
Photodynamic Therapy for X-ray-Induced Radiation-Resistant Cancer Cells




Abstract
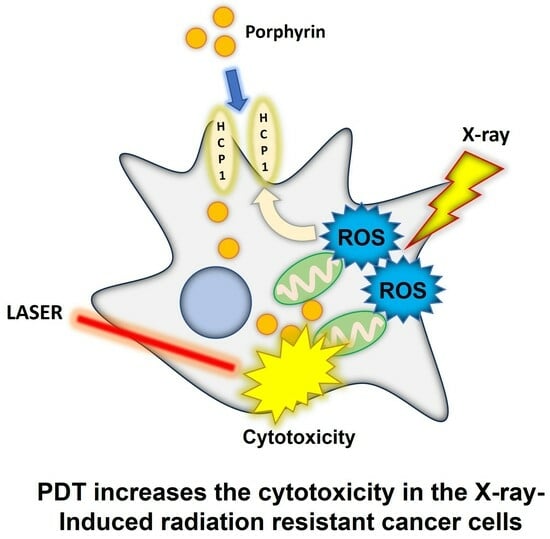
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. Establishment of X-ray-Resistant Cells
2.4. Intracellular ROS Production
2.5. Detection of Mitochondrial ROS
2.6. Measurement of HCP Expression
2.7. Intracellular Porphyrin Accumulation Assay
2.8. Cytotoxic Assay
2.9. Statistical Analysis
3. Results
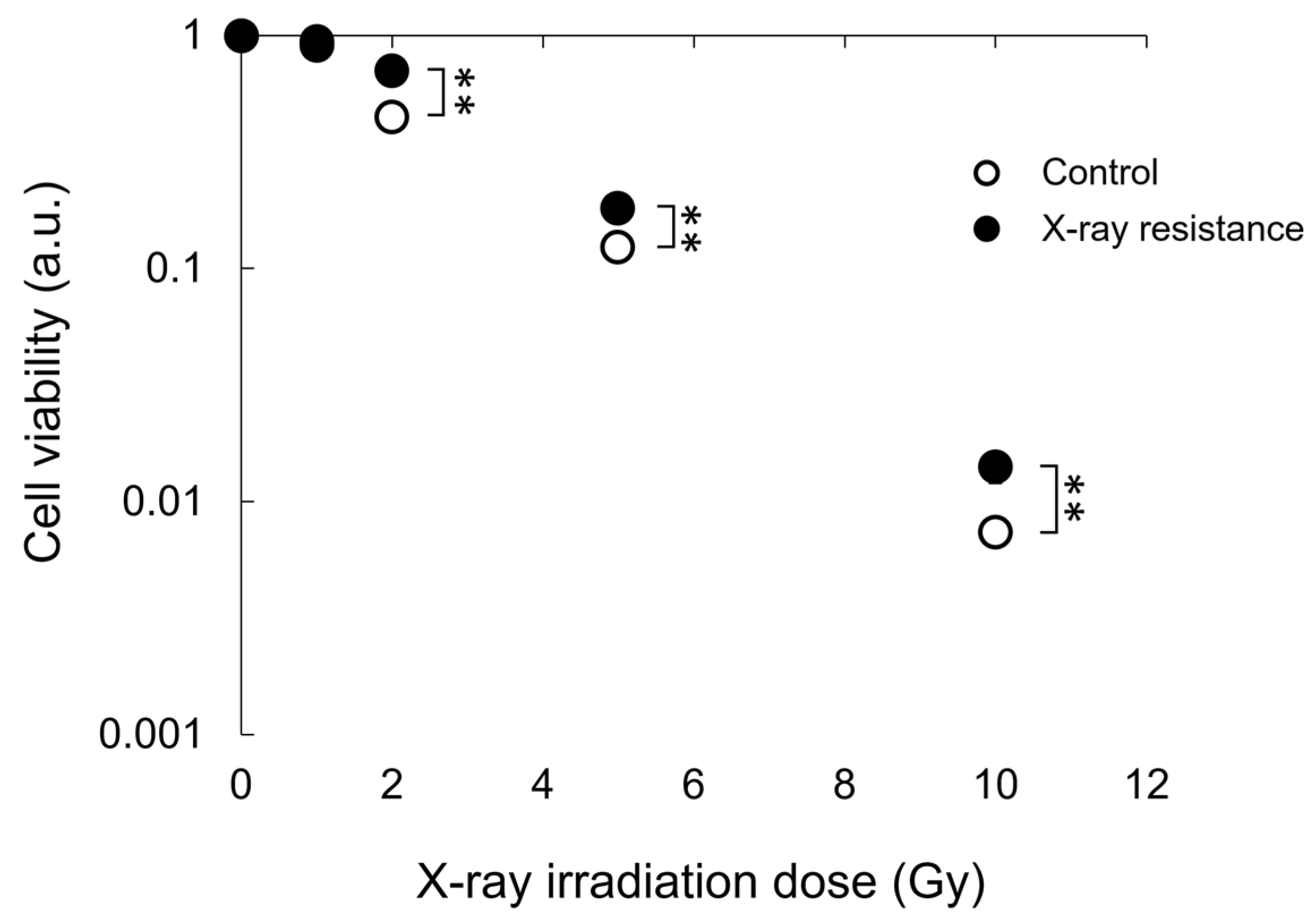
3.1. Confirmation of X-ray Resistance by Colony Formation Assay
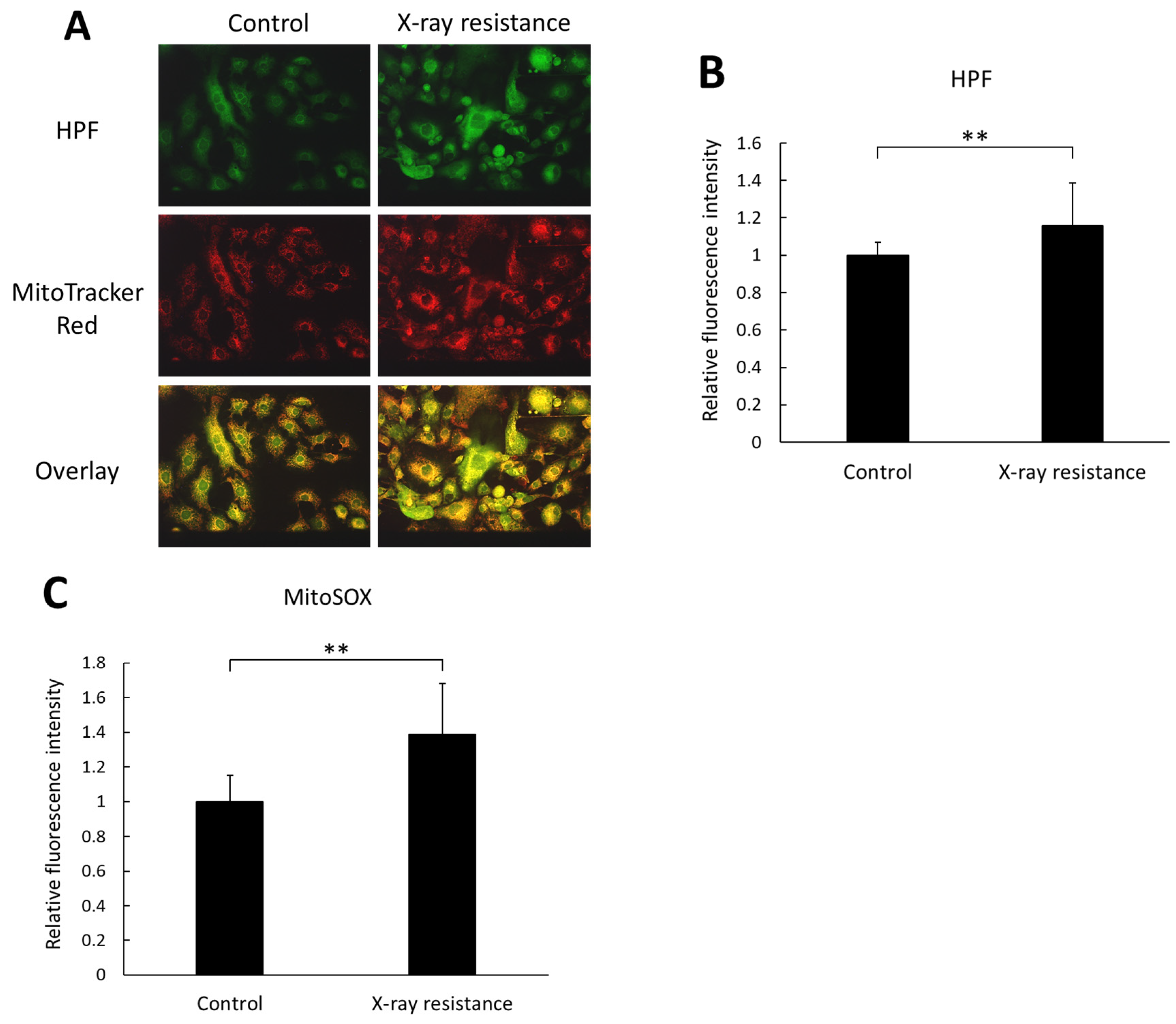
3.2. Increase in Intracellular and Mitochondria-Specific ROS Production in X-ray-Resistant Cells
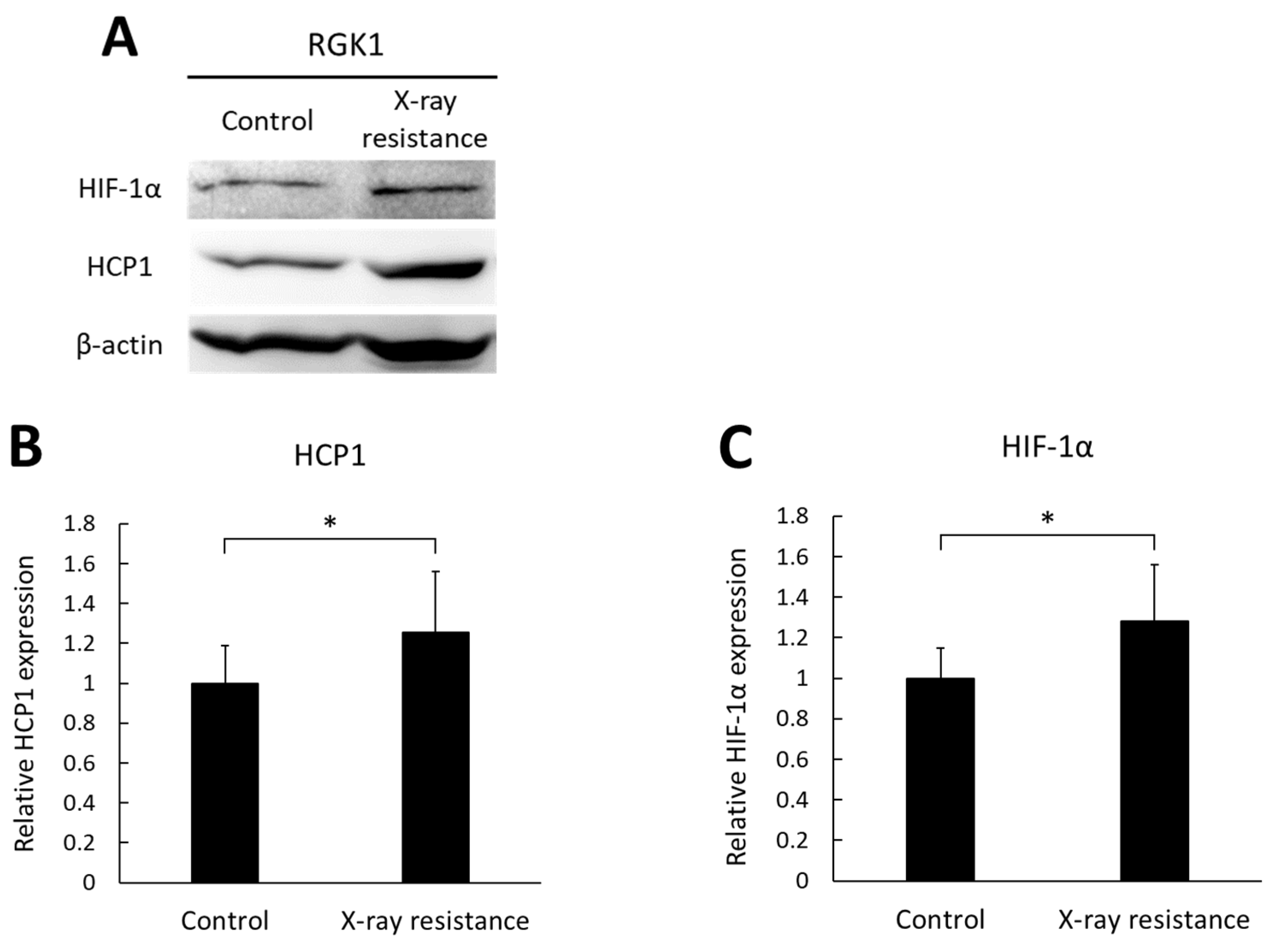
3.3. Increase in HCP1 and HIF-1α Expressions in X-ray-Resistant Cells
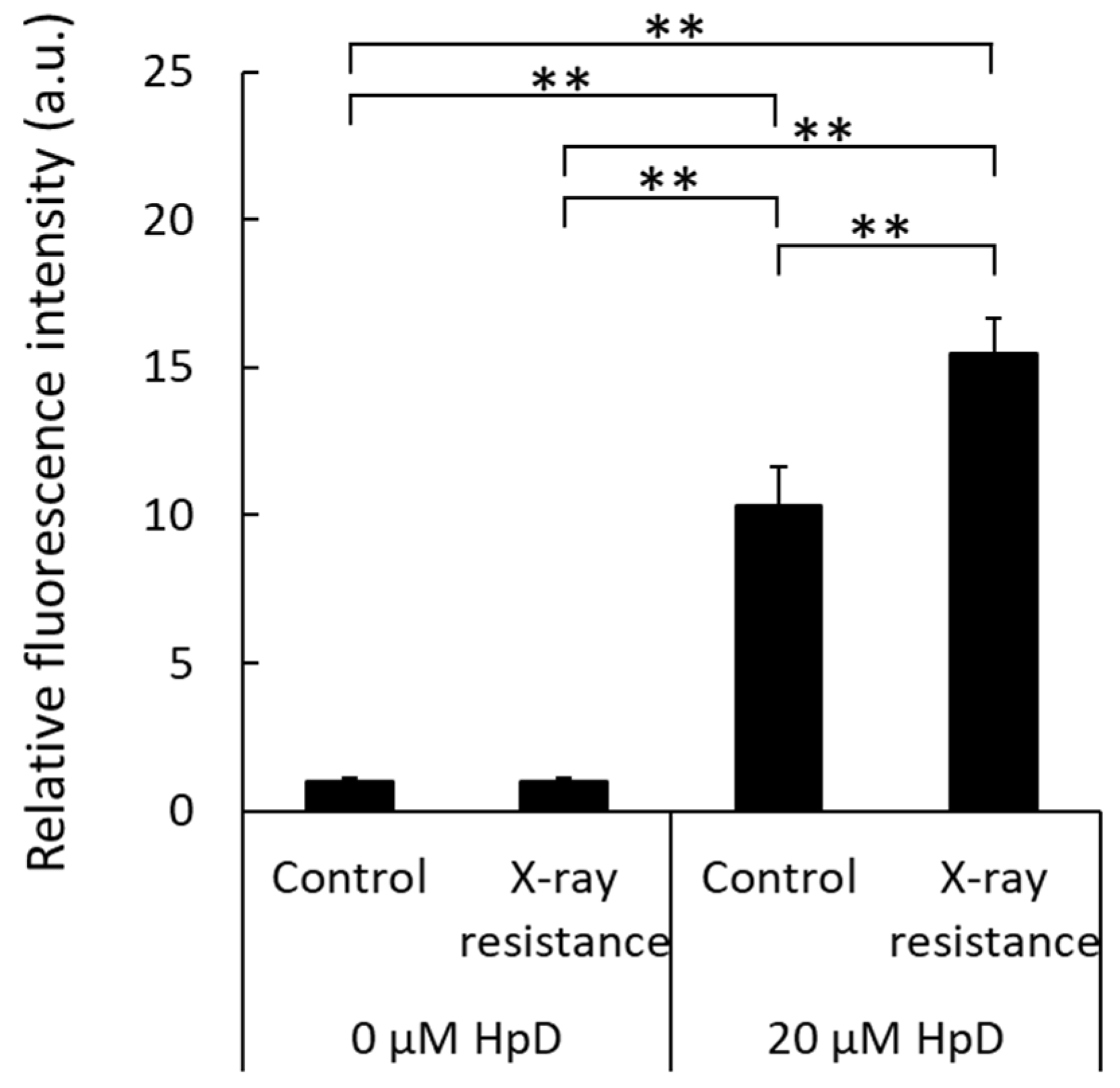
3.4. Increase in Porphyrin Accumulation in X-ray-Resistant Cells
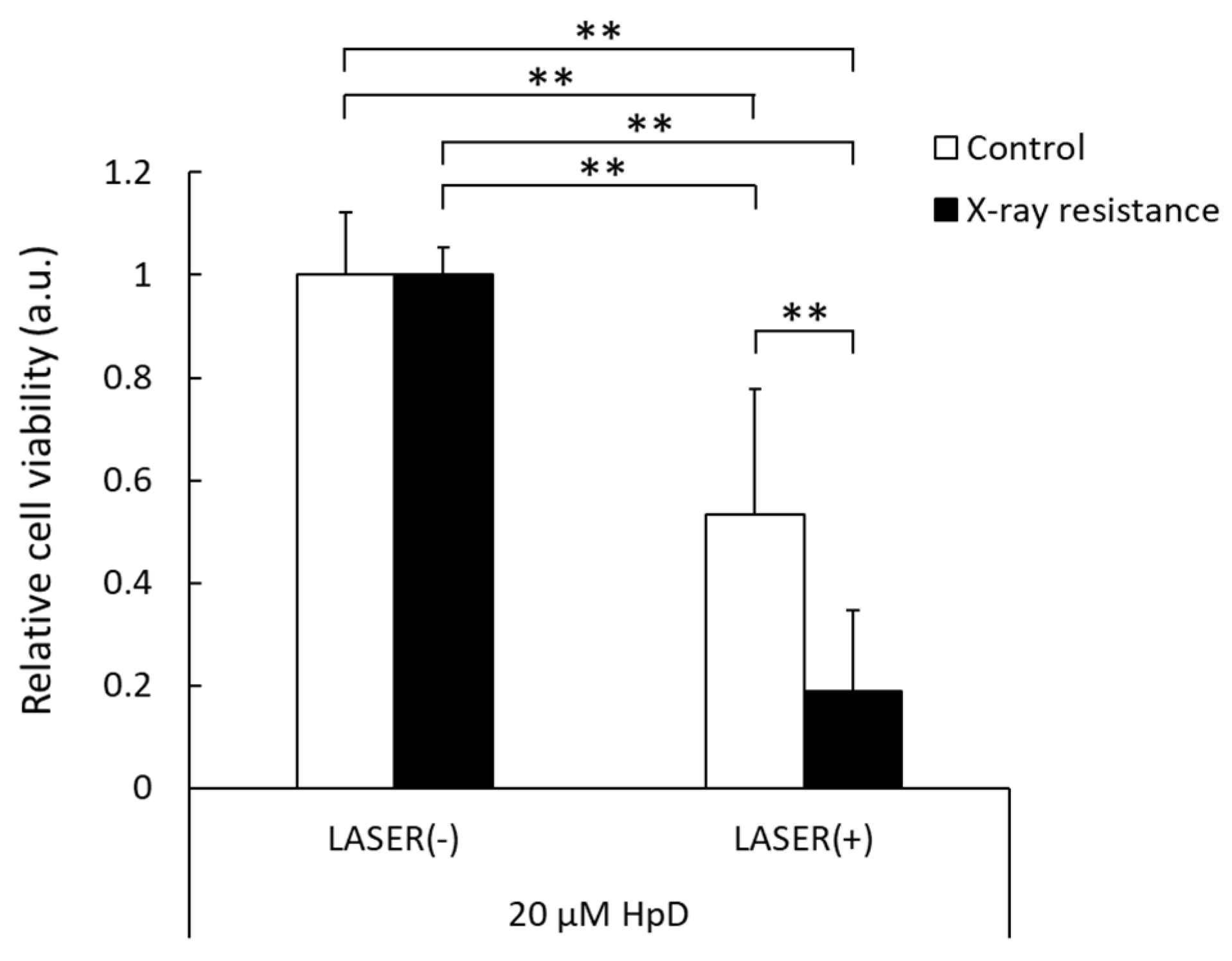
3.5. Decrease in Cell Viability by PDT in X-ray-Resistant Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lalo, R.; Zekja, I.; Kamberi, F. Association of Cardiovascular Disease Risk and Health-Related Behaviors in Stroke Patients. Int. J. Environ. Res. Public Health 2023, 20, 3693. [Google Scholar] [CrossRef] [PubMed]
- Toyoda, K.; Yasaka, M.; Iwade, K.; Nagata, K.; Koretsune, Y.; Sakamoto, T.; Uchiyama, S.; Gotoh, J.; Nagao, T.; Yamamoto, M.; et al. Dual antithrombotic therapy increases severe bleeding events in patients with stroke and cardiovascular disease: A prospective, multicenter, observational study. Stroke 2008, 39, 1740–1745. [Google Scholar] [CrossRef]
- Alam, M.; Goldberg, L.H. Serious Adverse Vascular Events Associated With Perioperative Interruption of Antiplatelet and Anticoagulant Therapy. Dermatolog. Surg. 2002, 28, 992–998. [Google Scholar] [CrossRef]
- Louison, S.; Gabrielle, P.H.; Soudry, A.; Meillon, C.; Blanc, J.; Béal, G.; Arsène, S.; Le Mer, Y.; Berrod, J.P.; Kodjikian, L.; et al. Perioperative risk of bleeding with antithrombotic agents in macular surgery: A national, prospective, multicentre study. Acta Ophthalmol. 2020, 98, e991–e997. [Google Scholar] [CrossRef]
- Vargason, A.M.; Anselmo, A.C.; Mitragotri, S. The evolution of commercial drug delivery technologies. Nat. Biomed. Eng. 2021, 5, 951–967. [Google Scholar] [CrossRef] [PubMed]
- Senapati, S.; Mahanta, A.K.; Kumar, S.; Maiti, P. Controlled drug delivery vehicles for cancer treatment and their performance. Signal Transduct. Target. Ther. 2018, 3, 7. [Google Scholar] [CrossRef]
- Ryan, J.L. Ionizing Radiation: The Good, the Bad, and the Ugly. J. Investig. Dermatol. 2012, 132, 985–993. [Google Scholar] [CrossRef]
- Naidu, M.U.R.; Ramana, G.V.; Rani, P.U.; Mohan, I.K.; Suman, A.; Roy, P. Chemotherapy-induced and/or radiation therapy-induced oral mucositis—Complicating the treatment of cancer. Neoplasia 2004, 6, 423–431. [Google Scholar] [CrossRef]
- Langendijk, J.A.; Lambin, P.; De Ruysscher, D.; Widder, J.; Bos, M.; Verheij, M. Selection of patients for radiotherapy with protons aiming at reduction of side effects: The model-based approach. Radiother. Oncol. 2013, 107, 267–273. [Google Scholar] [CrossRef]
- Schlaff, C.D.; Krauze, A.; Belard, A.; O’Connell, J.J.; Camphausen, K.A. Bringing the heavy: Carbon ion therapy in the radiobiological and clinical context. Radiother. Oncol. 2014, 9, 88. [Google Scholar] [CrossRef]
- Arruebo, M.; Vilaboa, N.; Sáez-Gutierrez, B.; Lambea, J.; Tres, A.; Valladares, M.; González-Fernández, Á. Assessment of the evolution of cancer treatment therapies. Cancers 2011, 3, 3279–3330. [Google Scholar] [CrossRef]
- Ohno, T. Particle radiotherapy with carbon ion beams. EPMA J. 2013, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Kamada, T.; Tsujii, H.; Blakely, E.A.; Debus, J.; De Neve, W.; Durante, M.; Jäkel, O.; Mayer, R.; Orecchia, R.; Pötter, R.; et al. Carbon ion radiotherapy in Japan: An assessment of 20 years of clinical experience. Lancet Oncol. 2015, 16, e93–e100. [Google Scholar] [CrossRef] [PubMed]
- Galeaz, C.; Totis, C.; Bisio, A. Radiation Resistance: A Matter of Transcription Factors. Front. Oncol. 2021, 11, 662840. [Google Scholar] [CrossRef] [PubMed]
- Alamilla-Presuel, J.C.; Burgos-Molina, A.M.; González-Vidal, A.; Sendra-Portero, F.; Ruiz-Gómez, M.J. Factors and molecular mechanisms of radiation resistance in cancer cells. Int. J. Radiat. Biol. 2022, 98, 1301–1315. [Google Scholar] [CrossRef]
- Nishida, K.; Tojo, T.; Kondo, T.; Yuasa, M. Evaluation of the correlation between porphyrin accumulation in cancer cells and functional positions for application as a drug carrier. Sci. Rep. 2021, 11, 2046. [Google Scholar] [CrossRef]
- Castano, A.P.; Mroz, P.; Hamblin, M.R. Photodynamic therapy and anti-tumour immunity. Nat. Rev. Cancer 2006, 6, 535–545. [Google Scholar] [CrossRef]
- Kaneko, S.; Kaneko, S. Fluorescence-Guided Resection of Malignant Glioma with 5-ALA. Int. J. Biomed. Imaging 2016, 2016, 6135293. [Google Scholar] [CrossRef]
- Hiyama, K.; Matsui, H.; Tamura, M.; Shimokawa, O.; Hiyama, M.; Kaneko, T.; Nagano, Y.; Hyodo, I.; Tanaka, J.; Miwa, Y.; et al. Cancer cells uptake porphyrins via heme carrier protein 1. J. Porphyr. Phthalocyanines 2013, 17, 36–43. [Google Scholar] [CrossRef]
- Ito, H.; Matsui, H.; Tamura, M.; Majima, H.J.; Indo, H.P.; Hyodo, I. Mitochondrial reactive oxygen species accelerate the expression of heme carrier protein 1 and enhance photodynamic cancer therapy effect. J. Clin. Biochem. Nutr. 2014, 55, 67–71. [Google Scholar] [CrossRef]
- Ogura, A.; Oowada, S.; Kon, Y.; Hirayama, A.; Yasui, H.; Meike, S.; Kobayashi, S.; Kuwabara, M.; Inanami, O. Redox regulation in radiation-induced cytochrome c release from mitochondria of human lung carcinoma A549 cells. Cancer Lett. 2009, 277, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Shimokawa, O.; Matsui, H.; Nagano, Y.; Kaneko, T.; Shibahara, T.; Nakahara, A.; Hyodo, I.; Yanaka, A.; Majima, H.J.; Nakamura, Y.; et al. Neoplastic transformation and induction of H+, K+-adenosine triphosphatase by N-methyl-N′-nitro-N-nitrosoguanidine in the gastric epithelial RGM-1 cell line. Vitr. Cell. Dev. Biol.-Anim. 2008, 44, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Setsukinai, K.; Urano, Y.; Kakinuma, K.; Majima, H.J.; Nagano, T. Development of novel fluorescence probes that can reliably detect reactive oxygen species and distinguish specific species. J. Biol. Chem. 2003, 278, 3170–3175. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-M.; Shah, A.M. Endothelial cell superoxide generation: Regulation and relevance for cardiovascular pathophysiology. Am. J. Physiol. Integr. Comp. Physiol. 2004, 287, R1014–R1030. [Google Scholar] [CrossRef]
- Indo, H.P.; Yen, H.C.; Nakanishi, I.; Matsumoto, K.I.; Tamura, M.; Nagano, Y.; Matsui, H.; Gusev, O.; Cornette, R.; Okuda, T.; et al. A mitochondrial superoxide theory for oxidative stress diseases and aging. J. Clin. Biochem. Nutr. 2015, 56, 1–7. [Google Scholar] [CrossRef]
- Kurokawa, H.; Ito, H.; Terasaki, M.; Matano, D.; Taninaka, A.; Shigekawa, H.; Matsui, H. Nitric oxide regulates the expression of heme carrier protein-1 via hypoxia inducible factor-1α stabilization. PLoS ONE 2019, 14, e0222074. [Google Scholar] [CrossRef]
- Koike, M.; Yutoku, Y.; Koike, A. Inhibition of crandell-rees feline kidney cell proliferation by x-ray-induced senescence. J. Vet. Med. Sci. 2021, 83, 798–804. [Google Scholar] [CrossRef]
- Majima, H.J.; Oberley, T.D.; Furukawa, K.; Mattson, M.P.; Yen, H.C.; Szweda, L.I.; St. Clair, D.K. Prevention of mitochondrial injury by manganese superoxide dismutase reveals a primary mechanism for alkaline-induced cell death. J. Biol. Chem. 1998, 273, 8217–8224. [Google Scholar] [CrossRef]
- Shayeghi, M.; Latunde-Dada, G.O.; Oakhill, J.S.; Laftah, A.H.; Takeuchi, K.; Halliday, N.; Khan, Y.; Warley, A.; McCann, F.E.; Hider, R.C.; et al. Identification of an intestinal heme transporter. Cell 2005, 122, 789–801. [Google Scholar] [CrossRef]
- Klimova, T.; Chandel, N.S. Mitochondrial complex III regulates hypoxic activation of HIF. Cell Death Differ. 2008, 15, 660–666. [Google Scholar] [CrossRef]
- Harada, H.; Inoue, M.; Itasaka, S.; Hirota, K.; Morinibu, A.; Shinomiya, K.; Zeng, L.; Ou, G.; Zhu, Y.; Yoshimura, M.; et al. Cancer cells that survive radiation therapy acquire HIF-1 activity and translocate towards tumour blood vessels. Nat. Commun. 2012, 3, 783. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, H.; Ito, H.; Terasaki, M.; Matsui, H. Hyperthermia enhances photodynamic therapy by regulation of HCP1 and ABCG2 expressions via high level ROS generation. Sci. Rep. 2019, 9, 1638. [Google Scholar] [CrossRef] [PubMed]
- Chi, C.; Ozawa, T.; Anzai, K. In vivo nitric oxide production and iNOs expression in X-ray irradiated mouse skin. Biol. Pharm. Bull. 2006, 29, 348–353. [Google Scholar] [CrossRef]
- Dewhirst, M.W.; Cao, Y.; Moeller, B. Cycling hypoxia and free radicals regulate angiogenesis and radiotherapy response. Nat. Rev. Cancer 2008, 8, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, F.; Ohgari, Y.; Yamaki, N.; Kitajima, S.; Shimokawa, O.; Matsui, H.; Taketani, S. The role of nitric oxide in δ-aminolevulinic acid (ALA)-induced photosensitivity of cancerous cells. Biochem. Biophys. Res. Commun. 2007, 353, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Ito, H. Peroxynitrite Production Induced by LPS and X-ray Treatment Enhances Cellular Incorporation of Porphyrin in Mouse RAW264 Macrophages. Appl. Sci. 2021, 11, 3503. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ito, H.; Shoji, Y.; Ueno, M.; Matsumoto, K.-i.; Nakanishi, I. Photodynamic Therapy for X-ray-Induced Radiation-Resistant Cancer Cells. Pharmaceutics 2023, 15, 2536. https://doi.org/10.3390/pharmaceutics15112536
Ito H, Shoji Y, Ueno M, Matsumoto K-i, Nakanishi I. Photodynamic Therapy for X-ray-Induced Radiation-Resistant Cancer Cells. Pharmaceutics. 2023; 15(11):2536. https://doi.org/10.3390/pharmaceutics15112536
Chicago/Turabian StyleIto, Hiromu, Yoshimi Shoji, Megumi Ueno, Ken-ichiro Matsumoto, and Ikuo Nakanishi. 2023. "Photodynamic Therapy for X-ray-Induced Radiation-Resistant Cancer Cells" Pharmaceutics 15, no. 11: 2536. https://doi.org/10.3390/pharmaceutics15112536
APA StyleIto, H., Shoji, Y., Ueno, M., Matsumoto, K.-i., & Nakanishi, I. (2023). Photodynamic Therapy for X-ray-Induced Radiation-Resistant Cancer Cells. Pharmaceutics, 15(11), 2536. https://doi.org/10.3390/pharmaceutics15112536