar] [CrossRef]
- Dalpiaz, P.L.M.; Lamas, A.Z.; Caliman, I.F.; Ribeiro, R.F.; Abreu, G.R.; Moyses, M.R.; Andrade, T.U.; Gouvea, S.A.; Alves, M.F.; Carmona, A.K.; et al. Sex Hormones Promote Opposite Effects on ACE and ACE2 Activity, Hypertrophy and Cardiac Contractility in Spontaneously Hypertensive Rats. PLoS ONE 2015, 10, e0127515. [Google Scholar] [CrossRef]
- Fischer, M.; Baessler, A.; Schunkert, H. Renin angiotensin system and gender differences in the cardiovascular system. Cardiovasc. Res. 2002, 53, 672–677. [Google Scholar] [CrossRef] [PubMed]
- Bartachian, M.; McManus, J.M.; Berk, M.P.; Nakamura, F.; Mukhopadhyay, S.; Xu, W.; Erzurum, S.; Drazba, J.; Peterson, J.; Klain, E.A.; et al. Androgen regulation of pulmonary AR, TMPRSS2 and ACE2 with implications for sex-discordant COVID-19 outcomes. Sci. Rep. 2021, 11, 11130. [Google Scholar] [CrossRef] [PubMed]
- Vaarala, M.H.; Porvari, K.S.; Kellokumpu, S.; Kyllönen, A.P.; Vihko, P.T. Expression of transmembrane serine protease TMPRSS2 in mouse and human tissues. J. Pathol. 2001, 193, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Zarubin, A.; Stepanov, V.; Markov, A.; Kolesnikov, N.; Marusin, A.; Khitrinskaya, I.; Swarovskaya, M.; Litvinov, S.; Ekomasova, N.; Dzhaubermezov, M.; et al. Pharmacogenetic Properties of TMPRSS2 Gene as a Potential Target for COVID-19 Therapy. Genes 2020, 12, 19. [Google Scholar] [CrossRef]
- Fortson, W.S.; Kayarthodi, S.; Fujimura, Y.; Xu, H.; Matthews, R.; Grizzle, W.E.; Rao, V.N.; Bhat, G.K.; Reddy, E.S.P. Histone deacetylase inhibitors, valproic acid and trichostatin-A induce apoptosis and affect acetylation status of p53 in ERG-positive prostate cancer cells. Int. J. Oncol. 2011, 39, 111–119. [Google Scholar] [CrossRef]
- Moreno-Pérez, O.; Merino, E.; Ramos, J.M.; Rodríguez, J.C.; Diaz, C.; Mas, P.; Reus, S.; Sánchez-Martínez, R.; Boix, V.; Chico-Sánchez, P.; et al. Valproic Acid Could Help in the Fight Against COVID-19: A case-control study. Neurologia 2022. [Google Scholar] [CrossRef]
- Collazos, J.; Domingo, P.; Fernández-Araujo, N.; Asensi-Díaz, E.; Vilchez-Rueda, H.; Lalueza, A.; Roy-Vallejo, E.; Blanes, R.; Raya-Cruz, M.; Sanz-Cánovas, J.; et al. Valproic Acid in COVID-19 Study Group, Exposure to valproic acid is associated with less pulmonary infiltrates and improvements in diverse clinical outcomes and laboratory parameters in patients hospitalized with COVID-19. PLoS ONE 2022, 17, e0262777. [Google Scholar] [CrossRef]
- Naasani, I.; Analysis, C.O.P.R. A Bioinformatic Approach to Accelerate Drug Repurposing against Covid-19 and Other Emerging Epidemics. SLAS Discov. 2021, 26, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Phiel, C.J.; Zhang, F.; Huang, E.Y.; Guenther, M.G.; Lazar, M.A.; Klein, P.S. Histone deacetylase is a direct target of valproic acid, a potent anticonvulsant, mood stabilizer, and teratogen. J. Biol. Chem. 2001, 276, 36734–36741. [Google Scholar] [CrossRef]
- Maschietto, F.; Qiu, T.; Wang, J.; Shi, Y.; Allen, B.; Lisi, G.P.; Lolis, E.; Batista, V.S. Valproate-coenzyme A conjugate blocks opening of receptor binding domains in the spike trimer of SARS-CoV-2 through an allosteric mechanism. Comput. Struct. Biotechnol. J. 2023, 21, 1066–1076. [Google Scholar] [CrossRef]
- Mengist, H.M.; Fan, X.; Jin, T. Designing of improved drugs for COVID-19: Crystal structure of SARS-CoV-2 main protease Mpro. Signal Transduct. Target. Ther. 2020, 5, 67. [Google Scholar] [CrossRef]
- Elfiky, A.A. Anti-HCV nucleotide inhibitors repurposing against COVID-19. Life Sci. 2020, 248, 117477. [Google Scholar] [CrossRef] [PubMed]
- Elfiky, A.A. Ribavirin; Remdesivir; Sofosbuvir; Galidesivir, and Tenofovir against SARS-CoV-2 RNA dependent RNA polymerase (RdRp): A molecular docking study. Life Sci. 2020, 253, 117592. [Google Scholar] [CrossRef] [PubMed]
- Mamdouh, F.; Mohamed, M.; El-Din, G.; Abuo-Rahma, G.E.-D.; Hayallah, A.M.; Aziz, M.; Nafady, A.; Samir, E. Molecular Docking Study Reveals the Potential Repurposing of Histone Deacetylase Inhibitors Against COVID-19. Int. J. Pharm. Sci. Res. 2020, 11, 4261–4270. [Google Scholar] [CrossRef]
- Jin, Z.; Du, X.; Xu, Y.; Deng, Y.; Liu, M.; Zhao, Y.; Zhang, B.; Li, X.; Zhang, L.; Peng, C.; et al. Structure of Mpro from SARS-CoV-2 and discovery of its inhibitors. Nature 2020, 582, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V.; Hayashi, Y.; Jung, S.-H. An Overview of Severe Acute Respiratory Syndrome-Coronavirus (SARS-CoV) 3CL Protease Inhibitors: Peptidomimetics and Small Molecule Chemotherapy. J. Med. Chem. 2016, 59, 6595–6628. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J. Role of HDAC2 in the pathophysiology of COPD. Annu. Rev. Physiol. 2009, 71, 451–464. [Google Scholar] [CrossRef] [PubMed]
- Dewe, J.M.; Fuller, B.L.; Lentini, J.M.; Kellner, S.M.; Fu, D. TRMT1-Catalyzed tRNA Modifications Are Required for Redox Homeostasis To Ensure Proper Cellular Proliferation and Oxidative Stress Survival. Mol. Cell. Biol. 2017, 37, e00214-17. [Google Scholar] [CrossRef]
- Matsuda, K.I.; Mori, H.; Nugent, B.M.; Pfaff, D.W.; McCarthy, M.M.; Kawata, M. Histone deacetylation during brain development is essential for permanent masculinization of sexual behavior. Endocrinology 2011, 152, 2760–2767. [Google Scholar] [CrossRef] [PubMed]
- Tyler, C.R.S.; Smoake, J.J.W.; Solomon, E.R.; Villicana, E.; Caldwell, K.K.; Allan, A.M. Sex-Dependent Effects of the Histone Deacetylase Inhibitor, Sodium Valproate, on Reversal Learning After Developmental Arsenic Exposure. Front. Genet. 2018, 9, 200. [Google Scholar] [CrossRef]
- Krämer, O.H.; Zhu, P.; Ostendorff, H.P.; Golebiewski, M.; Tiefenbach, J.; Peters, M.A.; Brill, B.; Groner, B.; Bach, I.; Heinzel, T.; et al. The histone deacetylase inhibitor valproic acid selectively induces proteasomal degradation of HDAC2. EMBO J. 2003, 22, 3411–3420. [Google Scholar] [CrossRef]
- Andreu, S.; Ripa, I.; Bello-Morales, R.; López-Guerrero, J.A. Valproic Acid and Its Amidic Derivatives as New Antivirals against Alphaherpesviruses. Viruses 2020, 12, 1356. [Google Scholar] [CrossRef]
- Michaelis, M.; Doerr, H.W.; Cinatl, J. Valproic acid as anti-cancer drug. Curr. Pharm. Des. 2007, 13, 3378–3393. [Google Scholar] [CrossRef]
- Codo, A.C.; Davanzo, G.G.; de Brito Monteiro, L.; de Souza, G.F.; Muraro, S.P.; Virgilio-da-Silva, J.V.; Prodonoff, J.S.; Carregari, V.C.; de Biagi Junior, C.A.O.; Crunfli, F.; et al. Elevated Glucose Levels Favor SARS-CoV-2 Infection and Monocyte Response through a HIF-1α/Glycolysis-Dependent Axis. Cell Metab. 2020, 32, 437–446.e5. [Google Scholar] [CrossRef]
- Sinclair, L.V.; Rolf, J.; Emslie, E.; Shi, Y.-B.; Taylor, P.M.; Cantrell, D.A. Control of amino-acid transport by antigen receptors coordinates the metabolic reprogramming essential for T cell differentiation. Nat. Immunol. 2013, 14, 500–508. [Google Scholar] [CrossRef]
- Kidani, Y.; Elsaesser, H.; Hock, M.B.; Vergnes, L.; Williams, K.J.; Argus, J.P.; Marbois, B.N.; Komisopoulou, E.; Wilson, E.B.; Osborne, T.F.; et al. Sterol regulatory element-binding proteins are essential for the metabolic programming of effector T cells and adaptive immunity. Nat. Immunol. 2013, 14, 489–499. [Google Scholar] [CrossRef]
- Wang, R.; Dillon, C.P.; Shi, L.Z.; Milasta, S.; Carter, R.; Finkelstein, D.; McCormick, L.L.; Fitzgerald, P.; Chi, H.; Munger, J.; et al. The transcription factor Myc controls metabolic reprogramming upon T lymphocyte activation. Immunity 2011, 35, 871–882. [Google Scholar] [CrossRef]
- Khan, S.; Jena, G. Valproic Acid Improves Glucose Homeostasis by Increasing Beta-Cell Proliferation, Function, and Reducing its Apoptosis through HDAC Inhibition in Juvenile Diabetic Rat. J. Biochem. Mol. Toxicol. 2016, 30, 438–446. [Google Scholar] [CrossRef]
- Rakitin, A.; Kõks, S.; Haldre, S. Valproate modulates glucose metabolism in patients with epilepsy after first exposure. Epilepsia 2015, 56, e172–e175. [Google Scholar] [CrossRef]
- Mihaylova, M.M.; Vasquez, D.S.; Ravnskjaer, K.; Denechaud, P.-D.; Yu, R.T.; Alvarez, J.G.; Downes, M.; Evans, R.M.; Montminy, M.; Shaw, R.J. Class IIa histone deacetylases are hormone-activated regulators of FOXO and mammalian glucose homeostasis. Cell 2011, 145, 607–621. [Google Scholar] [CrossRef]
- Thangaraju, M.; Ananth, S.; Martin, P.M.; Roon, P.; Smith, S.B.; Sterneck, E.; Prasad, P.D.; Ganapathy, V. c/ebpdelta Null mouse as a model for the double knock-out of slc5a8 and slc5a12 in kidney. J. Biol. Chem. 2006, 281, 26769–26773. [Google Scholar] [CrossRef]
- Frank, H.; Gröger, N.; Diener, M.; Becker, C.; Braun, T.; Boettger, T. Lactaturia and loss of sodium-dependent lactate uptake in the colon of SLC5A8-deficient mice. J. Biol. Chem. 2008, 283, 24729–24737. [Google Scholar] [CrossRef]
- Singh, N.; Thangaraju, M.; Prasad, P.D.; Martin, P.M.; Lambert, N.A.; Boettger, T.; Offermanns, S.; Ganapathy, V. Blockade of dendritic cell development by bacterial fermentation products butyrate and propionate through a transporter (Slc5a8)-dependent inhibition of histone deacetylases. J. Biol. Chem. 2010, 285, 27601–27608. [Google Scholar] [CrossRef]
- Zimmerman, M.A.; Singh, N.; Martin, P.M.; Thangaraju, M.; Ganapathy, V.; Waller, J.L.; Shi, H.; Robertson, K.D.; Munn, D.H.; Liu, K. Butyrate suppresses colonic inflammation through HDAC1-dependent Fas upregulation and Fas-mediated apoptosis of T cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G1405–G1415. [Google Scholar] [CrossRef]
- Guo, Z.; Zhong, F.; Hou, M.; Xie, J.; Zhang, A.Z.; Li, X.; Li, Y.; Chang, B.; Yang, J. Key enzyme in charge of ketone reabsorption of renal tubular SMCT1 may be a new target in diabetic kidney disease. Nephrol. Dial. Transplant. 2023, gfad173. [Google Scholar] [CrossRef] [PubMed]
- PubChem, SLC5A8—Solute Carrier Family 5 Member 8 (Human). Available online: https://pubchem.ncbi.nlm.nih.gov/gene/SLC5A8/human (accessed on 10 November 2023).
- Damanskienė, E.; Balnytė, I.; Valančiūtė, A.; Lesauskaitė, V.; Alonso, M.M.; Stakišaitis, D. The Comparative Experimental Study of Sodium and Magnesium Dichloroacetate Effects on Pediatric PBT24 and SF8628 Cell Glioblastoma Tumors Using a Chicken Embryo Chorioallantoic Membrane Model and on Cells In Vitro. Int. J. Mol. Sci. 2022, 23, 10455. [Google Scholar] [CrossRef]
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Cunningham, F.; Allen, J.E.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Austine-Orimoloye, O.; Azov, A.G.; Barnes, I.; Bennett, R.; et al. Ensembl 2022. Nucleic Acids Res. 2022, 50, D988–D995. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 15–21. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022, 50, gkac194. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. ClusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Sodhi, C.P.; Wohlford-Lenane, C.; Yamaguchi, Y.; Prindle, T.; Fulton, W.B.; Wang, S.; McCray, P.B.; Chappell, M.; Hackam, D.J.; Jia, H. Attenuation of pulmonary ACE2 activity impairs inactivation of des-Arg9 bradykinin/BKB1R axis and facilitates LPS-induced neutrophil infiltration. Am. J. Physiol. Lung Cell. Mol. Physiol. 2018, 314, L17–L31. [Google Scholar] [CrossRef]
- Chen, L.Y.C.; Hoiland, R.L.; Stukas, S.; Wellington, C.L.; Sekhon, M.S. Confronting the controversy: Interleukin-6; the COVID-19 cytokine storm syndrome. Eur. Respir. J. 2020, 56, 2003006. [Google Scholar] [CrossRef]
- Pantazi, I.; Al-Qahtani, A.A.; Alhamlan, F.S.; Alothaid, H.; Matou-Nasri, S.; Sourvinos, G.; Vergadi, E.; Tsatsanis, C. SARS-CoV-2/ACE2 Interaction Suppresses IRAK-M Expression and Promotes Pro-Inflammatory Cytokine Production in Macrophages. Front. Immunol. 2021, 12, 683800. [Google Scholar] [CrossRef]
- Chiu, K.H.-Y.; Yip, C.C.-Y.; Poon, R.W.-S.; Leung, K.-H.; Li, X.; Hung, I.F.-N.; To, K.K.-W.; Cheng, V.C.-C.; Yuen, K.-Y. Correlations of Myeloperoxidase (MPO), Adenosine deaminase (ADA), C-C motif chemokine 22 (CCL22), Tumour necrosis factor alpha (TNFα) and Interleukin-6 (IL-6) mRNA expression in the nasopharyngeal specimens with the diagnosis and severity of SARS-CoV-2 infections. Emerg. Microbes Infect. 2023, 12, 2157338. [Google Scholar] [CrossRef]
- Russell, C.D.; Valanciute, A.; Gachanja, N.N.; Stephen, J.; Penrice-Randal, R.; Armstrong, S.D.; Clohisey, S.; Wang, B.; Al Qsous, W.; Wallace, W.A.; et al. Tissue Proteomic Analysis Identifies Mechanisms and Stages of Immunopathology in Fatal COVID-19. Am. J. Respir. Cell Mol. Biol. 2022, 66, 196–205. [Google Scholar] [CrossRef]
- Arunachalam, P.S.; Wimmers, F.; Mok, C.K.P.; Perera, R.A.P.M.; Scott, M.; Hagan, T.; Sigal, N.; Feng, Y.; Bristow, L.; Tsang, O.T.-Y.; et al. Systems biological assessment of immunity to mild versus severe COVID-19 infection in humans. Science 2020, 369, 1210–1220. [Google Scholar] [CrossRef]
- Jorens, P.G.; De Jongh, R.; Bossaert, L.L.; De Backer, W.; Herman, A.G.; Pollet, H.; Bosmans, E.; Taupin, J.L.; Moreau, J.F. High levels of leukaemia inhibitory factor in ARDS. Cytokine 1996, 8, 873–876. [Google Scholar] [CrossRef]
- Chehboun, S.; Labrecque-Carbonneau, J.; Pasquin, S.; Meliani, Y.; Meddah, B.; Ferlin, W.; Sharma, M.; Tormo, A.; Masson, J.-F.; Gauchat, J.-F. Epstein-Barr virus-induced gene. J. Biol. Chem. 2017, 292, 6644–6656. [Google Scholar] [CrossRef]
- Attur, M.; Petrilli, C.; Adhikari, S.; Iturrate, E.; Li, X.; Tuminello, S.; Hu, N.; Chakravarti, A.; Beck, D.; Abramson, S.B. Interleukin-1 receptor antagonist gene ( IL1RN ) variants modulate the cytokine release syndrome and mortality of SARS-CoV-2. medRxiv 2023. preprint. [Google Scholar] [CrossRef]
- Allan, S.M.; Tyrrell, P.J.; Rothwell, N.J. Interleukin-1; neuronal injury. Nat. Rev. Immunol. 2005, 5, 629–640. [Google Scholar] [CrossRef]
- Tamayo-Velasco, Á.; Martínez-Paz, P.; Peñarrubia-Ponce, M.J.; de la Fuente, I.; Pérez-González, S.; Fernández, I.; Dueñas, C.; Gómez-Sánchez, E.; Lorenzo-López, M.; Gómez-Pesquera, E.; et al. IL-1α, and IL-27 Are Robust Biomarkers in Early Severity Stratification of COVID-19 Patients. J. Clin. Med. 2021, 10, 2017. [Google Scholar] [CrossRef]
- Liu, Y.; Li, S.; Zhang, G.; Nie, G.; Meng, Z.; Mao, D.; Chen, C.; Chen, X.; Zhou, B.; Zeng, G. Genetic variants in IL1A and IL1B contribute to the susceptibility to 2009 pandemic H1N1 influenza A virus. BMC Immunol 2013, 14, 37. [Google Scholar] [CrossRef]
- Rajamanickam, A.; Nathella, P.K.; Selvaraj, N.; Manoj, M.; Thangaraj, J.W.V.; Muthusamy, S.K.; Purushothaman, G.K.C.; Bhatnagar, T.; Ponnaiah, M.; Ramasamy, S.; et al. Characterization of IL-10 Family of Cytokines in Acute and Convalescent COVID-19 Individuals. J. Interferon Cytokine Res. 2023, 43, 469–477. [Google Scholar] [CrossRef]
- Huang, F.; Wachi, S.; Thai, P.; Loukoianov, A.; Tan, K.H.; Forteza, R.M.; Wu, R. Potentiation of IL-19 expression in airway epithelia by IL-17A and IL-4/IL-13: Important implications in asthma. J. Allergy Clin. Immunol. 2008, 121, 1415–1421.e3. [Google Scholar] [CrossRef]
- Sharif-Askari, F.S.; Sharif-Askari, N.S.; Hafezi, S.; Goel, S.; Alsayed, H.A.H.; Ansari, A.W.; Mahboub, B.; Al-Muhsen, S.; Temsah, M.-H.; Hamid, Q.; et al. Upregulation of interleukin-19 in saliva of patients with COVID-19. Sci. Rep. 2022, 12, 16019. [Google Scholar] [CrossRef]
- Zamani, B.; Momen-Heravi, M.; Erami, M.; Motedayyen, H.; ArefNezhad, R. Impacts of IL-27 and IL-32 in the pathogenesis and outcome of COVID-19 associated mucormycosis. J. Immunoassay Immunochem. 2023, 44, 242–255. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef]
- Yang, X.; Yu, Y.; Xu, J.; Shu, H.; Xia, J.; Liu, H.; Wu, Y.; Zhang, L.; Yu, Z.; Fang, M.; et al. Clinical course and outcomes of critically ill patients with SARS-CoV-2 pneumonia in Wuhan, China: A single-centered, retrospective, observational study. Lancet Respir. Med. 2020, 8, 475–481. [Google Scholar] [CrossRef]
- Meidaninikjeh, S.; Sabouni, N.; Marzouni, H.Z.; Bengar, S.; Khalili, A.; Jafari, R. Monocytes and macrophages in COVID-19: Friends and foes. Life Sci. 2021, 269, 119010. [Google Scholar] [CrossRef]
- Nunnari, G.; Sanfilippo, C.; Castrogiovanni, P.; Imbesi, R.; Volti, G.L.; Barbagallo, I.; Musumeci, G.; Di Rosa, M. Network perturbation analysis in human bronchial epithelial cells following SARS-CoV2 infection. Exp. Cell Res. 2020, 395, 112204. [Google Scholar] [CrossRef]
- Khalil, B.A.; Elemam, N.M.; Maghazachi, A.A. Chemokines and chemokine receptors during COVID-19 infection. Comput. Struct. Biotechnol. J. 2021, 19, 976–988. [Google Scholar] [CrossRef]
- Torri, A.; Beretta, O.; Ranghetti, A.; Granucci, F.; Ricciardi-Castagnoli, P.; Foti, M. Gene expression profiles identify inflammatory signatures in dendritic cells. PLoS ONE 2010, 5, e9404. [Google Scholar] [CrossRef]
- Baker, J.R.; Mahdi, M.; Nicolau, D.V.; Ramakrishnan, S.; Barnes, P.J.; Simpson, J.L.; Cass, S.P.; Russell, R.E.K.; Donnelly, L.E.; Bafadhel, M. Early Th2 inflammation in the upper respiratory mucosa as a predictor of severe COVID-19 and modulation by early treatment with inhaled corticosteroids: A mechanistic analysis. Lancet Respir. Med. 2022, 10, 545–556. [Google Scholar] [CrossRef]
- Li, Y.; Han, X. Bioinformatics Approaches to Identify the Comorbidity Complexities of SARS-CoV-2 Infection with Crohn’s Disease. J. Coll. Physicians Surg. Pak. 2023, 33, 1093–1099. [Google Scholar] [CrossRef]
- Jang, Y.; Seo, S.H. Gene expression pattern differences in primary human pulmonary epithelial cells infected with MERS-CoV or SARS-CoV-2. Arch Virol. 2020, 165, 2205–2211. [Google Scholar] [CrossRef]
- Liang, Y.; Li, H.; Li, J.; Yang, Z.-N.; Li, J.-L.; Zheng, H.-W.; Chen, Y.-L.; Shi, H.-J.; Guo, L.; Liu, L.-D. Role of neutrophil chemoattractant CXCL5 in SARS-CoV-2 infection-induced lung inflammatory innate immune response in an in vivo hACE2 transfection mouse model. Zool. Res. 2020, 41, 621–631. [Google Scholar] [CrossRef]
- Sims, J.T.; Krishnan, V.; Chang, C.-Y.; Engle, S.M.; Casalini, G.; Rodgers, G.H.; Bivi, N.; Nickoloff, B.J.; Konrad, R.J.; de Bono, S.; et al. Characterization of the cytokine storm reflects hyperinflammatory endothelial dysfunction in COVID-19. J. Allergy Clin. Immunol. 2021, 147, 107–111. [Google Scholar] [CrossRef]
- Perreau, M.; Suffiotti, M.; Marques-Vidal, P.; Wiedemann, A.; Levy, Y.; Laouénan, C.; Ghosn, J.; Fenwick, C.; Comte, D.; Roger, T.; et al. The cytokines HGF and CXCL13 predict the severity and the mortality in COVID-19 patients. Nat. Commun. 2021, 12, 4888. [Google Scholar] [CrossRef]
- López-Ayllón, B.D.; de Lucas-Rius, A.; Mendoza-García, L.; García-García, T.; Fernández-Rodríguez, R.; Suárez-Cárdenas, J.M.; Santos, F.M.; Corrales, F.; Redondo, N.; Pedrucci, F.; et al. SARS-CoV-2 accessory proteins involvement in inflammatory and profibrotic processes through IL11 signaling. Front. Immunol. 2023, 14, 1220306. [Google Scholar] [CrossRef]
- Oppmann, B.; Lesley, R.; Blom, B.; Timans, J.C.; Xu, Y.; Hunte, B.; Vega, F.; Yu, N.; Wang, J.; Singh, K.; et al. Novel p19 protein engages IL-12p40 to form a cytokine, IL-23, with biological activities similar as well as distinct from IL-12. Immunity 2000, 13, 715–725. [Google Scholar] [CrossRef]
- Nelson, R.T.; Boyd, J.; Gladue, R.P.; Paradis, T.; Thomas, R.; Cunningham, A.C.; Lira, P.; Brissette, W.H.; Hayes, L.; Hames, L.M.; et al. Genomic organization of the CC chemokine mip-3alpha/CCL20/larc/exodus/SCYA20, showing gene structure, splice variants, and chromosome localization. Genomics 2001, 73, 28–37. [Google Scholar] [CrossRef]
- Hue, S.; Beldi-Ferchiou, A.; Bendib, I.; Surenaud, M.; Fourati, S.; Frapard, T.; Rivoal, S.; Razazi, K.; Carteaux, G.; Delfau-Larue, M.-H.; et al. Uncontrolled Innate and Impaired Adaptive Immune Responses in Patients with COVID-19 Acute Respiratory Distress Syndrome. Am. J. Respir. Crit. Care Med. 2020, 202, 1509–1519. [Google Scholar] [CrossRef]
- Alberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. Helper T Cells and Lymphocyte Activation. In Molecular Biology of the Cell, 4th ed.; Garland Science: New York, NY, USA, 2002; Available online: https://www.ncbi.nlm.nih.gov/books/NBK26827/ (accessed on 20 November 2023).
- Jiyarom, B.; Giannakopoulos, S.; Strange, D.P.; Panova, N.; Gale, M.; Verma, S. RIG-I and MDA5 are modulated by bone morphogenetic protein (BMP6) and are essential for restricting Zika virus infection in human Sertoli cells. Front. Microbiol. 2022, 13, 1062499. [Google Scholar] [CrossRef]
- Eddowes, L.A.; Al-Hourani, K.; Ramamurthy, N.; Frankish, J.; Baddock, H.T.; Sandor, C.; Ryan, J.D.; Fusco, D.N.; Arezes, J.; Giannoulatou, E.; et al. Antiviral activity of bone morphogenetic proteins and activins. Nat. Microbiol. 2019, 4, 339–351. [Google Scholar] [CrossRef]
- Miller, D.S.J.; Hill, C.S.; Signaling, F.; Bradshaw, R.A.; Hart, G.W.; Stahl, P.D. Encyclopedia of Cell Biology, 2nd ed.; Academic Press: Oxford, UK, 2023; pp. 46–61. [Google Scholar] [CrossRef]
- Zovi, A.; Ferrara, F.; Langella, R.; Cavallaro, F.; Vitiello, A. Sex affects immune response capacity against COVID-19 infection. Rev. Med. Virol. 2023, 33, e2450. [Google Scholar] [CrossRef]
- GB Atypical Chemokine Receptors ACKR3 & ACKR5. Luxembourg Institute of Health, Luxembourg Institute of Health (n.d.). Available online: https://www.lih.lu/en/event/atypical-chemokine-receptors-ackr3-ackr5/ (accessed on 21 November 2023).
- Zhou, Z.; Ren, L.; Zhang, L.; Zhong, J.; Xiao, Y.; Jia, Z.; Guo, L.; Yang, J.; Wang, C.; Jiang, S.; et al. Heightened Innate Immune Responses in the Respiratory Tract of COVID-19 Patients. Cell Host Microbe 2020, 27, 883–890.e2. [Google Scholar] [CrossRef]
- Schioppa, T.; Sozio, F.; Barbazza, I.; Scutera, S.; Bosisio, D.; Sozzani, S.; Del Prete, A. Molecular Basis for CCRL2 Regulation of Leukocyte Migration. Front. Cell Dev. Biol. 2020, 8, 615031. [Google Scholar] [CrossRef]
- Galligan, C.L.; Matsuyama, W.; Matsukawa, A.; Mizuta, H.; Hodge, D.R.; Howard, O.M.Z.; Yoshimura, T. Up-regulated expression and activation of the orphan chemokine receptor, CCRL2, in rheumatoid arthritis. Arthritis Rheum. 2004, 50, 1806–1814. [Google Scholar] [CrossRef]
- Mannes, P.Z.; Barnes, C.E.; Biermann, J.; Latoche, J.D.; Day, K.E.; Zhu, Q.; Tabary, M.; Xiong, Z.; Nedrow, J.R.; Izar, B.; et al. Molecular imaging of chemokine-like receptor 1 (CMKLR1) in experimental acute lung injury. Proc. Natl. Acad. Sci. USA 2023, 120, e2216458120. [Google Scholar] [CrossRef] [PubMed]
- Liechti, T.; Iftikhar, Y.; Mangino, M.; Beddall, M.; Goss, C.W.; O’Halloran, J.A.; Mudd, P.A.; Roederer, M. Immune phenotypes that are associated with subsequent COVID-19 severity inferred from post-recovery samples. Nat. Commun. 2022, 13, 7255. [Google Scholar] [CrossRef] [PubMed]
- Blot, M.; Jacquier, M.; Glele, L.-S.A.; Beltramo, G.; Nguyen, M.; Bonniaud, P.; Prin, S.; Andreu, P.; Bouhemad, B.; Bour, J.-B.; et al. Pneumochondrie study group, CXCL10 could drive longer duration of mechanical ventilation during COVID-19 ARDS. Crit. Care 2020, 24, 632. [Google Scholar] [CrossRef]
- Liao, M.; Liu, Y.; Yuan, J.; Wen, Y.; Xu, G.; Zhao, J.; Cheng, L.; Li, J.; Wang, X.; Wang, F.; et al. Single-cell landscape of bronchoalveolar immune cells in patients with COVID-19. Nat. Med. 2020, 26, 842–844. [Google Scholar] [CrossRef]
- Chua, R.L.; Lukassen, S.; Trump, S.; Hennig, B.P.; Wendisch, D.; Pott, F.; Debnath, O.; Thürmann, L.; Kurth, F.; Völker, M.T.; et al. COVID-19 severity correlates with airway epithelium-immune cell interactions identified by single-cell analysis. Nat. Biotechnol. 2020, 38, 970–979. [Google Scholar] [CrossRef]
- Darbeheshti, F.; Mahdiannasser, M.; Uhal, B.D.; Ogino, S.; Gupta, S.; Rezaei, N. Interindividual immunogenic variants: Susceptibility to coronavirus, respiratory syncytial virus and influenza virus. Rev. Med. Virol. 2021, 31, e2234. [Google Scholar] [CrossRef]
- Miller, H.E.; Robinson, R.T. Early control of Mycobacterium tuberculosis infection requires il12rb1 expression by rag1-dependent lineages. Infect. Immun. 2012, 80, 3828–3841. [Google Scholar] [CrossRef]
- Fujii, H. Cell type-specific roles of Jak3 in IL-2-induced proliferative signal transduction. Biochem. Biophys. Res. Commun. 2007, 354, 825–829. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ward-Kavanagh, L.K.; Lin, W.W.; Šedý, J.R.; Ware, C.F. The TNF Receptor Superfamily in Co-stimulating and Co-inhibitory Responses. Immunity 2016, 44, 1005–1019. [Google Scholar] [CrossRef] [PubMed]
- Anisul, M.; Shilts, J.; Schwartzentruber, J.; Hayhurst, J.; Buniello, A.; Mohammed, E.S.E.; Zheng, J.; Holmes, M.; Ochoa, D.; Carmona, M.; et al. A proteome-wide genetic investigation identifies several SARS-CoV-2-exploited host targets of clinical relevance. Elife 2021, 10, e69719. [Google Scholar] [CrossRef]
- Moro-García, M.A.; Mayo, J.C.; Sainz, R.M.; Alonso-Arias, R. Influence of Inflammation in the Process of T Lymphocyte Differentiation: Proliferative, Metabolic, and Oxidative Changes. Front. Immunol. 2018, 9, 339. [Google Scholar] [CrossRef] [PubMed]
- Day, M. Covid-19: Ibuprofen should not be used for managing symptoms, say doctors and scientists. BMJ 2020, 368, m1086. [Google Scholar] [CrossRef] [PubMed]
- Itagaki, S.; Gopal, E.; Zhuang, L.; Fei, Y.-J.; Miyauchi, S.; Prasad, P.D.; Ganapathy, V. Interaction of ibuprofen and other structurally related NSAIDs with the sodium-coupled monocarboxylate transporter SMCT1 (SLC5A8). Pharm. Res. 2006, 23, 1209–1216. [Google Scholar] [CrossRef] [PubMed]
- Zmudzinski, M.; Rut, W.; Olech, K.; Granda, J.; Giurg, M.; Kaleta, R.; Zgarbova, M.; Kasprzyk, R.; Zhang, L.; Sun, X.; et al. Ebselen derivatives inhibit SARS-CoV-2 replication by inhibition of its essential proteins: PLpro and Mpro proteases, and nsp14 guanine N7-methyltransferase. Sci. Rep. 2023, 13, 1–16. [Google Scholar] [CrossRef]
- Mello, M.L.S. Sodium Valproate-Induced Chromatin Remodeling. Front. Cell. Dev. Biol. 2021, 9, 645518. [Google Scholar] [CrossRef]
- Stakišaitis, D.; Juknevičienė, M.; Damanskienė, E.; Valančiūtė, A.; Balnytė, I.; Alonso, M.M. The Importance of Gender-Related Anticancer Research on Mitochondrial Regulator Sodium Dichloroacetate in Preclinical Studies In Vivo. Cancers 2019, 11, 1210. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Log2 Fold Change | Gene Expression Change | Log2 Count Average of Control Cells | Log2 Count Average of Treated Cells | p Value |
|---|---|---|---|---|---|
| ACKR3 | −2.61 | ↓ | 7.78 | 6.67 | 3.87 × 10−5 |
| CCL22 | −3.10 | ↓ | 8.21 | 6.78 | 2.20 × 10−5 |
| CCL24 | −4.61 | ↓ | 11.70 | 8.63 | 3.32 × 10−5 |
| CCL4 | −1.92 | ↓ | 14.76 | 14.08 | 1.42 × 10−4 |
| CCR1 | −2.10 | ↓ | 12.41 | 10.88 | 8.55 × 10−4 |
| CCR2 | −1.80 | ↓ | 12.94 | 11.78 | 4.87 × 10−2 |
| CCRL2 | −2.45 | ↓ | 9.41 | 8.09 | 3.93 × 10−5 |
| CD209 | −2.85 | ↓ | 7.09 | 4.84 | 9.47 × 10−3 |
| CMKLR1 | −1.92 | ↓ | 9.09 | 8.09 | 6.53 × 10−3 |
| CSF1 | −3.34 | ↓ | 9.71 | 8.22 | 1.09 × 10−6 |
| CSF2 | −3.73 | ↓ | 7.00 | 4.91 | 1.04 × 10−4 |
| CSF3 | −2.98 | ↓ | 6.64 | 6.08 | 4.14 × 10−2 |
| CSF3R | −2.24 | ↓ | 12.90 | 11.30 | 1.09 × 10−3 |
| CX3CR1 | −2.27 | ↓ | 10.19 | 8.42 | 1.21 × 10−3 |
| CXCL1 | −6.29 | ↓ | 13.00 | 8.51 | 1.15 × 10−12 |
| CXCL13 | −3.51 | ↓ | 4.34 | 3.34 | 5.36 × 10−3 |
| CXCL2 | −3.98 | ↓ | 14.08 | 11.91 | 5.70 × 10−6 |
| CXCL3 | −4.52 | ↓ | 10.63 | 7.74 | 8.09 × 10−6 |
| CXCL5 | −7.23 | ↓ | 15.28 | 9.89 | 1.75 × 10−13 |
| CXCL6 | −2.75 | ↓ | 7.43 | 6.23 | 1.28 × 10−2 |
| CXCR2 | −1.81 | ↓ | 10.43 | 9.06 | 2.36 × 10−2 |
| CXCR3 | −1.91 | ↓ | 8.81 | 7.96 | 7.93 × 10−4 |
| EBI3 | −2.60 | ↓ | 5.75 | 4.89 | 5.28 × 10−4 |
| IL10 | −2.90 | ↓ | 7.88 | 6.81 | 5.50 × 10−5 |
| IL12RB1 | −1.85 | ↓ | 8.69 | 7.66 | 3.36 × 10−4 |
| IL19 | −5.40 | ↓ | 6.69 | 3.84 | 2.16 × 10−5 |
| IL1A | −4.14 | ↓ | 11.66 | 9.43 | 1.10 × 10−4 |
| IL1B | −2.98 | ↓ | 16.01 | 14.87 | 1.30 × 10−3 |
| IL1RN | −4.21 | ↓ | 12.46 | 9.74 | 3.63 × 10−9 |
| IL23A | −2.14 | ↓ | 9.45 | 9.33 | 4.72 × 10−3 |
| IL24 | −2.87 | ↓ | 10.20 | 9.16 | 1.42 × 10−2 |
| IL27 | −2.53 | ↓ | 5.98 | 4.25 | 3.72 × 10−3 |
| IL27RA | −1.89 | ↓ | 9.49 | 8.48 | 1.08 × 10−3 |
| IL2RA | −1.85 | ↓ | 9.60 | 9.21 | 7.22 × 10−3 |
| IL6 | −6.29 | ↓ | 11.31 | 7.22 | 5.84 × 10−8 |
| ITGB2 | −2.08 | ↓ | 14.09 | 12.86 | 6.84 × 10−7 |
| JAK3 | −1.88 | ↓ | 10.76 | 9.77 | 1.69 × 10−5 |
| LIF | −4.53 | ↓ | 7.88 | 4.76 | 1.64 × 10−4 |
| OSM | −3.21 | ↓ | 8.58 | 6.38 | 6.38 × 10−6 |
| TNFRSF14 | −1.91 | ↓ | 13.19 | 12.40 | 1.05 × 10−7 |
| Pathway ID | Number of Genes | Enrichment Score | NES | p Value | Treatment Has a Significant Effect on the Expression of a Gene |
|---|---|---|---|---|---|
| GO:0005125: Cytokine activity | 22 | −0.82 | −2.73 | 1.00 × 10−10 | Decreased: CCL22, CCL24, CSF1, CSF2, CSF3, CXCL1, CXCL2, CXCL3, CXCL5, CXCL6, CXCL13, EBI3, IL1A, IL1B, IL1RN, IL6, IL10, IL19, IL24, IL27, LIF, OSM |
| GO:0070098: Chemokine-mediated signaling pathway | 17 | −0.68 | −2.08 | 6.10 × 10−5 | Decreased: ACKR3, CCL4, CCL22, CCL24, CCR1, CCR2, CCRL2, CMKLR1, CX3CR1, CXCL1, CXCL2, CXCL3, CXCL5, CXCL6, CXCL13, CXCR2, CXCR3 |
| GO:0030593: Neutrophil chemotaxis | 11 | −0.63 | −1.86 | 1.50 × 10−3 | Decreased: CCL4, CCL22, CCL24, CSF3R, CXCL1, CXCL2, CXCL3, CXCL5, CXCL6, CXCL13, ITGB2 |
| GO:0048247: Lymphocyte chemotaxis | 5 | −0.68 | −1.53 | 3.60 × 10−2 | Decreased: CCL4, CCL22, CCL24, CXCL13, CXCR3 |
| GO:0042098: Regulation of T-cell proliferation | 14 | −0.47 | −1.52 | 4.10 × 10−2 | Decreased: CCR2, CD209, EBI3, IL1A, IL1B, IL2RA, IL6, IL10, IL12RB1, IL23A, IL27, IL27RA, JAK3, TNFRSF14 |
| Gene | Log2 Fold Change | Gene Expression Change | Log2 Count Average of Control Cells | Log2 Count Average of Treated Cells | p Value |
|---|---|---|---|---|---|
| ACKR3 | −1.79 | ↓ | 8.29 | 6.42 | 7.97 × 10−4 |
| BMP6 | 2.02 | ↑ | 8.31 | 10.51 | 4.11 × 10−5 |
| CCL2 | −3.12 | ↓ | 15.12 | 10.06 | 4.32 × 10−5 |
| CCL20 | 2.53 | ↑ | 11.32 | 13.39 | 1.85 × 10−3 |
| CCL24 | −3.04 | ↓ | 14.04 | 9.32 | 7.81 × 10−4 |
| CCL7 | −6.00 | ↓ | 12.10 | 5.33 | 1.18 × 10−12 |
| CCR1 | −2.67 | ↓ | 13.62 | 10.25 | 2.20 × 10−7 |
| CCR3 | −1.71 | ↓ | 7.47 | 5.23 | 1.94 × 10−2 |
| CSF3 | 3.10 | ↑ | 6.58 | 9.35 | 9.30 × 10−3 |
| CSF3R | −2.15 | ↓ | 13.24 | 10.89 | 1.33 × 10−4 |
| CXCL1 | −3.36 | ↓ | 13.29 | 9.97 | 3.55 × 10−6 |
| CXCL2 | 1.51 | ↑ | 12.31 | 13.66 | 3.45 × 10−2 |
| CXCL5 | −4.22 | ↓ | 15.61 | 10.21 | 1.37 × 10−7 |
| CXCL6 | −2.06 | ↓ | 9.39 | 7.24 | 2.42 × 10−2 |
| CXCL8 | 1.77 | ↑ | 16.81 | 18.50 | 1.76 × 10−2 |
| CXCR2 | −2.46 | ↓ | 11.26 | 8.86 | 2.10 × 10−4 |
| IL11 | 2.70 | ↑ | 0.76 | 3.54 | 1.07 × 10−3 |
| IL1A | 2.85 | ↑ | 9.39 | 11.86 | 1.14 × 10−3 |
| IL1B | 2.17 | ↑ | 14.61 | 16.62 | 3.98 × 10−3 |
| IL23A | 3.30 | ↑ | 9.09 | 11.97 | 9.80 × 10−8 |
| IL24 | 2.55 | ↑ | 9.12 | 11.02 | 7.66 × 10−3 |
| Pathway ID | Number of Genes | Enrichment Score | NES | p Value | Significant Effect of Treatment on Gene Expression |
|---|---|---|---|---|---|
| GO:0005125: Cytokine activity | 11 | 0.41 | 1.74 | 0.015 | Decreased: CSF3 Increased: BMP6, CCL20, CXCL2, CXCL8, IL11, IL1A, IL1B, IL23A, IL24, NODAL |
| GO:0070098: Chemokine-mediated signaling pathway | 10 | −0.61 | −1.89 | 0.025 | Decreased: ACKR3, CCL2, CCL24, CCL7, CCR1, CCR3, CXCL1, CXCL5, CXCL6, CXCR2 |
| GO:0030593: Neutrophil chemotaxis | 8 | −0.66 | −1.97 | 0.001 | Decreased: CCL2, CCL24, CCL7, CSF3R, CXCL1, CXCL5, CXCL6, CXCR2 |
| GO:0048247: Lymphocyte chemotaxis | 3 | −0.72 | −1.55 | 0.04 | Decreased: CCL2, CCL7, CCL24 |
| GO:0042098: T-cell proliferation | No effect of treatment on genes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stakišaitis, D.; Kapočius, L.; Tatarūnas, V.; Gečys, D.; Mickienė, A.; Tamošuitis, T.; Ugenskienė, R.; Vaitkevičius, A.; Balnytė, I.; Lesauskaitė, V. Effects of Combined Treatment with Sodium Dichloroacetate and Sodium Valproate on the Genes in Inflammation- and Immune-Related Pathways in T Lymphocytes from Patients with SARS-CoV-2 Infection with Pneumonia: Sex-Related Differences. Pharmaceutics 2024, 16, 409. https://doi.org/10.3390/pharmaceutics16030409
Stakišaitis D, Kapočius L, Tatarūnas V, Gečys D, Mickienė A, Tamošuitis T, Ugenskienė R, Vaitkevičius A, Balnytė I, Lesauskaitė V. Effects of Combined Treatment with Sodium Dichloroacetate and Sodium Valproate on the Genes in Inflammation- and Immune-Related Pathways in T Lymphocytes from Patients with SARS-CoV-2 Infection with Pneumonia: Sex-Related Differences. Pharmaceutics. 2024; 16(3):409. https://doi.org/10.3390/pharmaceutics16030409
Chicago/Turabian StyleStakišaitis, Donatas, Linas Kapočius, Vacis Tatarūnas, Dovydas Gečys, Auksė Mickienė, Tomas Tamošuitis, Rasa Ugenskienė, Arūnas Vaitkevičius, Ingrida Balnytė, and Vaiva Lesauskaitė. 2024. "Effects of Combined Treatment with Sodium Dichloroacetate and Sodium Valproate on the Genes in Inflammation- and Immune-Related Pathways in T Lymphocytes from Patients with SARS-CoV-2 Infection with Pneumonia: Sex-Related Differences" Pharmaceutics 16, no. 3: 409. https://doi.org/10.3390/pharmaceutics16030409
APA StyleStakišaitis, D., Kapočius, L., Tatarūnas, V., Gečys, D., Mickienė, A., Tamošuitis, T., Ugenskienė, R., Vaitkevičius, A., Balnytė, I., & Lesauskaitė, V. (2024). Effects of Combined Treatment with Sodium Dichloroacetate and Sodium Valproate on the Genes in Inflammation- and Immune-Related Pathways in T Lymphocytes from Patients with SARS-CoV-2 Infection with Pneumonia: Sex-Related Differences. Pharmaceutics, 16(3), 409. https://doi.org/10.3390/pharmaceutics16030409