



Extracellular Matrices as Bioactive Materials for In Situ Tissue Regeneration
 ,
,
Abstract
:1. Introduction
1.1. Collagens
1.2. Fibronectin
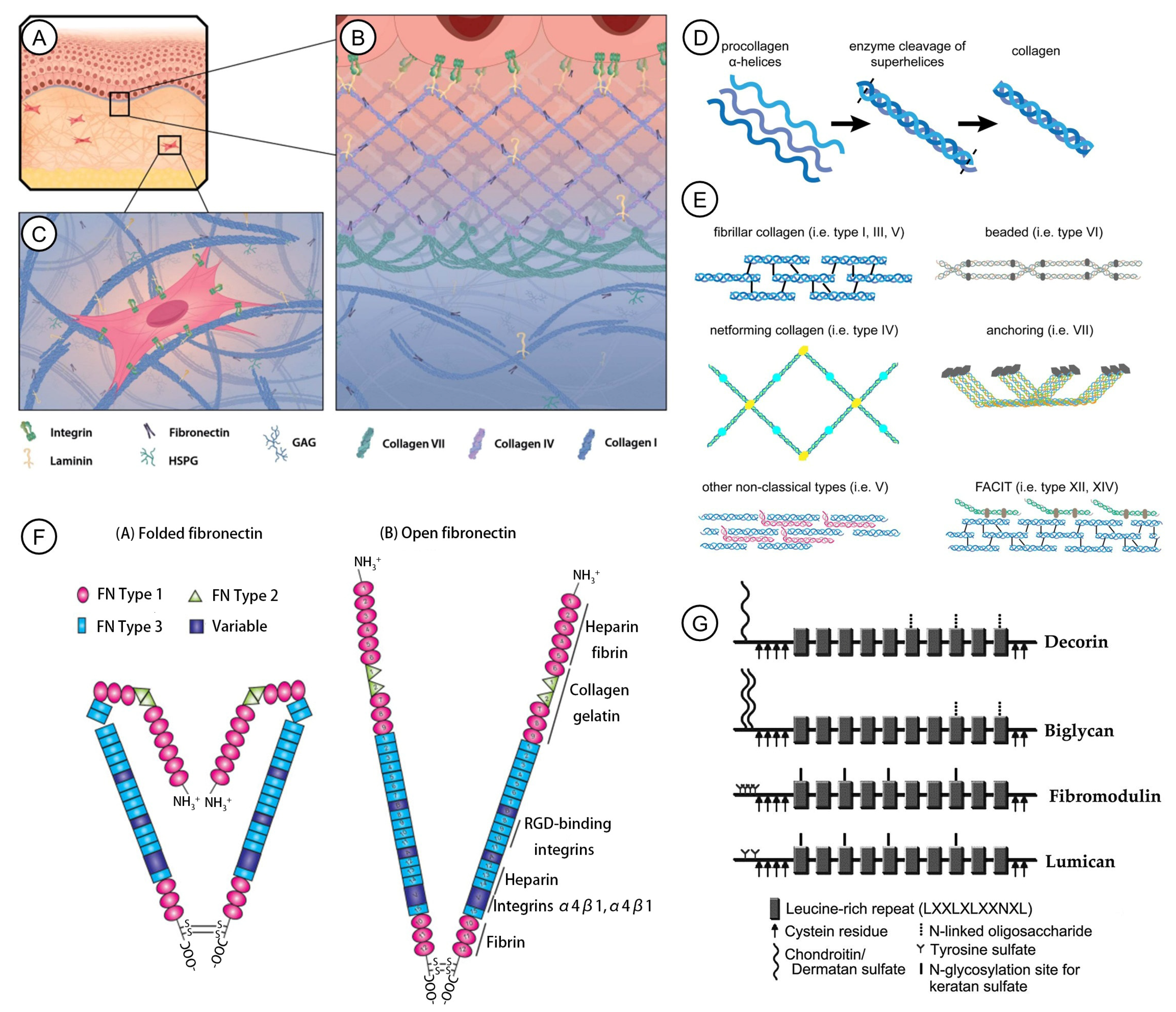
1.3. Elastin
1.4. Laminins
1.5. Other Molecules
2. Nano-Structures of ECM and Their Applications in Tissue Repair
3. Importance of NECM
4. Origins, Degradability, and Physical Properties
4.1. Origins
4.2. Degradability
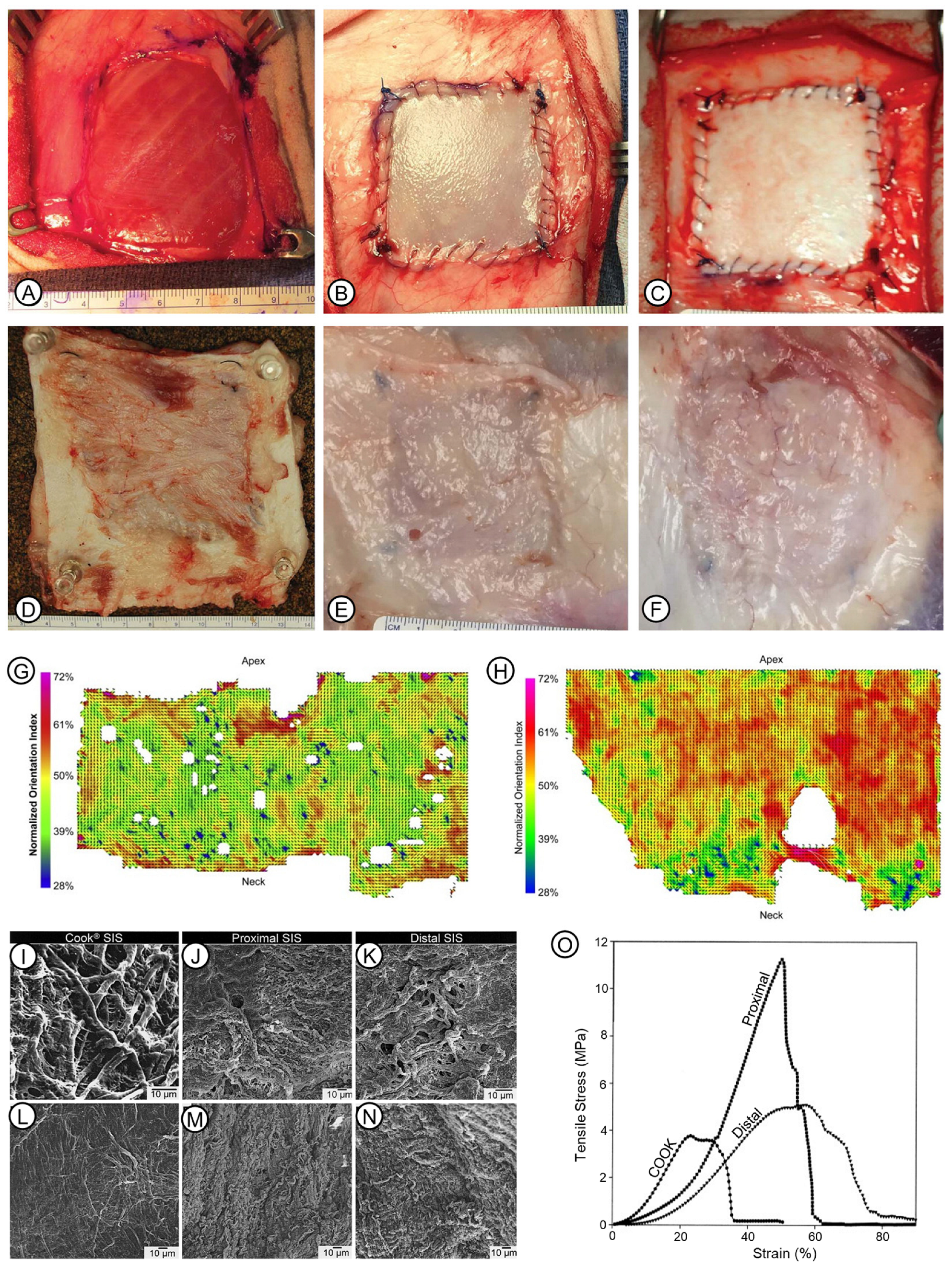
4.3. Physical Properties
5. Immune Response to NECMs
5.1. Innate Immune Response
5.2. Adaptive Immune Response
6. Clinical Applications
6.1. UBM
6.1.1. Wound Healing and Skin Repair
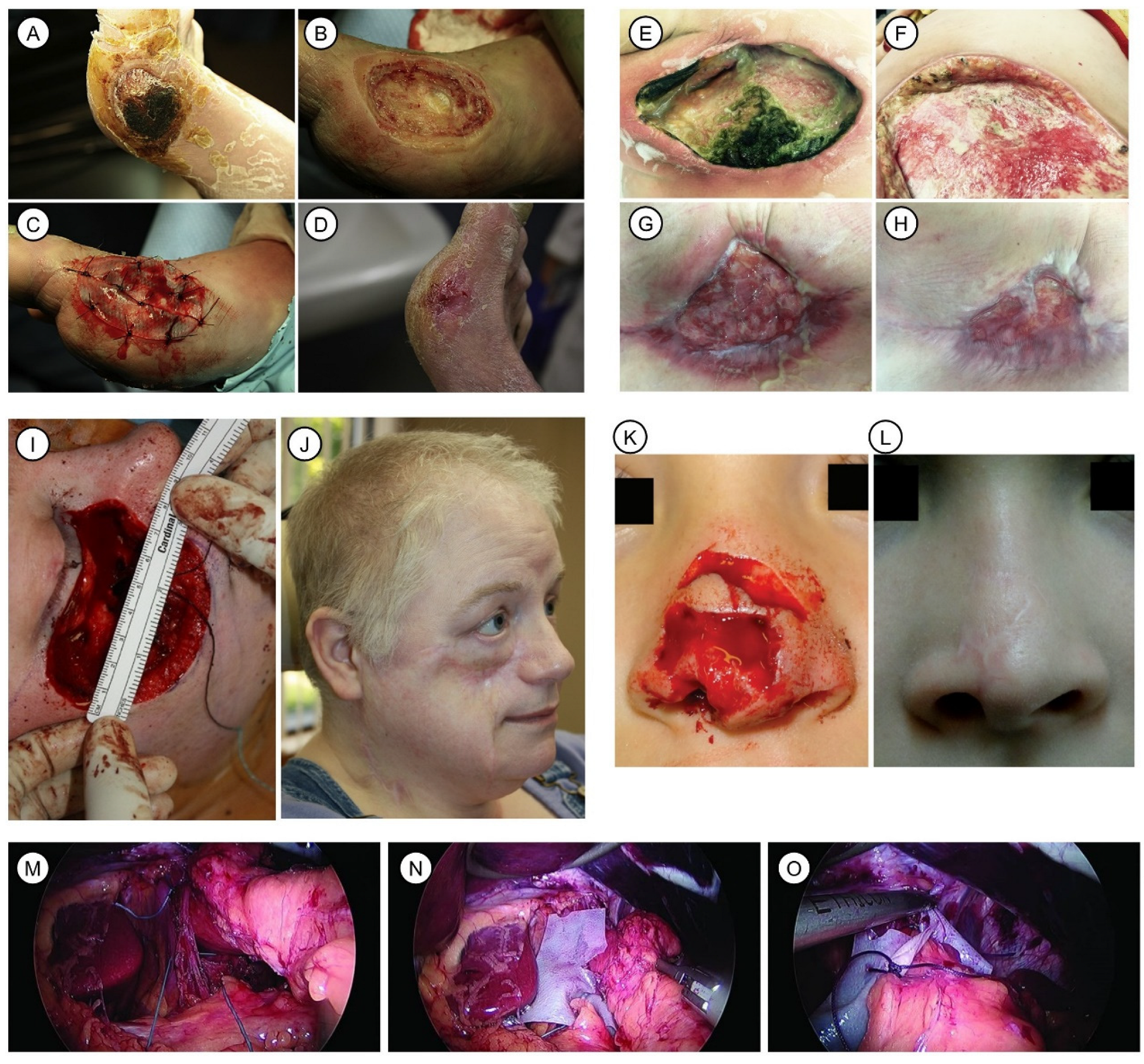
6.1.2. Plastic and Surgical Reconstruction
6.1.3. Hernia
6.2. Porcine Small Intestinal Submucosa (SIS)
6.2.1. Wound Healing and Skin Repair
6.2.2. Hernia
6.2.3. Defects in Other Tissues and Organs
6.3. ADM Derived from Human Cadaver Skin, Fetal Bovine Dermis, and Porcine Dermis
6.3.1. Breast Reconstruction
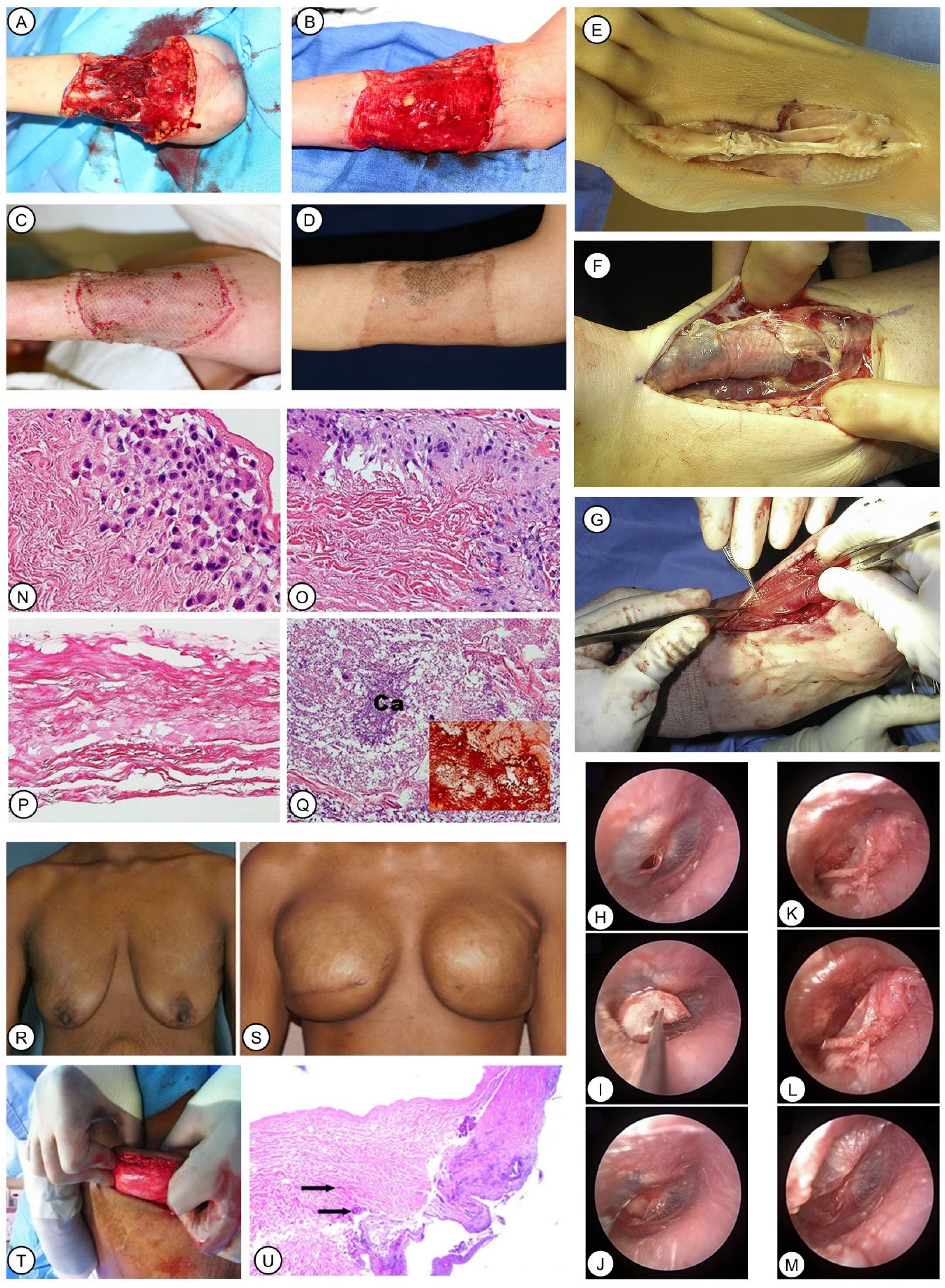
6.3.2. Wounds
6.3.3. Reconstructive Gynecology
6.3.4. Hernia
6.3.5. Other Defects
7. Indications, Features, and Clinical Relevance of Some Marketed NECM Devices
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Engel, J.; Chique, M. An overview of extracellular matrix structure and function. In The Extracellular Matrix: An Overview; Mecham, R.P., Ed.; Springer Nature: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Bowers, S.L.; Banerjee, I.; Baudino, T.A. The extracellular matrix: At the center of it all. J. Mol. Cell. Cardiol. 2010, 48, 474–482. [Google Scholar] [CrossRef]
- Abdulghani, S.; Mitchell, G.R. Biomaterials for in situ tissue regeneration: A review. Biomolecules 2019, 9, 750. [Google Scholar] [CrossRef]
- Shao, X.; Gomez, C.D.; Kapoor, N.; Considine, J.M.; Grams, C.; Gao, Y.; Naba, A. MatrisomeDB 2.0: 2023 updates to the ECM-protein knowledge database. Nucleic Acids Res. 2023, 51, D1519–D1530. [Google Scholar] [CrossRef] [PubMed]
- Esko, J.D.; Kimata, K.; Lindahl, U. Proteoglycans and Sulfated Glycosaminoglycans. In Essentials of Glycobiology; Varki, A., Cummings, R.D., Esko, J.D., Freeze, H.H., Stanley, P., Bertozzi, C.R., Hart, G.W., Etzler, M.E., Eds.; Cold Spring Harbor: New York, NY, USA, 2009. [Google Scholar]
- Schaefer, L.; Schaefer, R.M. Proteoglycans: From structural compounds to signaling molecules. Cell Tissue Res. 2010, 339, 237–246. [Google Scholar] [CrossRef]
- Pfisterer, K.; Shaw, L.E.; Symmank, D.; Weninger, W. The Extracellular Matrix in Skin Inflammation and Infection. Front. Cell Dev. Biol. 2021, 9, 682414. [Google Scholar] [CrossRef]
- Fassett, J.; Tobolt, D.; Hansen, L.K. Type I collagen structure regulates cell morphology and EGF signaling in primary rat hepatocytes through cAMP-dependent protein kinase A. Mol. Biol. Cell 2006, 17, 345–356. [Google Scholar] [CrossRef]
- Brodsky, B.; Persikov, A.V. Molecular structure of the collagen triple helix. Adv. Protein Chem. 2005, 70, 301–339. [Google Scholar]
- Gordon, M.K.; Hahn, R.A. Collagens. Cell Tissue Res. 2010, 339, 247–257. [Google Scholar] [CrossRef]
- Stylianou, A. Assessing Collagen D-Band Periodicity with Atomic Force Microscopy. Materials 2022, 15, 1608. [Google Scholar] [CrossRef]
- Wenstrup, R.J.; Florer, J.B.; Brunskill, E.W.; Bell, S.M.; Chervoneva, I.; Birk, D.E. Type V collagen controls the initiation of collagen fibril assembly. J. Biol. Chem. 2004, 279, 53331–53337. [Google Scholar] [CrossRef]
- To, W.S.; Midwood, K.S. Plasma and cellular fibronectin: Distinct and independent functions during tissue repair. Fibrogenesis Tissue Repair. 2011, 4, 21. [Google Scholar] [CrossRef]
- Drain, A.P.; Weaver, V.M. Chapter 7—Matrix molecules and their ligands. In Principles of Tissue Engineering, 5th ed.; Lanza, R., Langer, R., Vacanti, J.P., Atala, A., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 119–132. [Google Scholar]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef]
- Ni, G.X.; Li, Z.; Zhou, Y.Z. The role of small leucine-rich proteoglycans in osteoarthritis pathogenesis. Osteoarthr. Cartil. 2014, 22, 896–903. [Google Scholar] [CrossRef]
- Kozel, B.A.; Mecham, R.P. Elastic fiber ultrastructure and assembly. Matrix Biol. 2019, 84, 31–40. [Google Scholar] [CrossRef]
- Sun, B. The mechanics of fibrillar collagen extracellular matrix. Cell Rep. Phys. Sci. 2021, 2, 100515. [Google Scholar] [CrossRef]
- Wagenseil, J.E.; Mecham, R.P. New insights into elastic fiber assembly. Birth Defects Res. Part C Embryo Today Rev. 2007, 81, 229–240. [Google Scholar] [CrossRef]
- Ishihara, J.; Ishihara, A.; Fukunaga, K.; Sasaki, K.; White MJ, V.; Briquez, P.S.; Hubbell, J.A. Laminin heparin-binding peptides bind to several growth factors and enhance diabetic wound healing. Nat. Commun. 2018, 9, 2163. [Google Scholar] [CrossRef]
- Taniguchi, Y.; Ido, H.; Sanzen, N.; Hayashi, M.; Sato-Nishiuchi, R.; Futaki, S.; Sekiguchi, K. The C-terminal Region of Laminin β Chains Modulates the Integrin Binding Affinities of Laminins. J. Biol. Chem. 2009, 284, 7820–7831. [Google Scholar] [CrossRef]
- Bandzerewicz, A.; Gadomska-Gajadhur, A. Into the Tissues: Extracellular Matrix and Its Artificial Substitutes: Cell Signalling Mechanisms. Cells 2022, 11, 914. [Google Scholar] [CrossRef]
- Chen, S.; Birk, D.E. The regulatory roles of small leucine-rich proteoglycans in extracellular matrix assembly. FEBS J. 2013, 280, 2120–2137. [Google Scholar] [CrossRef]
- Sarrazin, S.; Lamanna, W.C.; Esko, J.D. Heparan sulfate proteoglycans. Cold Spring Harb. Perspect. Biol. 2011, 3, a004952. [Google Scholar] [CrossRef]
- Gubbiotti, M.A.; Neill, T.; Iozzo, R.V. A current view of perlecan in physiology and pathology: A mosaic of functions. Matrix Biol. 2017, 57–58, 285–298. [Google Scholar] [CrossRef]
- Guilak, F.; Hayes, A.J.; Melrose, J. Perlecan in Pericellular Mechanosensory Cell-Matrix Communication, Extracellular Matrix Stabilisation and Mechanoregulation of Load-Bearing Connective Tissues. Int. J. Mol. Sci. 2021, 22, 2716. [Google Scholar] [CrossRef]
- Burden, S.J.; Yumoto, N.; Zhang, W. The role of MuSK in synapse formation and neuromuscular disease. Cold Spring Harb. Perspect. Biol. 2013, 5, a009167. [Google Scholar] [CrossRef]
- Pozzi, A.; Yurchenco, P.D.; Iozzo, R.V. The nature and biology of basement membranes. Matrix Biol. 2017, 57–58, 1–11. [Google Scholar] [CrossRef]
- Chakraborty, S.; Sampath, D.; Lin, M.O.Y.; Bilton, M.; Huang, C.K.; Nai, M.H.; Njah, K.; Goy, P.A.; Wang, C.C.; Guccione, E.; et al. Agrin-Matrix Metalloproteinase-12 axis confers a mechanically competent microenvironment in skin wound healing. Nat. Commun. 2021, 12, 6349. [Google Scholar] [CrossRef]
- Tonti, O.R.; Larson, H.; Lipp, S.N.; Luetkemeyer, C.M.; Makam, M.; Vargas, D.; Wilcox, S.M.; Calve, S. Tissue-specific parameters for the design of ECM-mimetic biomaterials. Acta Biomater. 2021, 132, 83–102. [Google Scholar] [CrossRef]
- Van Smeden, J.; Janssens, M.; Gooris, G.S.; Bouwstra, J. The important role of stratum corneum lipids for the cutaneous barrier function. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2014, 1841, 295–313. [Google Scholar] [CrossRef]
- Skotland, T.; Sagini, K.; Sandvig, K.; Llorente, A. An emerging focus on lipids in extracellular vesicles. Adv. Drug Deliv. Rev. 2020, 159, 308–321. [Google Scholar] [CrossRef]
- Boskey, A.L.; Robey, P.G. The regulatory role of matrix proteins in mineralization of bone. In Osteoporosis; Academic Press: Cambridge, MA, USA, 2013; pp. 235–255. [Google Scholar]
- Gholami, A.; Hashemi, S.A.; Yousefi, K.; Mousavi, S.M.; Chiang, W.H.; Ramakrishna, S.; Mazraedoost, S.; Alizadeh, A.; Omidifar, N.; Babapoor, A. 3D nanostructures for tissue engineering, cancer therapy, and gene delivery. J. Nanomater. 2020, 2020, 1852946. [Google Scholar] [CrossRef]
- Singh, J.P.; Young, J.L. The cardiac nanoenvironment: Form and function at the nanoscale. Biophys. Rev. 2021, 13, 625–636. [Google Scholar] [CrossRef]
- Jara, C.P.; Wang, O.; do Prado, T.P.; Ismail, A.; Fabian, F.M.; Li, H.; Araújo, E.P. Novel fibrin-fibronectin matrix accelerates mice skin wound healing. Bioact. Mater. 2020, 5, 949–962. [Google Scholar] [CrossRef]
- Wallace, J.M.; Chen, Q.; Fang, M.; Erickson, B.; Orr, B.G.; Banaszak Holl, M.M. Type I collagen exists as a distribution of nanoscale morphologies in teeth, bones, and tendons. Langmuir 2010, 26, 7349–7354. [Google Scholar] [CrossRef]
- Kshitiz Afzal, J.; Kim, S.Y.; Kim, D.H. A nanotopography approach for studying the structure-function relationships of cells and tissues. Cell Adhes. Migr. 2015, 9, 300–307. [Google Scholar] [CrossRef]
- Stevens, M.M.; George, J.H. Exploring and engineering the cell surface interface. Science 2005, 3105751, 1135–1138. [Google Scholar] [CrossRef]
- Wei, G.; Ma, P.X. Nanostructured biomaterials for regeneration. Adv. Funct. Mater. 2008, 18, 3568–3582. [Google Scholar] [CrossRef]
- Maynard, S.A.; Gelmi, A.; Skaalure, S.C.; Pence, I.J.; Lee-Reeves, C.; Sero, J.E.; Stevens, M.M. Nanoscale molecular quantification of stem cell–hydrogel interactions. ACS Nano 2020, 14, 17321–17332. [Google Scholar] [CrossRef]
- Hynes, R.O. Integrins: Bidirectional, allosteric signaling machines. Cell 2002, 110, 673–687. [Google Scholar] [CrossRef]
- Szymanski, J.M.; Sevcik, E.N.; Zhang, K.; Feinberg, A.W. Stretch-dependent changes in molecular conformation in fibronectin nanofibers. Biomater. Sci. 2017, 5, 1629–1639. [Google Scholar] [CrossRef]
- Dzamba, B.J.; Peters, D.M. Arrangement of cellular fibronectin in noncollagenous fibrils in human fibroblast cultures. J. Cell Sci. 1991, 100, 605–612. [Google Scholar] [CrossRef]
- Smith, I.O.; Liu, X.H.; Smith, L.A.; Ma, P.X. Nanostructured polymer scaffolds for tissue engineering and regenerative medicine. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2009, 1, 226–236. [Google Scholar] [CrossRef]
- Yao, T.; Baker, M.B.; Moroni, L. Strategies to improve nanofibrous scaffolds for vascular tissue engineering. Nanomaterials 2020, 10, 887. [Google Scholar] [CrossRef]
- Woo, K.M.; Chen, V.J.; Ma, P.X. Nano-fibrous scaffolding architecture selectively enhances protein adsorption contributing to cell attachment. J. Biomed. Mater. Res. Part A Off. J. Soc. Biomater. Jpn. Soc. Biomater. Aust. Soc. Biomater. Korean Soc. Biomater. 2003, 67, 531–537. [Google Scholar] [CrossRef]
- Badylak, S.; Liang, A.; Record, R.; Tullius, R.; Hodde, J. Endothelial cell adherence to small intestinal submucosa: An acellular bioscaffold. Biomaterials 1999, 20, 2257–2263. [Google Scholar] [CrossRef]
- Woo, K.M.; Jun, J.H.; Chen, V.J.; Seo, J.; Baek, J.H.; Ryoo, H.M.; Ma, P.X. Nano-fibrous scaffolding promotes osteoblast differentiation and biomineralization. Biomaterials 2007, 28, 335–343. [Google Scholar] [CrossRef]
- Chen, V.J.; Smith, L.A.; Ma, P.X. Bone regeneration on computer-designed nano-fibrous scaffolds. Biomaterials 2006, 27, 3973–3979. [Google Scholar] [CrossRef]
- Li, W.J.; Danielson, K.G.; Alexander, P.G.; Tuan, R.S. Biological response of chondrocytes cultured in three-dimensional nanofibrous poly (ϵ-caprolactone) scaffolds. J. Biomed. Mater. Res. Part A Off. J. Soc. Biomater. Jpn. Soc. Biomater. Aust. Soc. Biomater. Korean Soc. Biomater. 2003, 67, 1105–1114. [Google Scholar]
- Shih YR, V.; Chen, C.N.; Tsai, S.W.; Wang, Y.J.; Lee, O.K. Growth of mesenchymal stem cells on electrospun type I collagen nanofibers. Stem Cells 2006, 24, 2391–2397. [Google Scholar] [CrossRef]
- Silva, G.A.; Czeisler, C.; Niece, K.L.; Beniash, E.; Harrington, D.A.; Kessler, J.A.; Stupp, S.I. Selective differentiation of neural progenitor cells by high-epitope density nanofibers. Science 2004, 3035662, 1352–1355. [Google Scholar] [CrossRef]
- Kim, B.-S.; Park, I.-K.; Hoshiba, T.; Jiang, H.-L.; Choi, Y.-J.; Akaike, T.; Cho, C.-S. Design of artificial extracellular matrices for tissue engineering. Prog. Polym. Sci. 2011, 36, 238–268. [Google Scholar] [CrossRef]
- Geckil, H.; Xu, F.; Zhang, X.; Moon, S.; Demirci, U. Engineering hydrogels as extracellular matrix mimics. Nanomedicine 2010, 5, 469–484. [Google Scholar] [CrossRef]
- Mason, B.N.; Reinhart-King, C.A. Controlling the mechanical properties of three-dimensional matrices via non-enzymatic collagen glycation. Organogenesis 2013, 9, 70–75. [Google Scholar] [CrossRef]
- Neishabouri, A.; Soltani Khaboushan, A.; Daghigh, F.; Kajbafzadeh, A.M.; Majidi Zolbin, M. Decellularization in Tissue Engineering and Regenerative Medicine: Evaluation, Modification, and Application Methods. Front. Bioeng. Biotechnol. 2022, 10, 805299. [Google Scholar] [CrossRef]
- Jun, I.; Han, H.S.; Edwards, J.R.; Jeon, H. Electrospun Fibrous Scaffolds for Tissue Engineering: Viewpoints on Architecture and Fabrication. Int. J. Mol. Sci. 2018, 19, 745. [Google Scholar] [CrossRef]
- Brown, B.N.; Badylak, S.F. Extracellular matrix as an inductive scaffold for functional tissue reconstruction. Transl. Res. 2014, 163, 268–285. [Google Scholar] [CrossRef]
- Billiet, T.; Vandenhaute, M.; Schelfhout, J.; Van Vlierberghe, S.; Dubruel, P. A review of trends and limitations in hydrogel-rapid prototyping for tissue engineering. Biomaterials 2012, 33, 6020–6041. [Google Scholar] [CrossRef]
- Habanjar, O.; Diab-Assaf, M.; Caldefie-Chezet, F.; Delort, L. 3D Cell Culture Systems: Tumor Application, Advantages, and Disadvantages. Int. J. Mol. Sci. 2021, 22, 12200. [Google Scholar] [CrossRef]
- Badylak, S.F. The extracellular matrix as a biologic scaffold material. Biomaterials 2007, 28, 3587–3593. [Google Scholar] [CrossRef]
- He, M.; Callanan, A. Comparison of methods for whole-organ decellularization in tissue engineering of bioartificial organs. Tissue Eng. Part B Rev. 2013, 19, 194–208. [Google Scholar] [CrossRef]
- Zantop, T.; Gilbert, T.W.; Yoder, M.C.; Badylak, S.F. Extracellular matrix scaffolds are repopulated by bone marrow-derived cells in a mouse model of achilles tendon reconstruction. J. Orthop. Res. 2006, 24, 1299–1309. [Google Scholar] [CrossRef]
- Badylak, S.F.; Park, K.; Peppas, N.; McCabe, G.; Yoder, M. Marrow-derived cells populate scaffolds composed of xenogeneic extracellular matrix. Exp. Hematol. 2001, 29, 1310–1318. [Google Scholar] [CrossRef]
- Choi, J.S.; Kim, J.D.; Yoon, H.S.; Cho, Y.W. Full-thickness skin wound healing using human placenta-derived extracellular matrix containing bioactive molecules. Tissue Eng. Part A 2013, 19, 329–339. [Google Scholar] [CrossRef]
- Kim, Y.; Ko, H.; Kwon, I.K.; Shin, K. Extracellular Matrix Revisited: Roles in Tissue Engineering. Int. Neurourol. J. 2016, 20, S23–S29. [Google Scholar] [CrossRef]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the extracellular matrix in development and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 786–801. [Google Scholar] [CrossRef]
- Neill, J.; James, K.; Lineaweaver, W. Utilizing biologic assimilation of bovine fetal collagen in staged skin grafting. Ann. Plast. Surg. 2012, 68, 451–456. [Google Scholar] [CrossRef]
- Neill, J.S.; Lineaweaver, W.C. Tissue response to bovine fetal collagen extracellular matrix in full-thickness skin wounds. Am. J. Clin. Pathol. 2013, 140, 248–252. [Google Scholar] [CrossRef]
- Brown, B.N.; Londono, R.; Tottey, S.; Zhang, L.; Kukla, K.A.; Wolf, M.T.; Daly, K.A.; Reing, J.E.; Badylak, S.F. Macrophage phenotype as a predictor of constructive remodeling following the implantation of biologically derived surgical mesh materials. Acta Biomater. 2012, 8, 978–987. [Google Scholar] [CrossRef]
- Young, D.A.; McGilvray, K.C.; Ehrhart, N.; Gilbert, T.W. Comparison of in vivo remodeling of urinary bladder matrix and acellular dermal matrix in an ovine model. Regen. Med. 2018, 13, 759–773. [Google Scholar] [CrossRef]
- Tottey, S.; Johnson, S.A.; Crapo, P.M.; Reing, J.E.; Zhang, L.; Jiang, H.; Medberry, C.J.; Reines, B.; Badylak, S.F. The effect of source animal age upon extracellular matrix scaffold properties. Biomaterials 2011, 32, 128–136. [Google Scholar] [CrossRef]
- Sicari, B.M.; Johnson, S.A.; Siu, B.F.; Crapo, P.M.; Daly, K.A.; Jiang, H.; Medberry, C.J.; Tottey, S.; Turner, N.J.; Badylak, S.F. The effect of source animal age upon the in vivo remodeling characteristics of an extracellular matrix scaffold. Biomaterials 2012, 33, 5524–5533. [Google Scholar] [CrossRef]
- Morris, A.H.; Stamer, D.K.; Kyriakides, T.R. The host response to naturally-derived extracellular matrix biomaterials. In Seminars in Immunology; Academic Press: Cambridge, MA, USA, 2017; Volume 29, pp. 72–91. [Google Scholar]
- Schwartz, J.D.; Monea, S.; Marcus, S.G.; Patel, S.; Eng, K.; Galloway, A.C.; Mignatti, P.; Shamamian, P. Soluble factor(s) released from neutrophils activates endothelial cell matrix metalloproteinase-2. J. Surg. Res. 1998, 76, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, S.D. Elastolytic metalloproteinases produced by human mononuclear phagocytes. Potential roles in destructive lung disease. Am. J. Respir. Crit. Care Med. 1994, 150, S160–S164. [Google Scholar] [CrossRef] [PubMed]
- Nelson, A.R.; Fingleton, B.; Rothenberg, M.L.; Matrisian, L.M. Matrix metalloproteinases: Biologic activity and clinical implications. J. Clin. Oncol. 2000, 18, 1135. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Barrantes, S.; Bernardo, M.; Toth, M.; Fridman, R. Regulation of membrane type-matrix metalloproteinases. In Seminars in Cancer Biology; Academic Press: Cambridge, MA, USA, 2002; Volume 12, pp. 131–138. [Google Scholar]
- Lopresti, S.T.; Brown, B.N. Host response to naturally derived biomaterials. In Host Response to Biomaterials; Academic Press: Cambridge, MA, USA, 2015; pp. 53–79. [Google Scholar]
- Carey, L.E.; Dearth, C.L.; Johnson, S.A.; Londono, R.; Medberry, C.J.; Daly, K.A.; Badylak, S.F. In vivo degradation of 14C-labeled porcine dermis biologic scaffold. Biomaterials 2014, 35, 8297–8304. [Google Scholar] [CrossRef] [PubMed]
- Valentin, J.E.; Badylak, J.S.; McCabe, G.P.; Badylak, S.F. Extracellular matrix bioscaffolds for orthopaedic applications: A comparative histologic study. JBJS 2006, 88, 2673–2686. [Google Scholar] [CrossRef] [PubMed]
- Vorotnikova, E.; McIntosh, D.; Dewilde, A.; Zhang, J.; Reing, J.E.; Zhang, L.; Cordero, K.; Bedelbaeva, K.; Gourevitch, D.; Braunhut, S.J.; et al. Extracellular matrix-derived products modulate endothelial and progenitor cell migration and proliferation in vitro and stimulate regenerative healing in vivo. Matrix Biol. 2010, 29, 690–700. [Google Scholar] [CrossRef] [PubMed]
- Reing, J.E.; Zhang, L.; Myers-Irvin, J.; Cordero, K.E.; Freytes, D.O.; Heber-Katz, E.; Bedelbaeva, K.; McIntosh, D.; Dewilde, A.; Badylak, S.F.; et al. Degradation products of extracellular matrix affect cell migration and proliferation. Tissue Eng. Part A 2009, 15, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Huleihel, L.; Hussey, G.S.; Naranjo, J.D.; Zhang, L.; Dziki, J.L.; Turner, N.J.; Stolz, D.B.; Badylak, S.F. Matrix-bound nanovesicles within ECM bioscaffolds. Sci. Adv. 2016, 2, e1600502. [Google Scholar] [CrossRef]
- Sadtler, K.; Allen, B.W.; Estrellas, K.; Housseau, F.; Pardoll, D.M.; Elisseeff, J.H. The scaffold immune microenvironment: Biomaterial-mediated immune polarization in traumatic and nontraumatic applications. Tissue Eng. Part A 2017, 23, 1044–1053. [Google Scholar] [CrossRef]
- Dziki, J.L.; Sicari, B.M.; Wolf, M.T.; Cramer, M.C.; Badylak, S.F. Immunomodulation and mobilization of progenitor cells by extracellular matrix bioscaffolds for volumetric muscle loss treatment. Tissue Eng. Part A 2016, 22, 1129–1139. [Google Scholar] [CrossRef]
- Sicari, B.M.; Dziki, J.L.; Siu, B.F.; Medberry, C.J.; Dearth, C.L.; Badylak, S.F. The promotion of a constructive macrophage phenotype by solubilized extracellular matrix. Biomaterials 2014, 35, 8605–8612. [Google Scholar] [CrossRef]
- Gilbert, T.W.; Wognum, S.; Joyce, E.M.; Freytes, D.O.; Sacks, M.S.; Badylak, S.F. Collagen fiber alignment and biaxial mechanical behavior of porcine urinary bladder derived extracellular matrix. Biomaterials 2008, 29, 4775–4782. [Google Scholar] [CrossRef] [PubMed]
- Raghavan, D.; Kropp, B.P.; Lin, H.K.; Zhang, Y.; Cowan, R.; Madihally, S.V. Physical characteristics of small intestinal submucosa scaffolds are location-dependent. J. Biomed. Mater. Res. A 2005, 73, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Garcia, F.D.; Fischer, T.; Hayn, A.; Mierke, C.T.; Burgess, J.K.; Harmsen, M.C. A Beginner’s Guide to the Characterization of Hydrogel Microarchitecture for Cellular Applications. Gels 2022, 8, 535. [Google Scholar] [CrossRef] [PubMed]
- Dahms, S.E.; Piechota, H.J.; Dahiya, R.; Lue, T.F.; Tanagho, E.A. Composition and biomechanical properties of the bladder acellular matrix graft: Comparative analysis in rat, pig and human. Br. J. Urol. 1998, 82, 411–419. [Google Scholar] [CrossRef]
- Wells, H.C.; Sizeland, K.H.; Kirby, N.; Hawley, A.; Mudie, S.; Haverkamp, R.G. Collagen Fibril Structure and Strength in Acellular Dermal Matrix Materials of Bovine, Porcine, and Human Origin. ACS Biomater. Sci. Eng. 2015, 1, 1026–1038. [Google Scholar] [CrossRef]
- Wang, X.; Chung, L.; Hooks, J.; Maestas, D.R., Jr.; Lebid, A.; Andorko, J.I.; Elisseeff, J.H. Type 2 immunity induced by bladder extracellular matrix enhances corneal wound healing. Sci. Adv. 2021, 7, eabe2635. [Google Scholar] [CrossRef]
- Puccio, F.; Solazzo, M.; Marciano, P. Comparison of three different mesh materials in tension-free inguinal hernia repair: Prolene versus Vypro versus surgisis. Int. Surg. 2005, 90, S21–S23. [Google Scholar]
- Matuska, A.M.; McFetridge, P.S. The effect of terminal sterilization on structural and biophysical properties of a decellularized collagen-based scaffold; implications for stem cell adhesion. J. Biomed. Mater. Res. Part B Appl. Biomater. 2015, 103, 397–406. [Google Scholar] [CrossRef]
- Daly, K.A.; Brown, B.N.; Badylak, S.F. Regenerative Medicine and the Foreign Body Response. In Tissue Engineering in Regenerative Medicine; Humana Press: Totowa, NJ, USA, 2011; pp. 353–375. [Google Scholar]
- Londono, R.; Badylak, S.F. Biologic scaffolds for regenerative medicine: Mechanisms of in vivo remodeling. Ann. Biomed. Eng. 2015, 43, 577–592. [Google Scholar] [CrossRef]
- Dziki, J.; Badylak, S.; Yabroudi, M.; Sicari, B.; Ambrosio, F.; Stearns, K.; Turner, N.; Wyse, A.; Boninger, M.L.; Rubin, J.P.; et al. An acellular biologic scaffold treatment for volumetric muscle loss: Results of a 13-patient cohort study. NPJ Regen. Med. 2016, 1, 16008. [Google Scholar] [CrossRef] [PubMed]
- Valentin, J.E.; Stewart-Akers, A.M.; Gilbert, T.W.; Badylak, S.F. Macrophage participation in the degradation and remodeling of extracellular matrix scaffolds. Tissue Eng. Part A 2009, 15, 1687–1694. [Google Scholar] [CrossRef] [PubMed]
- Badylak, S.F.; Valentin, J.E.; Ravindra, A.K.; McCabe, G.P.; Stewart-Akers, A.M. Macrophage phenotype as a determinant of biologic scaffold remodeling. Tissue Eng. Part A 2008, 14, 1835–1842. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Kang, R.; Coyne, C.B.; Zeh, H.J.; Lotze, M.T. PAMPs and DAMPs: Signal 0s that spur autophagy and immunity. Immunol. Rev. 2012, 249, 158–175. [Google Scholar] [CrossRef] [PubMed]
- Daly, K.A.; Liu, S.; Agrawal, V.; Brown, B.N.; Johnson, S.A.; Medberry, C.J.; Badylak, S.F. Damage associated molecular patterns within xenogeneic biologic scaffolds and their effects on host remodeling. Biomaterials 2012, 33, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Hemmi, H.; Takeuchi, O.; Kawai, T.; Kaisho, T.; Sato, S.; Sanjo, H.; Matsumoto, M.; Hoshino, K.; Wagner, H.; Akira, S.; et al. A Toll-like receptor recognizes bacterial DNA. Nature 2000, 4086813, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, T.W.; Freund, J.M.; Badylak, S.F. Quantification of DNA in biologic scaffold materials. J. Surg. Res. 2009, 152, 135–139. [Google Scholar] [CrossRef]
- Keane, T.J.; Londono, R.; Turner, N.J.; Badylak, S.F. Consequences of ineffective decellularization of biologic scaffolds on the host response. Biomaterials 2012, 33, 1771–1781. [Google Scholar] [CrossRef]
- Aamodt, J.M.; Grainger, D.W. Extracellular matrix-based biomaterial scaffolds and the host response. Biomaterials 2016, 86, 68–82. [Google Scholar] [CrossRef]
- Fishman, J.M.; Wiles, K.; Wood, K.J. The acquired immune system response to biomaterials, including both naturally occurring and synthetic biomaterials. In Host Response to Biomaterials; Academic Press: Cambridge, MA, USA, 2015; pp. 151–187. [Google Scholar]
- Allman, A.J.; McPherson, T.B.; Badylak, S.F.; Merrill, L.C.; Kallakury, B.; Sheehan, C.; Raeder, R.H.; Metzger, D.W. Xenogeneic extracellular matrix grafts elicit a TH2-restricted immune response1. Transplantation 2001, 71, 1631–1640. [Google Scholar] [CrossRef]
- Allman, A.J.; McPherson, T.B.; Merrill, L.C.; Badylak, S.F.; Metzger, D.W. The Th2-restricted immune response to xenogeneic small intestinal submucosa does not influence systemic protective immunity to viral and bacterial pathogens. Tissue Eng. 2002, 8, 53–62. [Google Scholar] [CrossRef]
- Galili, U. Human anti-gal and anti-non-gal immune response to porcine tissue implants. In Host Response to Biomaterials; Academic Press: Cambridge, MA, USA, 2015; pp. 239–267. [Google Scholar]
- Daly, K.A.; Stewart-Akers, A.M.; Hara, H.; Ezzelarab, M.; Long, C.; Cordero, K.; Badylak, S.F. Effect of the αGal epitope on the response to small intestinal submucosa extracellular matrix in a nonhuman primate model. Tissue Eng. Part A 2009, 15, 3877–3888. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.L.; Wong, J.L.; Athanasiou, K.A.; Griffiths, L.G. Stepwise solubilization-based antigen removal for xenogeneic scaffold generation in tissue engineering. Acta Biomater. 2013, 9, 6492–6501. [Google Scholar] [CrossRef] [PubMed]
- Frykberg, R.G.; Cazzell, S.M.; Arroyo-Rivera, J.; Tallis, A.; Reyzelman, A.M.; Saba, F.; Warren, L.; Stouch, B.C.; Gilbert, T.W. Evaluation of tissue engineering products for the management of neuropathic diabetic foot ulcers: An interim analysis. J. Wound Care 2016, 25, S18–S25. [Google Scholar] [CrossRef] [PubMed]
- Parry, J.A.; Shannon, S.F.; Strage, K.E.; Hadeed, M.M.; Heare, A.; Stacey, S.; Hammerberg, E.M.; Mauffrey, C. Urinary Bladder Matrix Grafting: A Simple and Effective Alternative to Flap Coverage for Wounds in High-Risk Orthopaedic Trauma Patients. J. Orthop. Trauma 2022, 36, e152–e157. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, B.A.; Geiger, S.E.; Deigni, O.A.; Watson, J.T. Management of Open Lower Extremity Wounds with Concomitant Fracture Using a Porcine Urinary Bladder Matrix. Wounds 2016, 28, 387–394. [Google Scholar] [PubMed]
- Kimmel, H.; Rahn, M.; Gilbert, T.W. The Clinical Effectiveness in Wound Healing with Extracellular Matrix Derived from Porcine Urinary Bladder Matrix: A Case Series on Severe Chronic Wounds. J. Am. Coll. Certif. Wound Spec. 2010, 2, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Lanteri Parcells, A.; Abernathie, B.; Datiashvili, R. The use of urinary bladder matrix in the treatment of complicated open wounds. Wounds 2014, 26, 189–196. [Google Scholar]
- Geiger, S.E.; Deigni, O.A.; Watson, J.T.; Kraemer, B.A. Management of Open Distal Lower Extremity Wounds with Exposed Tendons Using Porcine Urinary Bladder Matrix. Wounds 2016, 28, 306–316. [Google Scholar]
- Lecheminant, J.; Field, C. Porcine urinary bladder matrix: A retrospective study and establishment of protocol. J. Wound Care 2012, 21, 476, 478–480, 482. [Google Scholar] [CrossRef]
- Puckett, Y.; Pham, T.; McReynolds, S.; Ronaghan, C.A. Porcine Urinary Bladder Matrix for Management of Infected Radiation Mastectomy Wound. Cureus 2017, 9, e1451. [Google Scholar] [CrossRef] [PubMed]
- Rommer, E.A.; Peric, M.; Wong, A. Urinary bladder matrix for the treatment of recalcitrant nonhealing radiation wounds. Adv. Skin. Wound Care 2013, 26, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Vangsness, K.L.; Khim, P.; Raghuram, A.C.; Park, S.Y.; Yu, R.; Wong, A.K. Urinary Bladder Matrix Improves Irradiated Wound Healing in a Murine Model. Ann. Plast. Surg. 2022, 88, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Underwood, P.; Cardinal, P.; Keller, E.; Goodfellow, R.; Scalea, T.; Henry, S.; Lauerman, M.H. Extending Limb Salvage After Fourth and Fifth Transmetatarsal Amputation in Diabetic Foot Infections Using ACell® Urinary Bladder Matrix. Am. Surg. 2020, 89, 3134820973730. [Google Scholar] [CrossRef] [PubMed]
- Kruper, G.J.; Vandegriend, Z.P.; Lin, H.S.; Zuliani, G.F. Salvage of failed local and regional flaps with porcine urinary bladder extracellular matrix aided tissue regeneration. Case Rep. Otolaryngol. 2013, 2013, 917183. [Google Scholar] [CrossRef] [PubMed]
- Ong, A.A.; Nagy, R.; Fincham, M.R.; Nagy, M.L. Outcomes of Dog Bite Avulsion Injury Reconstruction with Urinary Bladder Matrix. Otolaryngol.-Head Neck Surg. 2020, 163, 1166–1168. [Google Scholar] [CrossRef]
- Zografakis, J.; Johnston, G.; Haas, J.; Berbiglia, L.; Bedford, T.; Spear, J.; Dan, A.; Pozsgay, M. Urinary Bladder Matrix Reinforcement for Laparoscopic Hiatal Hernia Repair. JSLS 2018, 22, e2017.00060. [Google Scholar] [CrossRef]
- Sasse, K.C.; Gevorkian, J.; Lambin, R.; Afshar, R.; Gardner, A.; Mehta, A.; Lambin, J.H.; Shinagawa, A. Large Hiatal Hernia Repair with Urinary Bladder Matrix Graft Reinforcement and Concomitant Sleeve Gastrectomy. JSLS 2019, 23, e2018.00106. [Google Scholar] [CrossRef]
- Wang, C.Q.; Tran, T.; Montera, B.; Karlnoski, R.; Feldman, J.; Albrink, M.H.; Velanovich, V. Symptomatic, Radiological, and Quality of Life Outcome of Paraesophageal Hernia Repair with Urinary Bladder Extracellular Surgical Matrix: Comparison with Primary Repair. Surg. Laparosc. Endosc. Percutan Tech. 2019, 29, 182–186. [Google Scholar] [CrossRef]
- Somani, A.K. Application expansion of small intestinal submucosa extracellular matrix in complex and surgical wounds. J. Wound Care 2023, 32, S20–S27. [Google Scholar] [CrossRef]
- Salgado, R.M.; Bravo, L.; García, M.; Melchor, J.M.; Krötzsch, E. Histomorphometric analysis of early epithelialization and dermal changes in mid-partial-thickness burn wounds in humans treated with porcine small intestinal submucosa and silver-containing hydrofiber. J. Burn. Care Res. 2014, 35, e330–e337. [Google Scholar] [CrossRef] [PubMed]
- Collini, F.J.; Stevenson, S.C.; Hodde, J.P. The bolster technique utilising small intestinal submucosa wound matrix: A novel approach to wound treatment. Int. Wound J. 2019, 16, 1222–1229. [Google Scholar] [CrossRef] [PubMed]
- Oelschlager, B.K.; Pellegrini, C.A.; Hunter, J.; Soper, N.; Brunt, M.; Sheppard, B.; Jobe, B.; Polissar, N.; Mitsumori, L.; Nelson, J.; et al. Biologic prosthesis reduces recurrence after laparoscopic paraesophageal hernia repair: A multicenter, prospective, randomized trial. Ann. Surg. 2006, 244, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Franklin, M.E.; Treviño, J.M., Jr.; Portillo, G.; Vela, I.; Glass, J.L.; González, J.J. The use of porcine small intestinal submucosa as a prosthetic material for laparoscopic hernia repair in infected and potentially contaminated fields: Long-term follow-up. Surg. Endosc. 2008, 22, 1941–1946. [Google Scholar] [CrossRef]
- Jafari, M.; Schneider-Bordat, L.; Hersant, B. Biological mesh used to repair perineal hernias following abdominoperineal resection for anorectal cancer. Ann. Chir. Plast. Esthet. 2020, 65, e15–e21. [Google Scholar] [CrossRef] [PubMed]
- Bibbo, C. The porcine small intestinal submucosa (SIS) patch in foot and ankle reconstruction. J. Foot Ankle Surg. 2010, 49, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Basonbul, R.A.; Cohen, M.S. Use of porcine small intestinal submucosa for pediatric endoscopic tympanic membrane repair. World J. Otorhinolaryngol.-Head Neck Surg. 2017, 3, 142–147. [Google Scholar] [CrossRef]
- Ballesteros, N.; Moscardi, P.R.M.; Blachman-Braun, R.; Salvitti, M.; Alam, A.; Castellan, M.; Kozakowski, K.; Gosalbez, R.; Labbie, A. Use of small intestinal submucosa for corporal body grafting in cases of epispadias and epispadias/exstrophy complex. J. Pediatr. Urol. 2019, 15, e401–e406. [Google Scholar] [CrossRef]
- Babu, R.; Chandrasekharam VV, S. A meta-analysis comparing dorsal plication and ventral lengthening for chordee correction during primary proximal hypospadias repair. Pediatr. Surg. Int. 2022, 38, 389–398. [Google Scholar] [CrossRef]
- Cosentino, M.; Kanashiro, A.; Vives, A.; Sanchez, J.; Peraza, M.F.; Moreno, D.; Perona, J.; De Marco, V.; Ruiz-Castañe, E.; Sarquella, J. Surgical treatment of Peyronie’s disease with small intestinal submucosa graft patch. Int. J. Impot. Res. 2016, 28, 106–109. [Google Scholar] [CrossRef]
- Weis, J.; Geiger, R.; Kilo, J.; Zimpfer, D. Cormatrix® for vessel reconstruction in paediatric cardiac surgery—A word of caution. Interact. Cardiovasc. Thorac. Surg. 2022, 34, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Rego, A.; Cheung, P.C.; Harris, W.J.; Brady, K.M.; Newman, J.; Still, R. Pericardial closure with extracellular matrix scaffold following cardiac surgery associated with a reduction of postoperative complications and 30-day hospital readmissions. J. Cardiothorac. Surg. 2019, 14, 61. [Google Scholar] [CrossRef] [PubMed]
- Woo, J.S.; Fishbein, M.C.; Reemtsen, B. Histologic examination of decellularized porcine intestinal submucosa extracellular matrix (CorMatrix) in pediatric congenital heart surgery. Cardiovasc. Pathol. 2016, 25, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Schnoeller, T.J.; de Petriconi, R.; Hefty, R.; Jentzmik, F.; Al Ghazal, A.; Steinestel, J.; Mueller, J.; Zengerling, F.; Schrader, M.; Schrader, A.J. The use of Surgisis® optimizes and simplifies partial nephrectomy for large renal tumors. Urol. A 2013, 52, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Ding, J.; Li, Y.; Hua, K.; Zhang, X. Comparison of two different methods for cervicovaginal reconstruction: A long-term follow-up. Int. Urogynecol. J. 2023, 34, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Badylak, S.; Kokini, K.; Tullius, B.; Simmons-Byrd, A.; Morff, R. Morphologic study of small intestinal submucosa as a body wall repair device. J. Surg. Res. 2002, 103, 190–202. [Google Scholar] [CrossRef] [PubMed]
- Rawlani, V.; Buck, D.W., 2nd; Johnson, S.A.; Heyer, K.S.; Kim, J.Y. Tissue expander breast reconstruction using prehydrated human acellular dermis. Ann. Plast. Surg. 2011, 66, 593–597. [Google Scholar] [CrossRef] [PubMed]
- Tevlin, R.; Borrelli, M.R.; Irizarry, D.; Nguyen, D.; Wan, D.C.; Momeni, A. Acellular Dermal Matrix Reduces Myofibroblast Presence in the Breast Capsule. Plast. Reconstr. Surg. Glob. Open 2019, 7, e2213. [Google Scholar] [CrossRef]
- Basu, C.B.; Leong, M.; Hicks, M.J. Acellular cadaveric dermis decreases the inflammatory response in capsule formation in reconstructive breast surgery. Plast. Reconstr. Surg. 2010, 126, 1842–1847. [Google Scholar] [CrossRef]
- Bullocks, J.M. DermACELL: A novel and biocompatible acellular dermal matrix in tissue expander and implant-based breast reconstruction. Eur. J. Plast. Surg. 2014, 37, 529–538. [Google Scholar] [CrossRef]
- Salzberg, C.A.; Dunavant, C.; Nocera, N. Immediate breast reconstruction using porcine acellular dermal matrix (Strattice™): Long-term outcomes and complications. J. Plast. Reconstr. Aesthet. Surg. 2013, 66, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Ricci, J.A.; Treiser, M.D.; Tao, R.; Jiang, W.; Guldbrandsen, G.; Halvorson, E.; Hergrueter, C.A.; Chun, Y.S. Predictors of Complications and Comparison of Outcomes Using SurgiMend Fetal Bovine and AlloDerm Human Cadaveric Acellular Dermal Matrices in Implant-Based Breast Reconstruction. Plast. Reconstr. Surg. 2016, 138, 583e–591e. [Google Scholar] [CrossRef] [PubMed]
- Cazzell, S. A Randomized Controlled Trial Comparing a Human Acellular Dermal Matrix Versus Conventional Care for the Treatment of Venous Leg Ulcers. Wounds 2019, 31, 68–74. [Google Scholar] [PubMed]
- Paredes, J.A.; Bhagwandin, S.; Polanco, T.; Lantis, J.C. Managing real world venous leg ulcers with fetal bovine acellular dermal matrix: A single centre retrospective study. J. Wound Care 2017, 26, S12–S19. [Google Scholar] [CrossRef] [PubMed]
- Lantis, J.C.; Snyder, R.; Reyzelman, A.M.; Van Gils, C.C.; Sigal, F.; Vayser, D.; Caporusso, J.M.; Cazzell, S.; Lavery, L.A. Fetal bovine acellular dermal matrix for the closure of diabetic foot ulcers: A prospective randomised controlled trial. J. Wound Care 2021, 30, S18–S27. [Google Scholar] [CrossRef] [PubMed]
- Kavros, S.J.; Dutra, T.; Gonzalez-Cruz, R.; Liden, B.; Marcus, B.; McGuire, J.; Nazario-Guirau, L. The use of PriMatrix, a fetal bovine acellular dermal matrix, in healing chronic diabetic foot ulcers: A prospective multicenter study. Adv. Skin. Wound Care 2014, 27, 356–362. [Google Scholar] [CrossRef]
- Ward, R.M.; Sung, V.W.; Clemons, J.L.; Myers, D.L. Vaginal paravaginal repair with an AlloDerm graft: Long-term outcomes. Am. J. Obstet. Gynecol. 2007, 197, 670.e1–670.e5. [Google Scholar] [CrossRef]
- Clemons, J.L.; Myers, D.L.; Aguilar, V.C.; Arya, L.A. Vaginal paravaginal repair with an AlloDerm graft. Am. J. Obstet. Gynecol. 2003, 189, 1612–1618. [Google Scholar] [CrossRef]
- Diaz, J.J.; Conquest, A.M., Jr.; Ferzoco, S.J.; Vargo, D.; Miller, P.; Wu, Y.C.; Donahue, R. Multi-institutional experience using human acellular dermal matrix for ventral hernia repair in a compromised surgical field. Arch. Surg. 2009, 144, 209–215. [Google Scholar] [CrossRef]
- Neumann, J.A.; Zgonis, M.H.; Rickert, K.D.; Bradley, K.E.; Kremen, T.J.; Boggess, B.R.; Toth, A.P. Interposition Dermal Matrix Xenografts: A Successful Alternative to Traditional Treatment of Massive Rotator Cuff Tears. Am. J. Sports Med. 2017, 45, 1261–1268. [Google Scholar] [CrossRef]
- Sclafani, A.P.; Romo, T., III; Jacono, A.A. Rejuvenation of the aging lip with an injectable acellular dermal graft (Cymetra). Arch. Facial Plast. Surg. 2002, 4, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Alei, G.; Letizia, P.; Ricottilli, F.; Simone, P.; Alei, L.; Massoni, F.; Ricci, S. Original technique for penile girth augmentation through porcine dermal acellular grafts: Results in a 69-patient series. J. Sex. Med. 2012, 9, 1945–1953. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of NECM Graft | Abbreviation | Animal Species Source | Source Tissue |
|---|---|---|---|
| urinary bladder matrix | UBM | porcine | the base membrane and lamina propria of urinary bladders |
| small intestinal submucosa | SIS | porcine | the submucosa of the small intestine |
| fetal bovine acellular dermal matrix | bADM | fetal bovine | dermis |
| porcine acellular dermal matrix | pADM | porcine | dermis |
| human acellular dermal matrix | hADM | human | dermis |
| acellular human amniotic membrane | AHAM | human | amniotic membrane |
| bovine pericardium extracellular matrix | BP ECM | Bovine | pricardium |
| Type of NECM | Uni-Layer Thickness (mm) | Normalized Force at Rupture [10 N/mm2] | Strain |
|---|---|---|---|
| SIS [90] | 0.25 & | 3.76 & | 0.23 & |
| UBM [92] | 0.15 | 0.29 | 0.40 |
| hADM [93] | 1.11 | 17.7 | 0.45 # |
| bADM [93] | 1.76 | 23.8 | 0.48 # |
| pADM [93] | 1.57 | 21.0 | 0.31 # |
| Name | Source | Appearance | Trade or Marketed Names | Indications | Feature and Clinical Relevance |
|---|---|---|---|---|---|
| Urinary bladder matrix | Porcine bladder | Membrane | Cytal® wound/burn matrix |
|
|
| Urinary bladder matrix | Porcine bladder | Particle | MicroMatrix® micronized particles |
|
|
| Acellular dermal matrix | Fetal bovine dermis | Membrane | PriMatrix® dermal repair scaffold |
|
|
| Small intestinal submucosa | Porcine jejunum | Membrane | OASIS® Matrix Products; Biodesign® |
| |
| Acellular dermal matrix | Human cadaver skin | Membrane | AlloDermTM RTM |
|
|
| Acellular dermal matrix | Human cadaver skin | Particle | Cymetra |
|
|
| Amniotic allograft membrane | Human amniotic tissue | Membrane | AminoExcel® |
|
|
| Acellular dermal scaffold | Porcine dermis | Membrane | STRATTICE™ Reconstructive Tissue Matrix |
|
|
| Acellular dermal scaffold | Porcine dermis | Membrane | CollaMend® |
|
|
| Bovine pericardium | Bovine pericardium | Membrane | Collagen Solutions |
|
|
| Bioimplant for soft tissue repair | Horse pericardium | Membrane | OrthADAPTTM bioimplant |
|
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, P.; Yang, F.; Jia, X.; Xiao, Y.; Hua, C.; Xing, M.; Lyu, G. Extracellular Matrices as Bioactive Materials for In Situ Tissue Regeneration. Pharmaceutics 2023, 15, 2771. https://doi.org/10.3390/pharmaceutics15122771
Zhao P, Yang F, Jia X, Xiao Y, Hua C, Xing M, Lyu G. Extracellular Matrices as Bioactive Materials for In Situ Tissue Regeneration. Pharmaceutics. 2023; 15(12):2771. https://doi.org/10.3390/pharmaceutics15122771
Chicago/Turabian StyleZhao, Peng, Fengbo Yang, Xiaoli Jia, Yuqin Xiao, Chao Hua, Malcolm Xing, and Guozhong Lyu. 2023. "Extracellular Matrices as Bioactive Materials for In Situ Tissue Regeneration" Pharmaceutics 15, no. 12: 2771. https://doi.org/10.3390/pharmaceutics15122771
APA StyleZhao, P., Yang, F., Jia, X., Xiao, Y., Hua, C., Xing, M., & Lyu, G. (2023). Extracellular Matrices as Bioactive Materials for In Situ Tissue Regeneration. Pharmaceutics, 15(12), 2771. https://doi.org/10.3390/pharmaceutics15122771