

The Intra-Articular Delivery of a Low-Dose Adeno-Associated Virus-IL-1 Receptor Antagonist Vector Alleviates the Progress of Arthritis in an Osteoarthritis Rat Model

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Viral Vector Production
2.3. Animals
2.4. In Vivo Bioluminescence Imaging
2.5. Rat OA Model
2.6. X-Ray Clinical Score
- 0: A normal state, with no observed narrowing of the joint space and the absence of reactive bone changes on the X-ray;
- I: There is a possibility of mild narrowing of the joint space and initial, minor osteophyte formation;
- II: The definite presence of small osteophytes and potential narrowing of the joint space, as evidenced on the X-ray;
- III: Characterized by multiple, moderate-sized osteophytes, definitive narrowing of the joint space, some degree of subchondral bone sclerosis, and a likelihood of bony deformity within the joint;
- IV: A severe state with numerous large osteophytes, marked narrowing of the joint space, pronounced subchondral bone sclerosis, and overt bony deformity of the joint.
2.7. Micro-CT Examination
2.8. ELISA Assay
2.9. Statistical Analysis
3. Results
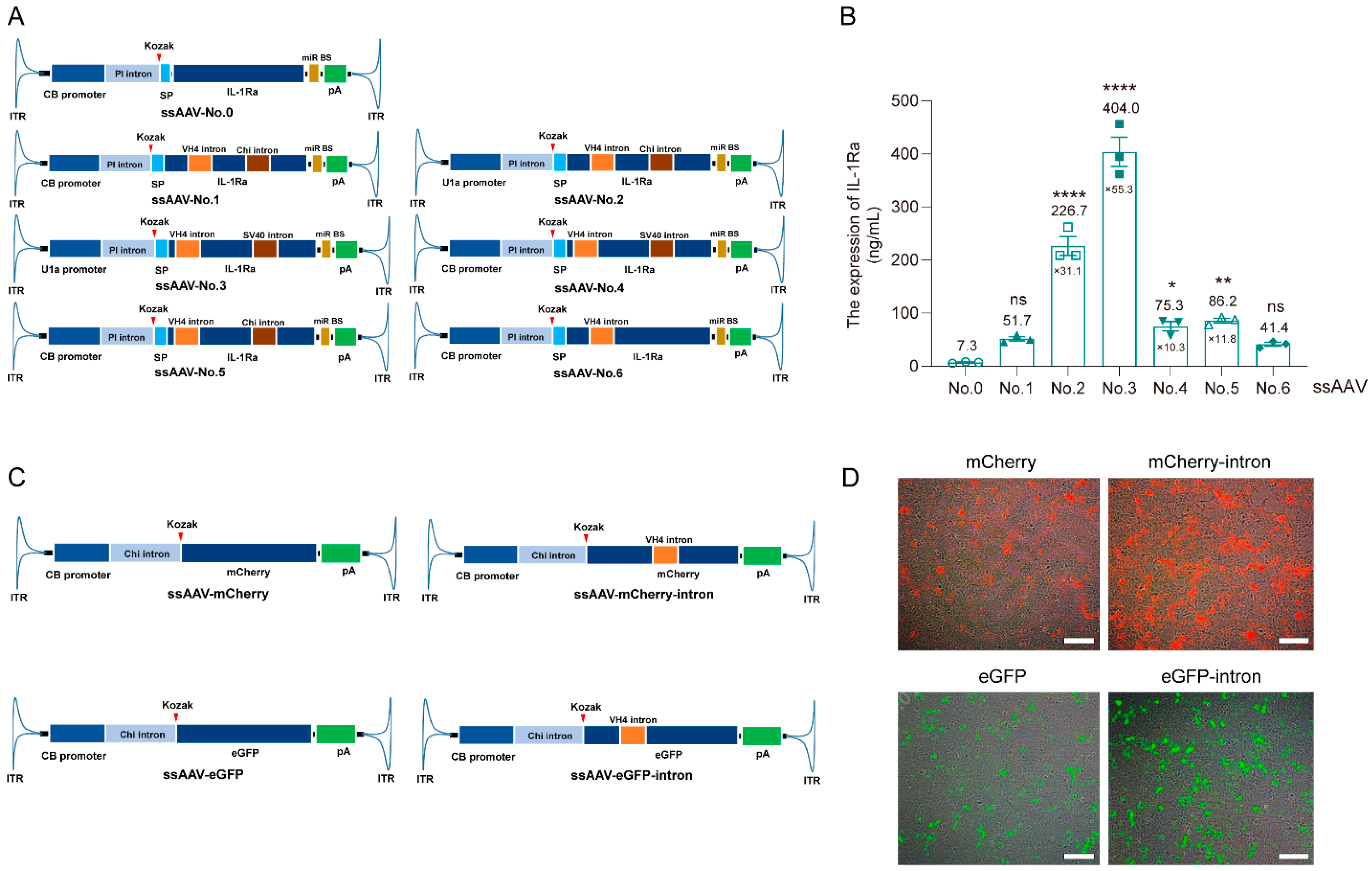
3.1. Optimize the Expression Cassette of IL-1Ra Plasmid Using Intron Insertion into CDS Technology
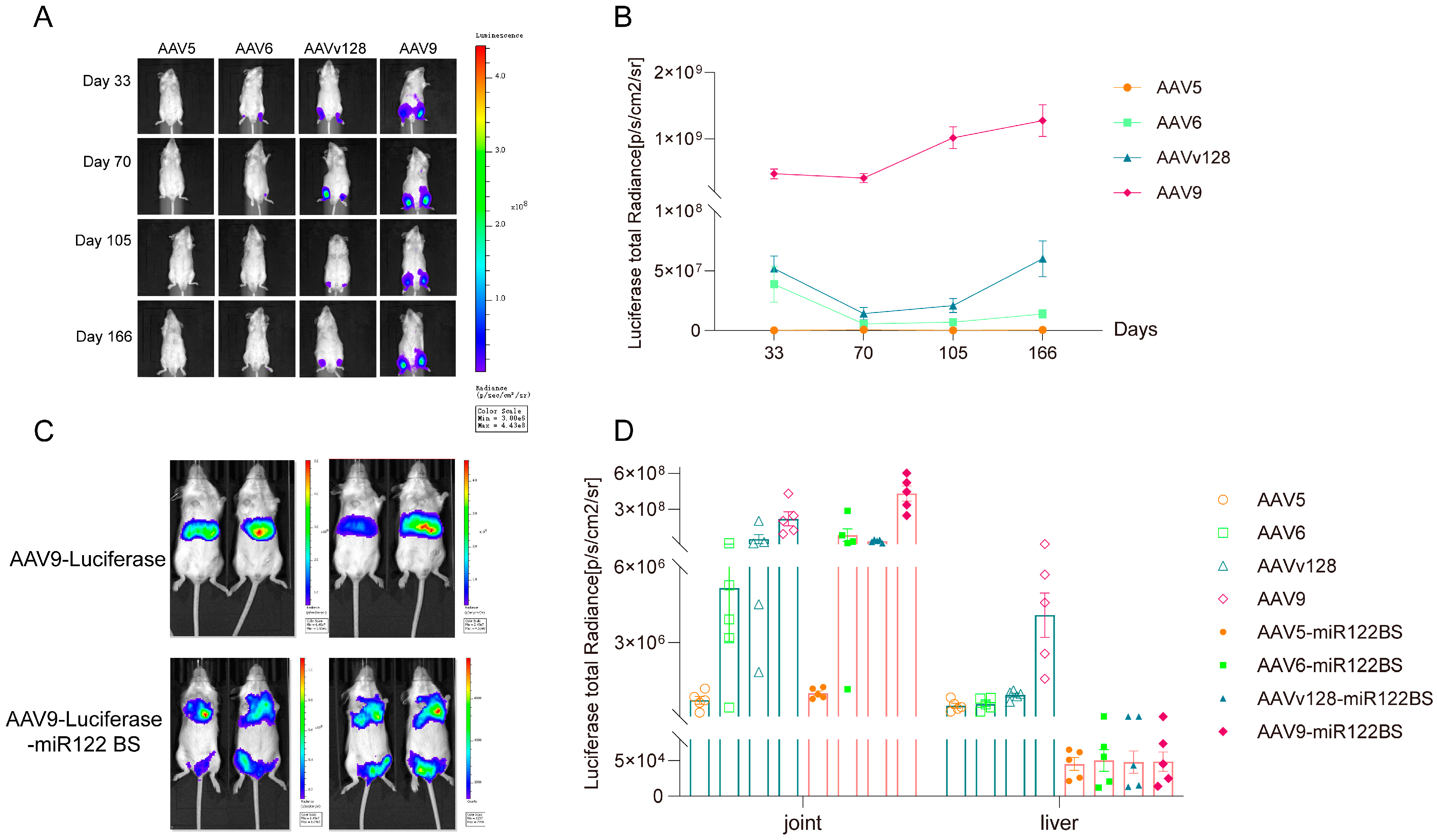
3.2. Intra-Articular Injection of AAV9-Luciferase-miR122 BS Vector Significantly Reduces the Expression Level in Liver
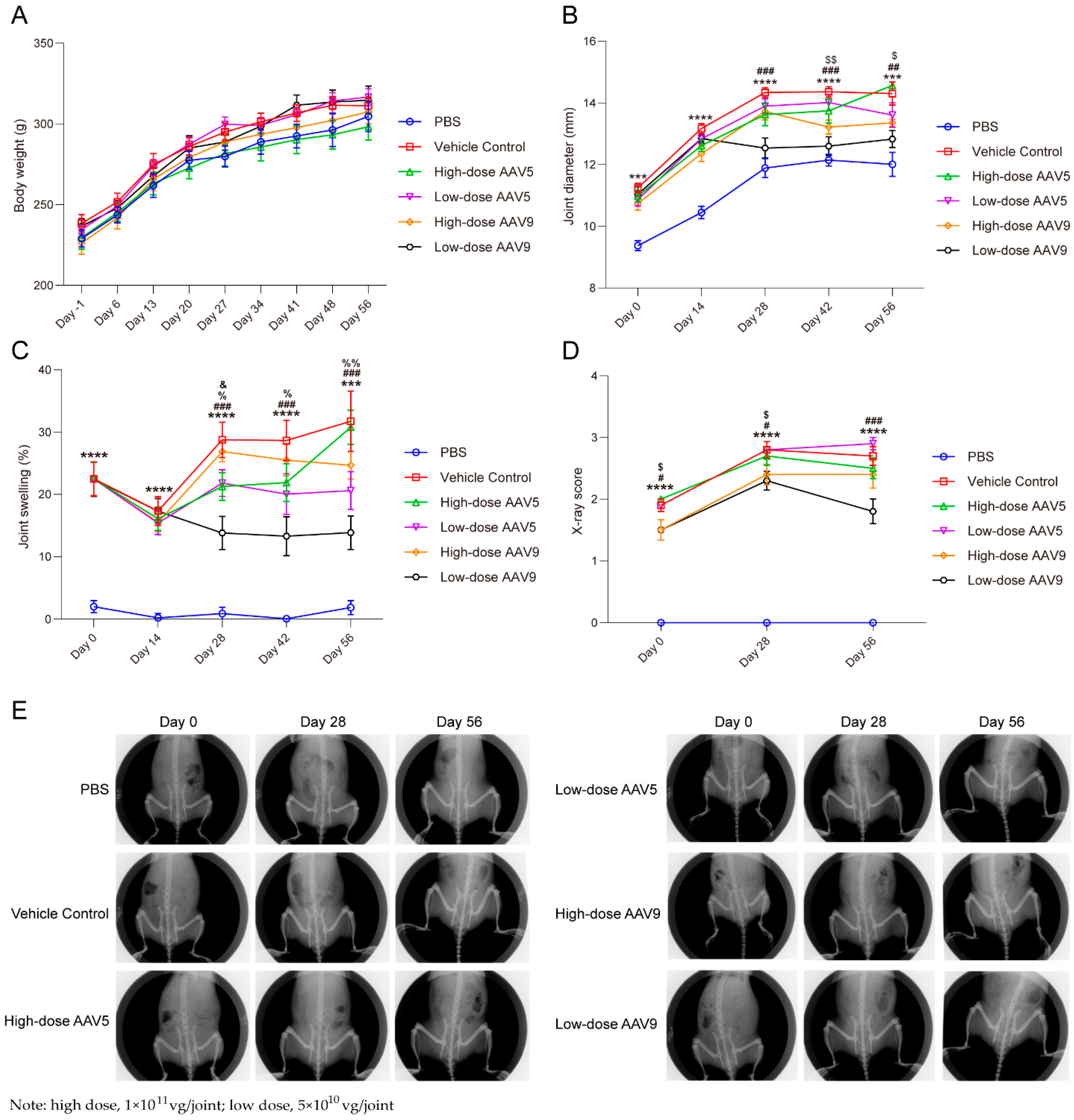
3.3. Low-Dose AAV Has Good Efficacy on OA Model in Rats, While High-Dose AAV Has Poor Therapeutic Effect
3.4. AAV-IL-1Ra Gene Therapy Substantially Protects Trabecular Bone-Related Indicators in Animal Models
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- GBD 2019 Diseases and Injuries Collaborators. Global burden of 369 diseases and injuries in 204 countries and territories, 1990–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1204–1222. [Google Scholar] [CrossRef] [PubMed]
- Im, G.-I.; Kim, T.-K. Regenerative Therapy for Osteoarthritis: A Perspective. Int. J. Stem Cells 2020, 13, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Grässel, S.; Muschter, D. Recent advances in the treatment of osteoarthritis. F1000Research 2020, 9, 325. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Jeong, S.; Kim, H.; Kang, D.; Lee, J.; Kang, S.-B.; Kim, J.-H. Disease-modifying therapeutic strategies in osteoarthritis: Current status and future directions. Exp. Mol. Med. 2021, 53, 1689–1696. [Google Scholar] [CrossRef] [PubMed]
- Martel-Pelletier, J.; Barr, A.J.; Cicuttini, F.M.; Conaghan, P.G.; Cooper, C.; Goldring, M.B.; Goldring, S.R.; Jones, G.; Teichtahl, A.J.; Pelletier, J.-P. Osteoarthritis. Nat. Rev. Dis. Prim. 2016, 2, 16072. [Google Scholar] [CrossRef]
- Urban, H.; Little, C.B. The role of fat and inflammation in the pathogenesis and management of osteoarthritis. Rheumatology 2018, 57, iv10–iv21. [Google Scholar] [CrossRef]
- Arend, W.P.; Malyak, M.; Guthridge, C.J.; Gabay, C. Interleukin-1 receptor antagonist: Role in biology. Annu. Rev. Immunol. 1998, 16, 27–55. [Google Scholar] [CrossRef]
- Chevalier, X.; Goupille, P.; Beaulieu, A.D.; Burch, F.X.; Bensen, W.G.; Conrozier, T.; Loeuille, D.; Kivitz, A.J.; Silver, D.; Appleton, B.E. Intraarticular injection of anakinra in osteoarthritis of the knee: A multicenter, randomized, double-blind, placebo-controlled study. Arthritis Care Res. 2009, 61, 344–352. [Google Scholar] [CrossRef]
- Evans, C.H.; Kraus, V.B.; Setton, L.A. Progress in intra-articular therapy. Nat. Rev. Rheumatol. 2013, 10, 11–22. [Google Scholar] [CrossRef]
- Wang, D.; Tai, P.W.L.; Gao, G. Adeno-associated virus vector as a platform for gene therapy delivery. Nat. Rev. Drug Discov. 2019, 18, 358–378. [Google Scholar] [CrossRef]
- Li, C.; Samulski, R.J. Engineering adeno-associated virus vectors for gene therapy. Nat. Rev. Genet. 2020, 21, 255–272. [Google Scholar] [CrossRef] [PubMed]
- Ke, X.; Jiang, H.; Li, Q.; Luo, S.; Qin, Y.; Li, J.; Xie, Q.; Zheng, Q. Preclinical evaluation of KH631, a novel rAAV8 gene therapy product for neovascular age-related macular degeneration. Mol. Ther. 2023, 31, 3308–3321. [Google Scholar] [CrossRef]
- Levings, R.S.W.; Smith, A.D.; Broome, T.A.; Rice, B.L.; Gibbs, E.P.; Myara, D.A.; Hyddmark, E.V.; Nasri, E.; Zarezadeh, A.; Levings, P.P.; et al. Self-Complementary Adeno-Associated Virus-Mediated Interleukin-1 Receptor Antagonist Gene Delivery for the Treatment of Osteoarthritis: Test of Efficacy in an Equine Model. Hum. Gene Ther. Clin. Dev. 2018, 29, 101–112. [Google Scholar] [CrossRef]
- Shen, W.; Liu, S.; Ou, L. rAAV immunogenicity, toxicity, and durability in 255 clinical trials: A meta-analysis. Front. Immunol. 2022, 13, 1001263. [Google Scholar] [CrossRef] [PubMed]
- Qiao, C.; Yuan, Z.; Li, J.; He, B.; Zheng, H.; Mayer, C.; Xiao, X. Liver-specific microRNA-122 target sequences incorporated in AAV vectors efficiently inhibits transgene expression in the liver. Gene Ther. 2011, 18, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, Z.; Chen, G.; Chen, Z.; Guo, C.; Ji, X.; Cui, Q.; Sun, Y.; Hu, X.; Zheng, Y.; et al. MicroRNA-122-Mediated Liver Detargeting Enhances the Tissue Specificity of Cardiac Genome Editing. Circulation 2024, 149, 1778–1781. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.N.; Sha, S.; Hong, M.S.; Maloney, A.J.; Barone, P.W.; Neufeld, C.; Wolfrum, J.; Springs, S.L.; Sinskey, A.J.; Braatz, R.D. Mechanistic model for production of recombinant adeno-associated virus via triple transfection of HEK293 cells. Mol. Ther.-Methods Clin. Dev. 2021, 21, 642–655. [Google Scholar] [CrossRef]
- Brand, D.D.; Latham, K.A.; Rosloniec, E.F. Collagen-induced arthritis. Nat. Protoc. 2007, 2, 1269–1275. [Google Scholar] [CrossRef]
- Cuzzocrea, S.; Mazzon, E.; Dugo, L.; Serraino, I.; Britti, D.; De Maio, M.; Caputi, A.P. Absence of endogeneous interleukin-10 enhances the evolution of murine type-II collagen-induced arthritis. Eur. Cytokine Netw. 2001, 12, 568–580. [Google Scholar]
- Ke, X.; Xie, Q.; Luo, S.; Li, Q.; Zheng, Q.; Zhang, Z. Intra-Articular Delivery of an AAV-Anti-TNF-α Vector Alleviates the Progress of Arthritis in a RA Mouse Model. Hum. Gene Ther. 2024, 35, 754–766. [Google Scholar] [CrossRef]
- Luo, S.; Jiang, H.; Li, Q.; Qin, Y.; Yang, S.; Li, J.; Xu, L.; Gou, Y.; Zhang, Y.; Liu, F.; et al. An adeno-associated virus variant enabling efficient ocular-directed gene delivery across species. Nat. Commun. 2024, 15, 3780. [Google Scholar] [CrossRef] [PubMed]
- Shaul, O. How introns enhance gene expression. Int. J. Biochem. Cell Biol. 2017, 91, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Lacy-Hulbert, A.; Thomas, R.; Li, X.-P.; E Lilley, C.; Coffin, R.S.; Roes, J. Interruption of coding sequences by heterologous introns can enhance the functional expression of recombinant genes. Gene Ther. 2001, 8, 649–653. [Google Scholar] [CrossRef] [PubMed]
- Baier, T.; Jacobebbinghaus, N.; Einhaus, A.; Lauersen, K.J.; Kruse, O. Introns mediate post-transcriptional enhancement of nuclear gene expression in the green microalga Chlamydomonas reinhardtii. PLoS Genet. 2020, 16, e1008944. [Google Scholar] [CrossRef]
- Rose, A.B.; Carter, A.; Korf, I.; Kojima, N. Intron sequences that stimulate gene expression in Arabidopsis. Plant Mol. Biol. 2016, 92, 337–346. [Google Scholar] [CrossRef]
- Broderick, L.; Hoffman, H.M. IL-1 and autoinflammatory disease: Biology, pathogenesis and therapeutic targeting. Nat. Rev. Rheumatol. 2022, 18, 448–463. [Google Scholar] [CrossRef]
- Filipowicz, W.; Grosshans, H. The liver-specific microRNA miR-122: Biology and therapeutic potential. Prog. Drug Res. 2011, 67, 221–238. [Google Scholar]
- Girard, M.; Jacquemin, E.; Munnich, A.; Lyonnet, S.; Henrion-Caude, A. miR-122, a paradigm for the role of microRNAs in the liver. J. Hepatol. 2008, 48, 648–656. [Google Scholar] [CrossRef]
- Jopling, C. Liver-specific microRNA-122: Biogenesis and function. RNA Biol. 2012, 9, 137–142. [Google Scholar] [CrossRef]
- Valdmanis, P.N.; Kim, H.K.; Chu, K.; Zhang, F.; Xu, J.; Munding, E.M.; Shen, J.; Kay, M.A. miR-122 removal in the liver activates imprinted microRNAs and enables more effective microRNA-mediated gene repression. Nat. Commun. 2018, 9, 5321. [Google Scholar] [CrossRef]
- Lu, H.; Qu, G.; Yang, X.; Xu, R.; Xiao, W. Systemic elimination of de novo capsid protein synthesis from replication-competent AAV contamination in the liver. Hum. Gene Ther. 2011, 22, 625–632. [Google Scholar] [CrossRef]
- Bandiera, S.; Pfeffer, S.; Baumert, T.F.; Zeisel, M.B. miR-122—A key factor and therapeutic target in liver disease. J. Hepatol. 2015, 62, 448–457. [Google Scholar] [CrossRef]
- Lee, J.; Mou, H.; Ibraheim, R.; Liang, S.-Q.; Liu, P.; Xue, W.; Sontheimer, E.J. Tissue-restricted genome editing in vivo specified by microRNA-repressible anti-CRISPR proteins. RNA 2019, 25, 1421–1431. [Google Scholar] [CrossRef]
- Yang, Y.-S.; Lin, C.; Ma, H.; Xie, J.; Kaplan, F.S.; Gao, G.; Shim, J.-H. AAV-Mediated Targeting of the Activin A-ACVR1(R206H) Signaling in Fibrodysplasia Ossificans Progressiva. Biomolecules 2023, 13, 1364. [Google Scholar] [CrossRef]
- Yin, L.; Keeler, G.D.; Zhang, Y.; Hoffman, B.E.; Ling, C.; Qing, K.; Srivastava, A. AAV3-miRNA vectors for growth suppression of human hepatocellular carcinoma cells in vitro and human liver tumors in a murine xenograft model in vivo. Gene Ther. 2021, 28, 422–434. [Google Scholar] [CrossRef]
- Xie, J.; Burt, D.R.; Gao, G. Adeno-associated virus-mediated microRNA delivery and therapeutics. Semin. Liver Dis. 2015, 35, 81–88. [Google Scholar] [CrossRef]
- Dismuke, D.J.; Tenenbaum, L.; Samulski, R.J. Biosafety of recombinant adeno-associated virus vectors. Curr. Gene Ther. 2014, 13, 434–452. [Google Scholar] [CrossRef]
- Ling, Q.; Herstine, J.A.; Bradbury, A.; Gray, S.J. AAV-based in vivo gene therapy for neurological disorders. Nat. Rev. Drug Discov. 2023, 22, 789–806. [Google Scholar] [CrossRef]
- Campochiaro, P.A.; Avery, R.; Brown, D.M.; Heier, J.S.; Ho, A.C.; Huddleston, S.M.; Jaffe, G.J.; Khanani, A.M.; Pakola, S.; Pieramici, D.J.; et al. Gene therapy for neovascular age-related macular degeneration by subretinal delivery of RGX-314: A phase 1/2a dose-escalation study. Lancet 2024, 403, 1563–1573. [Google Scholar] [CrossRef]
- Wang, J.-H.; Gessler, D.J.; Zhan, W.; Gallagher, T.L.; Gao, G. Adeno-associated virus as a delivery vector for gene therapy of human diseases. Signal Transduct. Target. Ther. 2024, 9, 78. [Google Scholar] [CrossRef]
- Gerwin, N.; Bendele, A.; Glasson, S.; Carlson, C. The OARSI histopathology initiative—Recommendations for histological assessments of osteoarthritis in the rat. Osteoarthr. Cartil. 2010, 18 (Suppl. S3), S24–S34. [Google Scholar] [CrossRef] [PubMed]
- Thysen, S.; Luyten, F.P.; Lories, R.J.U. Targets, models and challenges in osteoarthritis research. Dis. Model. Mech. 2015, 8, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Luo, S.; Zhang, Y.; Ou, Y.; Wang, H.; Guo, Z.; He, C.; Bai, S.; He, P.; Jiang, M.; et al. Single-shot AAV-vectored vaccine against SARS-CoV-2 with fast and long-lasting immunity. Acta Pharm. Sin. B 2022, 13, 2219–2233. [Google Scholar] [CrossRef] [PubMed]
- Wu, I.; Zeng, A.; Greer-Short, A.; Aycinena, J.A.; Tefera, A.E.; Shenwai, R.; Farshidfar, F.; Van Pell, M.; Xu, E.; Reid, C.; et al. AAV9:PKP2 improves heart function and survival in a Pkp2-deficient mouse model of arrhythmogenic right ventricular cardiomyopathy. Commun. Med. 2024, 4, 38. [Google Scholar] [CrossRef]
- Schulz, M.; Levy, D.; Petropoulos, C.J.; Bashirians, G.; Winburn, I.; Mahn, M.; Somanathan, S.; Cheng, S.H.; Byrne, B.J. Binding and neutralizing anti-AAV antibodies: Detection and implications for rAAV-mediated gene therapy. Mol. Ther. 2023, 31, 616–630. [Google Scholar] [CrossRef]
- Wang, G.; Evans, C.H.; Benson, J.M.; Hutt, J.A.; Seagrave, J.; Wilder, J.A.; Grieger, J.C.; Samulski, R.J.; Terse, P.S. Safety and biodistribution assessment of sc-rAAV2.5IL-1Ra administered via intra-articular injection in a mono-iodoacetate-induced osteoarthritis rat model. Mol. Ther.-Methods Clin. Dev. 2016, 3, 15052. [Google Scholar] [CrossRef]
- Levings, R.S.W.; Broome, T.A.; Smith, A.D.; Rice, B.L.; Gibbs, E.P.; Myara, D.A.; Hyddmark, E.V.; Nasri, E.; Zarezadeh, A.; Levings, P.P.; et al. Gene Therapy for Osteoarthritis: Pharmacokinetics of Intra-Articular Self-Complementary Adeno-Associated Virus Interleukin-1 Receptor Antagonist Delivery in an Equine Model. Hum. Gene Ther. Clin. Dev. 2018, 29, 90–100. [Google Scholar] [CrossRef]
- Xie, J.; Zhang, D.; Lin, Y.; Yuan, Q.; Zhou, X. Anterior Cruciate Ligament Transection-Induced Cellular and Extracellular Events in Menisci: Implications for Osteoarthritis. Am. J. Sports Med. 2018, 46, 1185–1198. [Google Scholar] [CrossRef]
- Martino, A.T.; Suzuki, M.; Markusic, D.M.; Zolotukhin, I.; Ryals, R.C.; Moghimi, B.; Ertl, H.C.J.; Muruve, D.A.; Lee, B.; Herzog, R.W. The genome of self-complementary adeno-associated viral vectors increases Toll-like receptor 9-dependent innate immune responses in the liver. Blood 2011, 117, 6459–6468. [Google Scholar] [CrossRef]
- Ashley, S.N.; Somanathan, S.; Giles, A.R.; Wilson, J.M. TLR9 signaling mediates adaptive immunity following systemic AAV gene therapy. Cell Immunol. 2019, 346, 103997. [Google Scholar] [CrossRef]
- Zhu, J.; Huang, X.; Yang, Y. The TLR9-MyD88 pathway is critical for adaptive immune responses to adeno-associated virus gene therapy vectors in mice. J. Clin. Investig. 2009, 119, 2388–2398. [Google Scholar] [CrossRef] [PubMed]
- Muhuri, M.; Zhan, W.; Maeda, Y.; Li, J.; Lotun, A.; Chen, J.; Sylvia, K.; Dasgupta, I.; Arjomandnejad, M.; Nixon, T.; et al. Novel Combinatorial MicroRNA-Binding Sites in AAV Vectors Synergistically Diminish Antigen Presentation and Transgene Immunity for Efficient and Stable Transduction. Front. Immunol. 2021, 12, 674242. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Muhuri, M.; Li, S.; Qin, W.; Xu, G.; Luo, L.; Li, J.; Letizia, A.J.; Wang, S.K.; Chan, Y.K.; et al. Circumventing cellular immunity by miR142-mediated regulation sufficiently supports rAAV-delivered OVA expression without activating humoral immunity. J. Clin. Investig. 2019, 4, 99052. [Google Scholar] [CrossRef] [PubMed]
- Ertl, H.C.J. T Cell-Mediated Immune Responses to AAV and AAV Vectors. Front. Immunol. 2021, 12, 666666. [Google Scholar] [CrossRef]
- Ertl, H.C.J. Immunogenicity and toxicity of AAV gene therapy. Front. Immunol. 2022, 13, 975803. [Google Scholar] [CrossRef]
- Chand, D.H.; Zaidman, C.; Arya, K.; Millner, R.; Farrar, M.A.; Mackie, F.E.; Goedeker, N.L.; Dharnidharka, V.R.; Dandamudi, R.; Reyna, S.P. Thrombotic Microangiopathy Following Onasemnogene Abeparvovec for Spinal Muscular Atrophy: A Case Series. J. Pediatr. 2021, 231, 265–268. [Google Scholar] [CrossRef]
- Mullard, A. Gene therapy community grapples with toxicity issues, as pipeline matures. Nat. Rev. Drug Discov. 2021, 20, 804–805. [Google Scholar] [CrossRef]
- Hunter, D.J.; Guermazi, A.; Roemer, F.; Zhang, Y.; Neogi, T. Structural correlates of pain in joints with osteoarthritis. Osteoarthr. Cartil. 2013, 21, 1170–1178. [Google Scholar] [CrossRef]
- Lafeber, F.P.; van Spil, W.E. Osteoarthritis year 2013 in review: Biomarkers; reflecting before moving forward, one step at a time. Osteoarthr. Cartil. 2013, 21, 1452–1464. [Google Scholar] [CrossRef]
- Li, X.; Shen, L.; Deng, Z.; Huang, Z. New treatment for osteoarthritis: Gene therapy. Precis. Clin. Med. 2023, 6, pbad014. [Google Scholar] [CrossRef]
- Zhang, X.; Mao, Z.; Yu, C. Suppression of early experimental osteoarthritis by gene transfer of interleukin-1 receptor antagonist and interleukin-10. J. Orthop. Res. 2004, 22, 742–750. [Google Scholar] [CrossRef] [PubMed]
- Stone, A.; Grol, M.W.; Ruan, M.Z.C.; Dawson, B.; Chen, Y.; Jiang, M.-M.; Song, I.-W.; Jayaram, P.; Cela, R.; Gannon, F.; et al. Combinatorial Prg4 and Il-1ra Gene Therapy Protects Against Hyperalgesia and Cartilage Degeneration in Post-Traumatic Osteoarthritis. Hum. Gene Ther. 2019, 30, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Karuppal, R. Current concepts in the articular cartilage repair and regeneration. J. Orthop. 2017, 14, A1–A3. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, S.; Jiang, H.; Li, Q.; Yang, S.; Yu, X.; Xu, X.; Xie, Q.; Ke, X.; Zheng, Q. The Intra-Articular Delivery of a Low-Dose Adeno-Associated Virus-IL-1 Receptor Antagonist Vector Alleviates the Progress of Arthritis in an Osteoarthritis Rat Model. Pharmaceutics 2024, 16, 1518. https://doi.org/10.3390/pharmaceutics16121518
Luo S, Jiang H, Li Q, Yang S, Yu X, Xu X, Xie Q, Ke X, Zheng Q. The Intra-Articular Delivery of a Low-Dose Adeno-Associated Virus-IL-1 Receptor Antagonist Vector Alleviates the Progress of Arthritis in an Osteoarthritis Rat Model. Pharmaceutics. 2024; 16(12):1518. https://doi.org/10.3390/pharmaceutics16121518
Chicago/Turabian StyleLuo, Shuang, Hao Jiang, Qingwei Li, Shiping Yang, Xuemei Yu, Xiongliang Xu, Qing Xie, Xiao Ke, and Qiang Zheng. 2024. "The Intra-Articular Delivery of a Low-Dose Adeno-Associated Virus-IL-1 Receptor Antagonist Vector Alleviates the Progress of Arthritis in an Osteoarthritis Rat Model" Pharmaceutics 16, no. 12: 1518. https://doi.org/10.3390/pharmaceutics16121518
APA StyleLuo, S., Jiang, H., Li, Q., Yang, S., Yu, X., Xu, X., Xie, Q., Ke, X., & Zheng, Q. (2024). The Intra-Articular Delivery of a Low-Dose Adeno-Associated Virus-IL-1 Receptor Antagonist Vector Alleviates the Progress of Arthritis in an Osteoarthritis Rat Model. Pharmaceutics, 16(12), 1518. https://doi.org/10.3390/pharmaceutics16121518