
The Remodulation of Actin Bundles during the Stimulation of Mitochondria in Adult Human Fibroblasts in Response to Light




Abstract
:
1. Introduction
2. Materials and Methods
2.1. Preparation of Cell Cultures
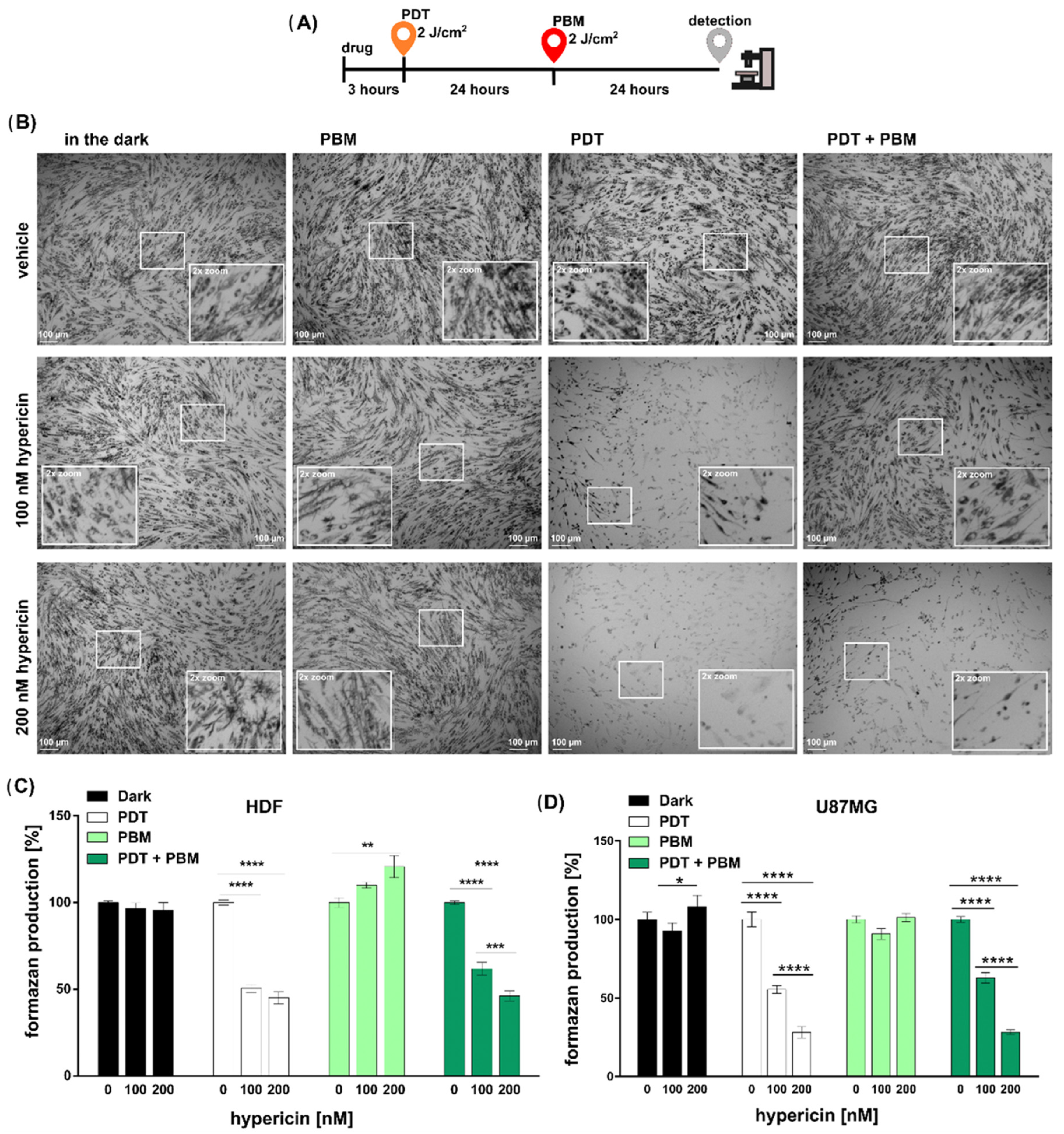
2.2. Therapeutical Protocols
2.3. Cell Metabolism/Viability Assay
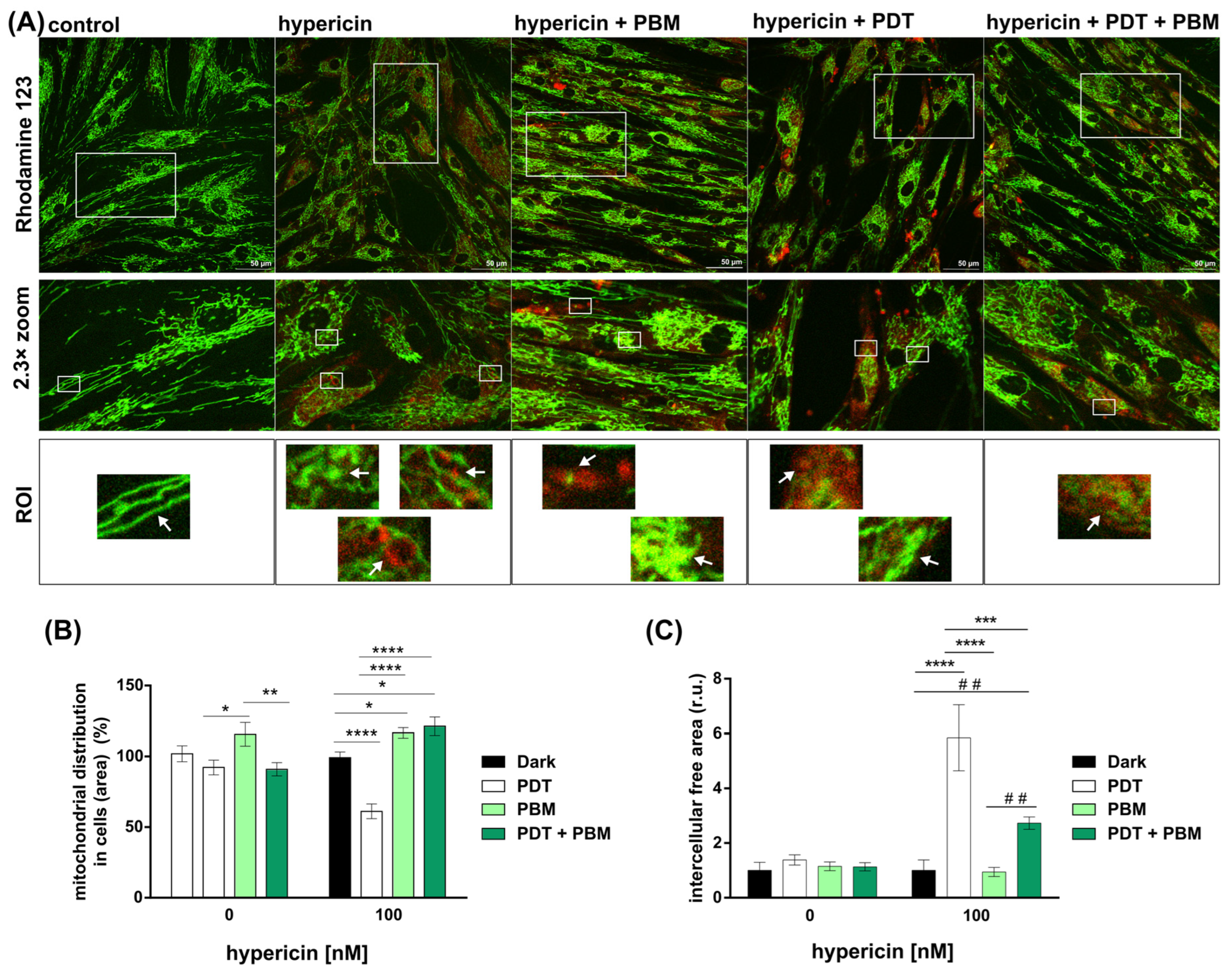
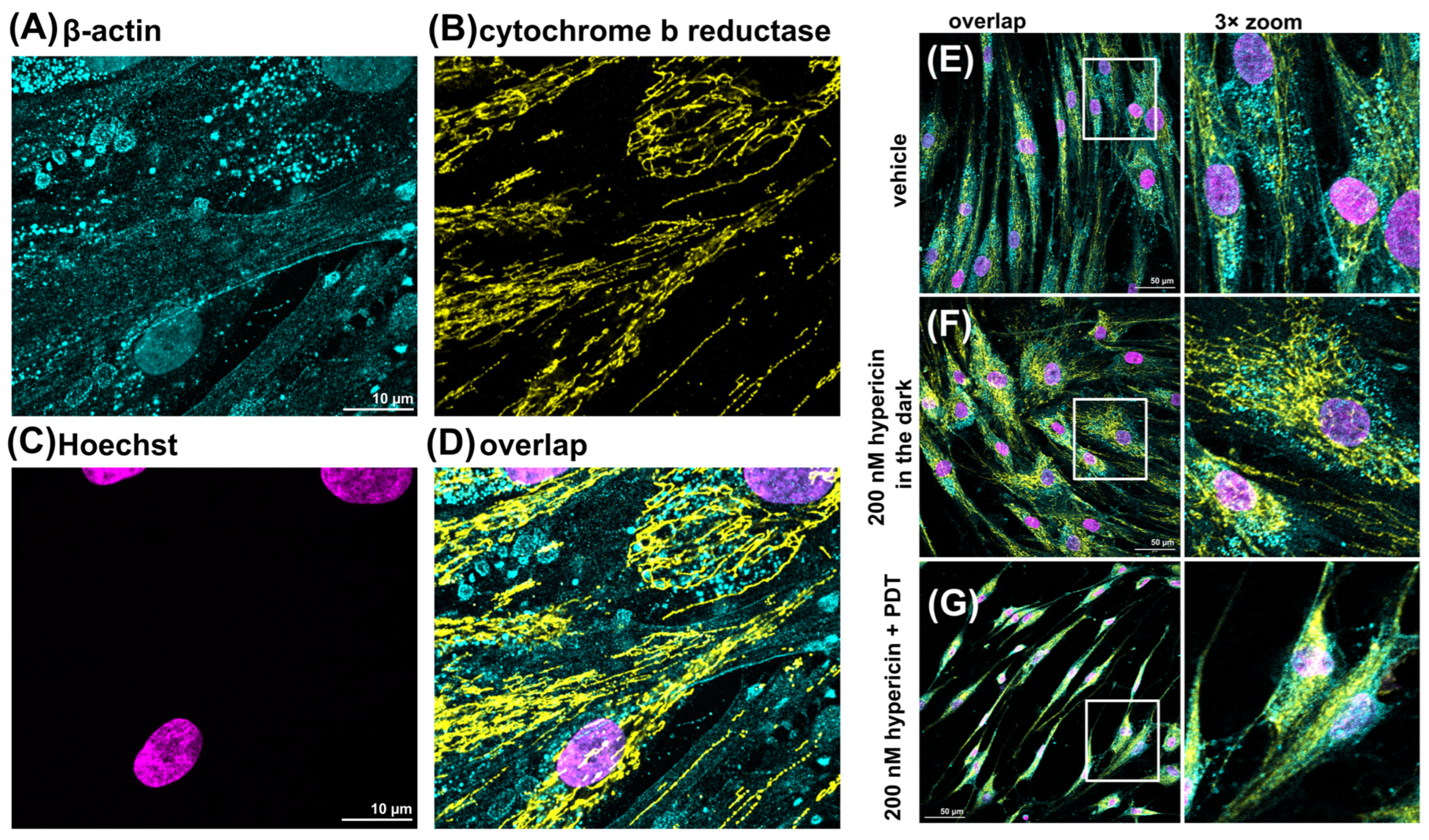
2.4. Confocal Fluorescence Microscopy
2.5. Stimulated Emission Depletion (STED) Microscopy
2.6. Statistical Analysis
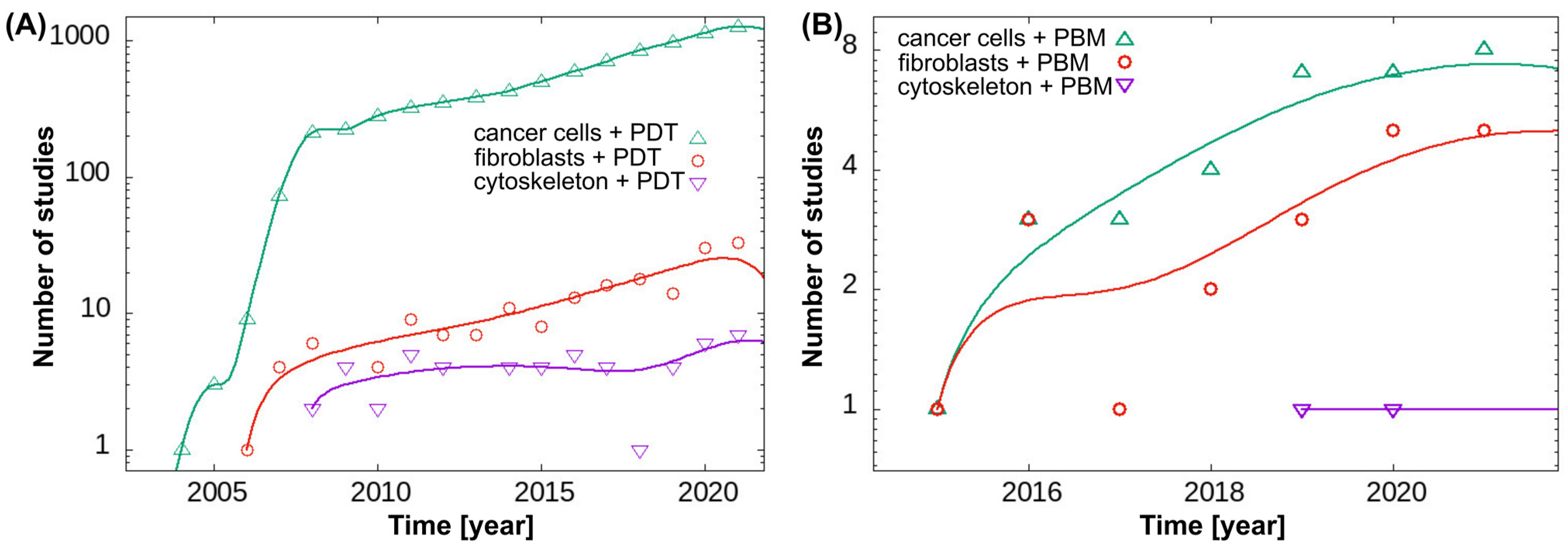
2.7. An Overview of the Study Method with an Emphasis on Relevant Publication Trends
3. Results
3.1. The Metabolic and Structural Alteration of Mitochondria Induced by Light
3.2. The Organization of β-actin in HDF Cells Subjected to PDT and PBM
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hohmann, T.; Dehghani, F. The Cytoskeleton—A Complex Interacting Meshwork. Cells 2019, 8, 362. [Google Scholar] [CrossRef] [PubMed]
- Rotty, J.D.; Bear, J.E. Competition and Collaboration between Different Actin Assembly Pathways Allows for Homeostatic Control of the Actin Cytoskeleton. Bioarchitecture 2014, 5, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Nakamura, F. Actin-Associated Proteins and Small Molecules Targeting the Actin Cytoskeleton. Int. J. Mol. Sci. 2022, 23, 2118. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Dominguez, R. Regulation of Actin Cytoskeleton Dynamics in Cells. Mol. Cells 2010, 29, 311–325. [Google Scholar] [CrossRef] [PubMed]
- Hartman, M.A.; Spudich, J.A. The Myosin Superfamily at a Glance. J. Cell Sci. 2012, 125, 1627–1632. [Google Scholar] [CrossRef]
- Weber, L.P.; Seto, M.; Sasaki, Y.; Swärd, K.; Walsh, M.P. The Involvement of Protein Kinase C in Myosin Phosphorylation and Force Development in Rat Tail Arterial Smooth Muscle. Biochem. J. 2000, 352, 573–582. [Google Scholar] [CrossRef]
- Dugina, V.B.; Shagieva, G.S.; Shakhov, A.S.; Alieva, I.B. The Cytoplasmic Actins in the Regulation of Endothelial Cell Function. Int. J. Mol. Sci. 2021, 22, 7836. [Google Scholar] [CrossRef]
- Illescas, M.; Peñas, A.; Arenas, J.; Martín, M.A.; Ugalde, C. Regulation of Mitochondrial Function by the Actin Cytoskeleton. Front. Cell Dev. Biol. 2021, 9, 795838. [Google Scholar] [CrossRef]
- Boldogh, I.R.; Pon, L.A. Interactions of Mitochondria with the Actin Cytoskeleton. Biochim. Biophys. Acta Mol. Cell Res. 2006, 1763, 450–462. [Google Scholar] [CrossRef]
- Xie, X.; Venit, T.; Drou, N.; Percipalle, P. In Mitochondria β-Actin Regulates MtDNA Transcription and Is Required for Mitochondrial Quality Control. iScience 2018, 3, 226–237. [Google Scholar] [CrossRef]
- Ferguson, S.M.; De Camilli, P. Dynamin, a Membrane-Remodelling GTPase. Nat. Rev. Mol. Cell Biol. 2012, 13, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Cipolat, S.; De Brito, O.M.; Dal Zilio, B.; Scorrano, L. OPA1 Requires Mitofusin 1 to Promote Mitochondrial Fusion. Proc. Natl. Acad. Sci. USA 2004, 101, 15927–15932. [Google Scholar] [CrossRef] [PubMed]
- Ashrafi, G.; Schwarz, T.L. The Pathways of Mitophagy for Quality Control and Clearance of Mitochondria. Cell Death Differ. 2013, 20, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Kast, D.J.; Dominguez, R. The Cytoskeleton–Autophagy Connection. Curr. Biol. 2017, 27, R318–R326. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Mehrvar, S.; Maleki, S.; Schmitt, H.; Summerfelt, P.; Dubis, A.M.; Abroe, B.; Connor, T.B.; Carroll, J.; Huddleston, W.; et al. Photobiomodulation Preserves Mitochondrial Redox State and Is Retinoprotective in a Rodent Model of Retinitis Pigmentosa. Sci. Rep. 2020, 10, 20382. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, F.d.S.; Gonzalez-Lima, F.; Coimbra, N.C. Mitochondrial Photobiomodulation as a Neurotherapeutic Strategy for Epilepsy. Front. Neurol. 2022, 13, 873496. [Google Scholar] [CrossRef]
- Hamblin, M.R. Mechanisms and Mitochondrial Redox Signaling in Photobiomodulation. Photochem. Photobiol. 2018, 94, 199–212. [Google Scholar] [CrossRef]
- Pastore, D.; Greco, M.; Passarella, S. Specific Helium-Neon Laser Sensitivity of the Purified Cytochrome c Oxidase. Int. J. Radiat. Biol. 2000, 76, 863–870. [Google Scholar] [CrossRef]
- Pevna, V.; Horvath, D.; Wagnieres, G.; Huntosova, V. Photobiomodulation and Photodynamic Therapy-Induced Switching of Autophagy and Apoptosis in Human Dermal Fibroblasts. J. Photochem. Photobiol. B Biol. 2022, 234, 112539. [Google Scholar] [CrossRef]
- Pevna, V.; Wagnières, G.; Huntosova, V. Autophagy and Apoptosis Induced in U87 Mg Glioblastoma Cells by Hypericin-Mediated Photodynamic Therapy Can Be Photobiomodulated with 808 Nm Light. Biomedicines 2021, 9, 1703. [Google Scholar] [CrossRef]
- Pevná, V.; Wagnières, G.; Jancura, D.; Huntošová, V. Effect of Photobiomodulation on Protein Kinase Cδ, Cytochrome C, and Mitochondria in U87 MG Cells. Cells 2023, 12, 1441. [Google Scholar] [CrossRef] [PubMed]
- de Magalhães, A.C.; Guimarães-Filho, Z.; Yoshimura, E.M.; Lilge, L. Photobiomodulation Therapy Can Change Actin Filaments of 3T3 Mouse Fibroblast. Lasers Med. Sci. 2020, 35, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Mokoena, D.R.; Houreld, N.N.; Dhilip Kumar, S.S.; Abrahamse, H. Photobiomodulation at 660 Nm Stimulates Fibroblast Differentiation. Lasers Surg. Med. 2020, 52, 671–681. [Google Scholar] [CrossRef]
- Castano, A.P.; Demidova, T.N.; Hamblin, M.R. Mechanisms in Photodynamic Therapy: Part One–Photosensitizers, Photochemistry and Cellular Localization. Photodiagnosis Photodyn. Ther. 2004, 1, 279–293. [Google Scholar] [CrossRef] [PubMed]
- Kwiatkowski, S.; Knap, B.; Przystupski, D.; Saczko, J.; Kędzierska, E.; Knap-Czop, K.; Kotlińska, J.; Michel, O.; Kotowski, K.; Kulbacka, J. Photodynamic Therapy–Mechanisms, Photosensitizers and Combinations. Biomed. Pharmacother. 2018, 106, 1098–1107. [Google Scholar] [CrossRef]
- Agostinis, P.; Berg, K.; Cengel, K.A.; Foster, T.H.; Girotti, A.W.; Gollnick, S.O.; Hahn, S.M.; Hamblin, M.R.; Juzeniene, A.; Kessel, D.; et al. Photodynamic Therapy of Cancer: An Update. CA Cancer J. Clin. 2011, 61, 250–281. [Google Scholar] [CrossRef]
- Gunaydin, G.; Gedik, M.E.; Ayan, S. Photodynamic Therapy for the Treatment and Diagnosis of Cancer–A Review of the Current Clinical Status. Front. Chem. 2021, 9, 686303. [Google Scholar] [CrossRef]
- Yoo, S.W.; Oh, G.; Ahn, J.C.; Chung, E. Non-Oncologic Applications of Nanomedicine-Based Photo-Therapy. Biomedicines 2021, 9, 113. [Google Scholar] [CrossRef]
- Dubey, T.; Chinnathambi, S. Photodynamic Sensitizers Modulate Cytoskeleton Structural Dynamics in Neuronal Cells. Cytoskeleton 2021, 78, 232–248. [Google Scholar] [CrossRef]
- Malohlava, J.; Tomankova, K.; Malina, L.; Jiravova, J.; Hanakova, A.; Pizova, K.; Zapletalova, J.; Kolarova, H. Effect of Porphyrin Sensitizer MgTPPS4 on Cytoskeletal System of HeLa Cell Line—Microscopic Study. Cell Biochem. Biophys. 2016, 74, 419–425. [Google Scholar] [CrossRef]
- Jendželovská, Z.; Jendželovský, R.; Kuchárová, B.; Fedoročko, P. Hypericin in the Light and in the Dark: Two Sides of the Same Coin. Front. Plant Sci. 2016, 7, 560. [Google Scholar] [CrossRef] [PubMed]
- Lenkavska, L.; Tomkova, S.; Horvath, D.; Huntosova, V. Searching for Combination Therapy by Clustering Methods: Stimulation of PKC in Golgi Apparatus Combined with Hypericin Induced PDT. Photodiagnosis Photodyn. Ther. 2020, 31, 101813. [Google Scholar] [CrossRef] [PubMed]
- Misuth, M.; Joniova, J.; Horvath, D.; Dzurova, L.; Nichtova, Z.; Novotova, M.; Miskovsky, P.; Stroffekova, K.; Huntosova, V. The Flashlights on a Distinct Role of Protein Kinase C δ: Phosphorylation of Regulatory and Catalytic Domain upon Oxidative Stress in Glioma Cells. Cell. Signal. 2017, 34, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Dzurová, L.; Petrovajova, D.; Nadova, Z.; Huntosova, V.; Miskovsky, P.; Stroffekova, K. The Role of Anti-Apoptotic Protein Kinase Cα in Response to Hypericin Photodynamic Therapy in U-87 MG Cells. Photodiagnosis Photodyn. Ther. 2014, 11, 213–226. [Google Scholar] [CrossRef] [PubMed]
- Kılıç Süloğlu, A.; Selmanoğlu, G.; Akay, M.T. Alterations in Dysadherin Expression and F-Actin Reorganization: A Possible Mechanism of Hypericin-Mediated Photodynamic Therapy in Colon Adenocarcinoma Cells. Cytotechnology 2015, 67, 311–330. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, L.A.; Pinheiro, S.L. Clinical Evaluation of Intravascular Blood Irradiation with Laser, Photobiomodulation, and Photodynamic Therapy in Cancer Patients with Mucositis. Photobiomodulation Photomed. Laser Surg. 2021, 39, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, S.L.; Bonadiman, A.C.; Dos Anjos Borges Lemos, A.L.; Annicchino, B.M.; Segatti, B.; Pucca, D.S.; Dutra, P.T.; De Carvalho E Silva, R.M.; Leal, F. Photobiomodulation Therapy in Cancer Patients with Mucositis: A Clinical Evaluation. Photobiomodulation Photomed. Laser Surg. 2019, 37, 142–150. [Google Scholar] [CrossRef]
- Pedroni, A.C.F.; Miniello, T.G.; Hirota, C.; Carvalho, M.H.; Lascala, C.A.; Marques, M.M. Successful Application of Antimicrobial Photodynamic and Photobiomodulation Therapies for Controlling Osteoradionecrosis and Xerostomia after Laryngeal Carcinoma Treatment: A Case Report of Full Oral Rehabilitation. Photodiagnosis Photodyn. Ther. 2020, 31, 101835. [Google Scholar] [CrossRef]
- Rezende, S.B.; Campos, L.; Palma, L.F.; Tateno, R.Y.; Simões, A.; Macedo, M.C.; da Silva, R.L. Photobiomodulation and Antimicrobial Photodynamic Therapy for Oral Cytomegalovirus Reactivation Following Acute Graft-versus-Host Disease. Photodiagnosis Photodyn. Ther. 2020, 32, 101849. [Google Scholar] [CrossRef]
- Negri, L.B.; Martins, T.J.; da Silva, R.S.; Hamblin, M.R. Photobiomodulation Combined with Photodynamic Therapy Using Ruthenium Phthalocyanine Complexes in A375 Melanoma Cells: Effects of Nitric Oxide Generation and ATP Production. J. Photochem. Photobiol. B Biol. 2019, 198, 111564. [Google Scholar] [CrossRef]
- de Faria, C.M.G.; Costa, C.S.; Bagnato, V.S. Photobiomodulation Effects on Photodynamic Therapy in HNSCC Cell Lines. J. Photochem. Photobiol. B Biol. 2021, 217, 112170. [Google Scholar] [CrossRef] [PubMed]
- Joniová, J.; Kazemiraad, C.; Gerelli, E.; Wagnières, G. Stimulation and Homogenization of the Protoporphyrin IX Endogenous Production by Photobiomodulation to Increase the Potency of Photodynamic Therapy. J. Photochem. Photobiol. B Biol. 2021, 225, 112347. [Google Scholar] [CrossRef] [PubMed]
- Aniogo, E.C.; George, B.P.; Abrahamse, H. Photobiomodulation Improves Anti-Tumor Efficacy of Photodynamic Therapy against Resistant MCF-7 Cancer Cells. Biomedicines 2023, 11, 1547. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Huntošová, V.; Datta, S.; Lenkavská, L.; MáčAjová, M.; Bilčík, B.; Kundeková, B.; Čavarga, I.; Kronek, J.; Jutková, A.; Miškovský, P.; et al. Alkyl Chain Length in Poly(2-Oxazoline)-Based Amphiphilic Gradient Copolymers Regulates the Delivery of Hydrophobic Molecules: A Case of the Biodistribution and the Photodynamic Activity of the Photosensitizer Hypericin. Biomacromolecules 2021, 22, 4199–4216. [Google Scholar] [CrossRef] [PubMed]
- Lenkavska, L.; Blascakova, L.; Jurasekova, Z.; Macajova, M.; Bilcik, B.; Cavarga, I.; Miskovsky, P.; Huntosova, V. Benefits of Hypericin Transport and Delivery by Low- and High-Density Lipoproteins to Cancer Cells: From in Vitro to Ex Ovo. Photodiagnosis Photodyn. Ther. 2019, 25, 214–224. [Google Scholar] [CrossRef]
- Ritz, R.; Wein, H.T.; Dietz, K.; Schenk, M.; Roser, F.; Tatagiba, M.; Strauss, W.S.L. Photodynamic Therapy of Malignant Glioma with Hypericin: Comprehensive in Vitro Study in Human Glioblastoma Cell Lines. Int. J. Oncol. 2007, 30, 659–667. [Google Scholar] [CrossRef]
- Ritz, R.; Daniels, R.; Noell, S.; Feigl, G.C.; Schmidt, V.; Bornemann, A.; Ramina, K.; Mayer, D.; Dietz, K.; Strauss, W.S.L.; et al. Hypericin for Visualization of High Grade Gliomas: First Clinical Experience. Eur. J. Surg. Oncol. 2012, 38, 352–360. [Google Scholar] [CrossRef]
- Pevná, V.; Máčajová, M.; Hovan, A.; Bánó, G.; Meta, M.; Bilčík, B.; Palková, J.; Huntošová, V. Spheroidal Model of SKBR3 and U87MG Cancer Cells for Live Imaging of Caspase-3 during Apoptosis Induced by Singlet Oxygen in Photodynamic Therapy. Biomedicines 2022, 10, 2141. [Google Scholar] [CrossRef] [PubMed]
- Kamuhabwa, A.; Agostinis, P.; Ahmed, B.; Landuyt, W.; Van Cleynenbreugel, B.; Van Poppel, H.; de Witte, P. Hypericin as a Potential Phototherapeutic Agent in Superficial Transitional Cell Carcinoma of the Bladder. Photochem. Photobiol. Sci. 2004, 3, 772–780. [Google Scholar] [CrossRef]
- Theodossiou, T.A.; Hothersall, J.S.; De Witte, P.A.; Pantos, A.; Agostinis, P. The Multifaceted Photocytotoxic Profile of Hypericin. Mol. Pharm. 2009, 6, 1775–1789. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, X.K.; Zhu, Z.J.; Liang, Z.W.; Li, P.H.; Ma, Y.G.; Ding, T.; Li, K.; Zuo, X.S.; Ju, C.; et al. Photobiomodulation Provides Neuroprotection through Regulating Mitochondrial Fission Imbalance in the Subacute Phase of Spinal Cord Injury. Neural Regen. Res. 2023, 18, 2005–2010. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.C.; Hang, N.L.T.; Colley, M.M.S.; Chang, J.; Hsiao, Y.C.; Lu, L.S.; Li, B.S.; Chang, C.J.; Yang, T. Sen Single Cell Effects of Photobiomodulation on Mitochondrial Membrane Potential and Reactive Oxygen Species Production in Human Adipose Mesenchymal Stem Cells. Cells 2022, 11, 972. [Google Scholar] [CrossRef]
- Suzuki, T.; Tanaka, M.; Sasaki, M.; Ichikawa, H.; Nishie, H.; Kataoka, H. Vascular Shutdown by Photodynamic Therapy Using Talaporfin Sodium. Cancers 2020, 12, 2369. [Google Scholar] [CrossRef]
- Pires Marques, E.C.; Piccolo Lopes, F.; Nascimento, I.C.; Morelli, J.; Pereira, M.V.; Machado Meiken, V.M.; Pinheiro, S.L. Photobiomodulation and Photodynamic Therapy for the Treatment of Oral Mucositis in Patients with Cancer. Photodiagnosis Photodyn. Ther. 2020, 29, 101621. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Photosensitizer | PBM Wavelength | PDT Wavelength | Model | Disease | Reference |
|---|---|---|---|---|---|
| In Vitro/In Vivo | |||||
| curcumin | 660 nm | * 468 nm | human oral cavity | oral mucositis | [36] |
| human intravascular blood | |||||
| curcumin | 660 nm | * 468 nm | human oral cavity | oral mucositis | [55] |
| hypericin | * 808 nm | 590 nm | U87 MG | glioblastoma multiforme - | [19] |
| HDF | |||||
| zinc tetrasulfonic acid phthalocyanine (ZnPcS4) | 681 nm | 680 nm | MCF-7 | breast cancer | [43] |
| methylene blue | 660 nm (0.3 J, 3 s per point) | * 660 nm (5 J, 50 s per point) | human oral cavity | oral cytomegalovirus infection | [39] |
| ruthenium complexes [Ru(Pc)] or trans-[Ru(NO)(NO2)(Pc)] | 850 nm | * 660 nm | A375 | melanoma | [40] |
| Photogem® | * 780 nm | 630 nm | SCC-25 and SCC-4 | head and neck squamous cell carcinomas | [41] |
| curcumin | 660 nm | * 468 nm | human oral cavity | oral mucositis | [37] |
| methylene blue | 660 nm (1 J, 10 s per point | * 660 nm (2 J, 20 s per point) | human oral cavity | osteoradionecrosis | [38] |
| 808 nm (2 J, 20 s per point) | top of the major salivary glands | xerostomia |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olejárová, S.; Horváth, D.; Huntošová, V. The Remodulation of Actin Bundles during the Stimulation of Mitochondria in Adult Human Fibroblasts in Response to Light. Pharmaceutics 2024, 16, 20. https://doi.org/10.3390/pharmaceutics16010020
Olejárová S, Horváth D, Huntošová V. The Remodulation of Actin Bundles during the Stimulation of Mitochondria in Adult Human Fibroblasts in Response to Light. Pharmaceutics. 2024; 16(1):20. https://doi.org/10.3390/pharmaceutics16010020
Chicago/Turabian StyleOlejárová, Soňa, Denis Horváth, and Veronika Huntošová. 2024. "The Remodulation of Actin Bundles during the Stimulation of Mitochondria in Adult Human Fibroblasts in Response to Light" Pharmaceutics 16, no. 1: 20. https://doi.org/10.3390/pharmaceutics16010020
APA StyleOlejárová, S., Horváth, D., & Huntošová, V. (2024). The Remodulation of Actin Bundles during the Stimulation of Mitochondria in Adult Human Fibroblasts in Response to Light. Pharmaceutics, 16(1), 20. https://doi.org/10.3390/pharmaceutics16010020