
Therapeutic Effects of Oral Application of Menthol and Extracts from Tormentil (Potentilla erecta), Raspberry Leaves (Rubus idaeus), and Loosestrife (Lythrum salicaria) during Acute Murine Campylobacteriosis


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Chromatographic Analysis
2.3. Microbiota Depleted IL-10−/− Mice
2.4. Campylobacter Jejuni Infection
2.5. Compounds and Treatment Regimens
2.5.1. Tormentil
2.5.2. Raspberry Leaves
2.5.3. Loosestrife
2.5.4. Menthol
2.6. Gastrointestinal Pathogen Burdens
2.7. Monitoring of Clinical Conditions of Animals
2.8. Sampling
2.9. Histopathology
2.10. In Situ Immunohistochemistry
2.11. Pro-Inflammatory Cytokines
2.12. Statistics
3. Results
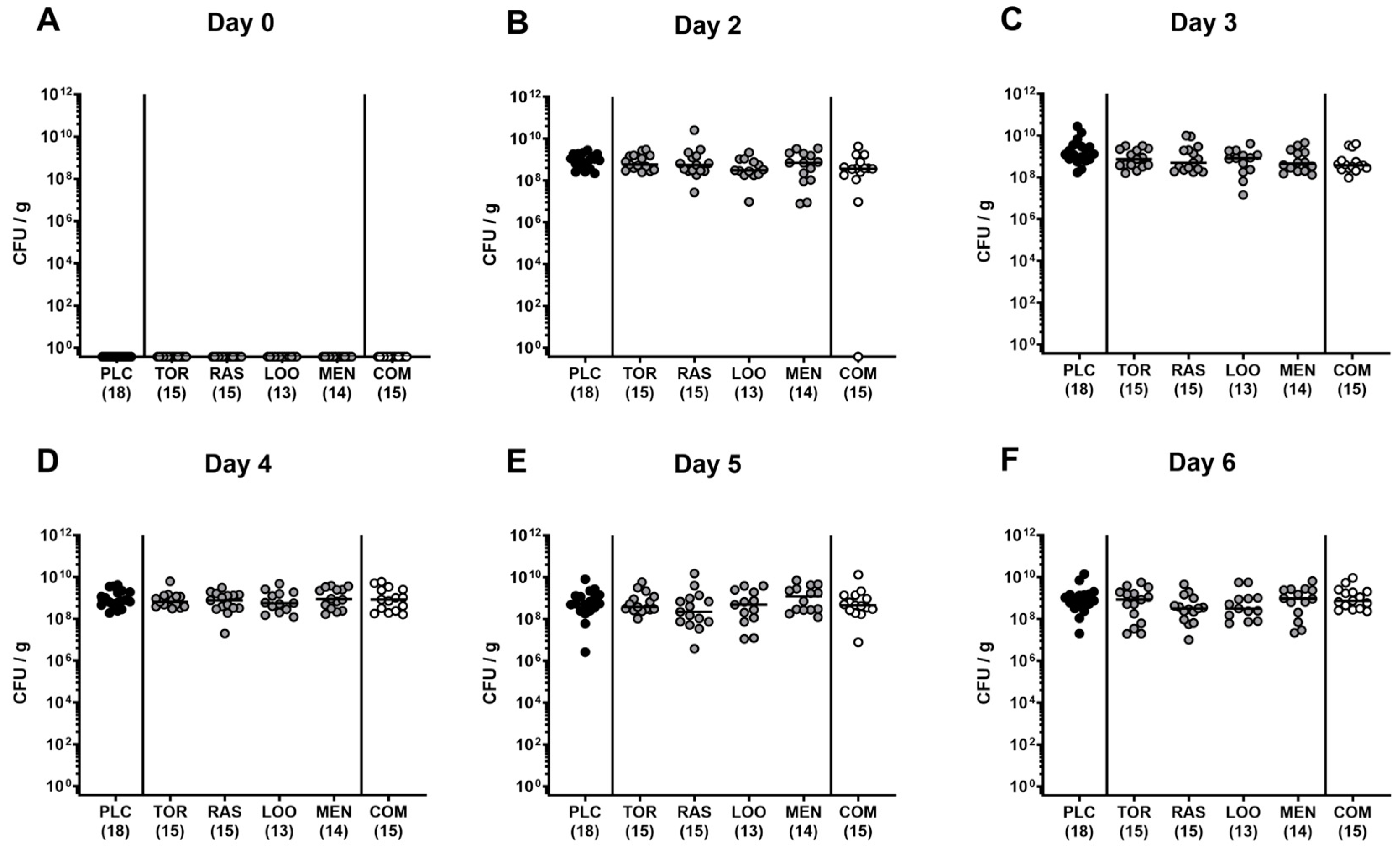
3.1. Gastrointestinal C. jejuni Numbers Following Oral Treatment of Infected Gut Microbiota-depleted IL-10−/− Mice with Menthol and Extracts Derived from Tormentil, Raspberry Leaves, and Loosestrife Alone or in Combination
3.2. Clinical Conditions over Time Following Oral Treatment of Infected Gut Microbiota-depleted IL-10−/− Mice with Distinct Natural Compounds Alone or in Combination
3.3. Inflammatory Signs Following Oral Treatment of Infected Gut Microbiota-Depleted IL-10−/− Mice with Distinct Natural Compounds Alone or in Combination
3.4. Colonic Immune Cell Responses Following Oral Treatment of Infected Gut Microbiota-Depleted IL-10−/− Mice with Distinct Natural Compounds Alone or in Combination
3.5. Intestinal Pro-Inflammatory Cytokine Secretion Following Oral Treatment of Gut Infected Microbiota-Depleted IL-10−/− Mice with Distinct Natural Compounds Alone or in Combination
3.6. C. jejuni Translocation to MLN Following Oral Treatment of Infected Gut Microbiota-Depleted IL-10−/− Mice with Distinct Natural Compounds Alone or in Combination
3.7. Extra-Intestinal Pro-Inflammatory Cytokine Secretion Following Oral Treatment of Infected Gut Microbiota-Depleted IL-10−/− Mice with Distinct Natural Compounds Alone or in Combination
4. Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract | Compound | Study | Summary of the Effects | Ref. |
|---|---|---|---|---|
| TOR RAS LOO | Ellagic acid | In vivo (mice) | Antibacterial activity against Helicobacter pylori | [83] |
| TNBS-induced colitis in rats | Downregulated iNOS mRNA (colon) Lower TNF-α concentrations (colon) | [82] | ||
| DSS-induced colitis in mice | Less intestinal inflammation Less diarrhea Attenuated histopathological changes (colon) Downregulated COX-2 and iNOS mRNA (colon) Lower IL-6, IFN-γ, TNF-α concentrations (colon) | [73] | ||
| TOR | Procyanidin | In vitro | Antibacterial activity against Campylobacter jejuni | [85] |
| In vitro | Antibacterial activity against Listeria monocytogenes | [86] | ||
| DSS-induced colitis in mice | Less severe colitis Decreased colonic infiltration with macrophages Downregulated IL-1β, TNF-α, IL-6 mRNA Suppressed NF-κB signaling | [87] | ||
| DSS-induced colitis in mice | Less severe colitis Reduced shortening of colonic length Decreased proinflammatory macrophages Downregulated IL-1β, IL-6, TNF-α, iNOS mRNA Upregulated TGF-β, CD206 mRNA | [88] | ||
| Gallic acid | DSS-induced colitis in mice | Reduced shortening of colonic length Enhanced reconstruction of microvilli (colon) Attenuated histopathological changes (colon) Downregulated TNF-α, IL-1β, IFN-γ, IL-6, IL-17 mRNA (colon) | [89] | |
| TNBS-induced colitis in mice | Attenuated histopathological changes (colon) Inhibited apoptosis (colon) Upregulated IL-4, IL-10 mRNA (colon) Downregulated IL-1, IL-6, IL-12, IL-17, IL-23, TGF-β, TNF-α mRNA (colon) Suppressed NF-κB signaling | [90] | ||
| Agrimoniin | In vitro | Antibacterial activity against Helicobacter pylori | [96] | |
| In vitro | Inhibited IL-8 secretion in TNF-α -treated human gastric epithelial cells Suppressed NF-κB signaling | [97] | ||
| RAS | Quercetin | In vitro | Antibacterial activity against Klebsiella pneumoniae, Pseudomonas aeruginosa, Yersinia enterocolitica | [91] |
| In vitro | Antibacterial activity against Vibrio parahaemolyticus | [92] | ||
| Acetic acid-induced colitis in mice | Attenuated histopathological changes (colon) Decreased recruitment of neutrophils (colon) Lower IL-1β, IL-33 concentrations (colon) Higher IL-10 concentrations (colon) | [93] | ||
| DSS-induced colitis in mice | Less severe colitis Attenuated histopathological changes (colon) Decreased IFN-γ+ and TNF-α+ CD4+ T cells (colon, MLN) Lower TNF-α, IFN-γ, IL-6 concentrations (colon) Upregulated IL-10 mRNA (colon) | [94] | ||
| LOO | Vescalagin and castalagin | In vitro | Antibactericidal activity against Staphylococcus epidermidis, Staphylococcus aureus, methicillin-resistant Staphylococcus aureus, and Pseudomonas aeruginosa | [95] |
| In vitro | Inhibited IL-8 production from human neutrophils | [49] |
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fitzgerald, C. Campylobacter. Clin. Lab. Med. 2015, 35, 289–298. [Google Scholar] [CrossRef] [PubMed]
- The European Union One Health 2021 Zoonoses Report. EFSA J. 2022, 20, e07666. [CrossRef]
- Vos, T.; Lim, S.S.; Abbafati, C.; Abbas, K.M.; Abbasi, M.; Abbasifard, M.; Abbasi-Kangevari, M.; Abbastabar, H.; Abd-Allah, F.; Abdelalim, A.; et al. Global burden of 369 diseases and injuries in 204 countries and territories, 1990–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1204–1222. [Google Scholar] [CrossRef] [PubMed]
- Berthenet, E.; Thépault, A.; Chemaly, M.; Rivoal, K.; Ducournau, A.; Buissonnière, A.; Bénéjat, L.; Bessède, E.; Mégraud, F.; Sheppard, S.K.; et al. Source attribution of Campylobacter jejuni shows variable importance of chicken and ruminants reservoirs in non-invasive and invasive French clinical isolates. Sci. Rep. 2019, 9, 8098. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.; Leite, D.; Fernandes, M.; Mena, C.; Gibbs, P.; Teixeira, P. Campylobacter spp. as a Foodborne Pathogen: A Review. Front. Microbiol. 2011, 2, 200. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, M.; Ahmed, S.; Barnes, A.N. A systematic review of waterborne and water-related disease in animal populations of Florida from 1999–2019. PLoS ONE 2021, 16, e0255025. [Google Scholar] [CrossRef] [PubMed]
- Igwaran, A.; Okoh, A.I. Human campylobacteriosis: A public health concern of global importance. Heliyon 2019, 5, e02814. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Sahin, O.; Grover, M.; Zhang, Q. New and alternative strategies for the prevention, control, and treatment of antibiotic-resistant Campylobacter. Transl. Res. 2020, 223, 76–88. [Google Scholar] [CrossRef]
- Finsterer, J. Triggers of Guillain-Barré Syndrome: Campylobacter jejuni Predominates. Int. J. Mol. Sci. 2022, 23, 14222. [Google Scholar] [CrossRef]
- Backert, S.; Tegtmeyer, N.; Cróinín, T.Ó.; Boehm, M.; Heimesaat, M.M. Chapter 1—Human campylobacteriosis. In Campylobacter; Klein, G., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 1–25. [Google Scholar] [CrossRef]
- Pogreba-Brown, K.; Austhof, E.; Armstrong, A.; Schaefer, K.; Villa Zapata, L.; McClelland, D.J.; Batz, M.B.; Kuecken, M.; Riddle, M.; Porter, C.K.; et al. Chronic Gastrointestinal and Joint-Related Sequelae Associated with Common Foodborne Illnesses: A Scoping Review. Foodborne Pathog. Dis. 2020, 17, 67–86. [Google Scholar] [CrossRef]
- Kaakoush, N.O.; Castano-Rodriguez, N.; Mitchell, H.M.; Man, S.M. Global Epidemiology of Campylobacter Infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef] [PubMed]
- Koga, M.; Gilbert, M.; Takahashi, M.; Li, J.; Koike, S.; Hirata, K.; Yuki, N. Comprehensive Analysis of Bacterial Risk Factors for the Development of Guillain-Barré Syndrome after Campylobacter jejuni Enteritis. J. Infect. Dis. 2006, 193, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, N.P.; Kuijf, M.L.; Ang, C.W.; Schiellerup, P.; Krogfelt, K.A.; Jacobs, B.C.; van Belkum, A.; Endtz, H.P.; Bergman, M.P. Sialylation of Campylobacter jejuni lipo-oligosaccharides is associated with severe gastro-enteritis and reactive arthritis. Microbes Infect. 2009, 11, 988–994. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, S.; Bereswill, S.; Heimesaat, M.M. Novel Clinical Campylobacter jejuni Infection Models Based on Sensitization of Mice to Lipooligosaccharide, a Major Bacterial Factor Triggering Innate Immune Responses in Human Campylobacteriosis. Microorganisms 2020, 8, 482. [Google Scholar] [CrossRef] [PubMed]
- Bereswill, S.; Fischer, A.; Plickert, R.; Haag, L.M.; Otto, B.; Kuhl, A.A.; Dasti, J.I.; Zautner, A.E.; Munoz, M.; Loddenkemper, C.; et al. Novel murine infection models provide deep insights into the "menage a trois" of Campylobacter jejuni, microbiota and host innate immunity. PLoS ONE 2011, 6, e20953. [Google Scholar] [CrossRef]
- Tegtmeyer, N.; Sharafutdinov, I.; Harrer, A.; Soltan Esmaeili, D.; Linz, B.; Backert, S. Campylobacter Virulence Factors and Molecular Host-Pathogen Interactions. Curr. Top. Microbiol. Immunol. 2021, 431, 169–202. [Google Scholar] [CrossRef] [PubMed]
- Heimesaat, M.M.; Alutis, M.; Grundmann, U.; Fischer, A.; Tegtmeyer, N.; Böhm, M.; Kühl, A.A.; Göbel, U.B.; Backert, S.; Bereswill, S. The role of serine protease HtrA in acute ulcerative enterocolitis and extra-intestinal immune responses during Campylobacter jejuni infection of gnotobiotic IL-10 deficient mice. Front. Cell. Infect. Microbiol. 2014, 4, 77. [Google Scholar] [CrossRef]
- Wagenaar, J.A.; French, N.P.; Havelaar, A.H. Preventing Campylobacter at the source: Why is it so difficult? Clin. Infect. Dis. 2013, 57, 1600–1606. [Google Scholar] [CrossRef]
- Poly, F.; Noll, A.J.; Riddle, M.S.; Porter, C.K. Update on Campylobacter vaccine development. Hum. Vaccin. Immunother. 2019, 15, 1389–1400. [Google Scholar] [CrossRef]
- Humphries, R.M.; Linscott, A.J. Practical Guidance for Clinical Microbiology Laboratories: Diagnosis of Bacterial Gastroenteritis. Clin. Microbiol. Rev. 2015, 28, 3–31. [Google Scholar] [CrossRef]
- Bereswill, S.; Mousavi, S.; Weschka, D.; Buczkowski, A.; Schmidt, S.; Heimesaat, M.M. Peroral Clove Essential Oil Treatment Ameliorates Acute Campylobacteriosis—Results from a Preclinical Murine Intervention Study. Microorganisms 2021, 9, 735. [Google Scholar] [CrossRef] [PubMed]
- Heimesaat, M.M.; Mousavi, S.; Weschka, D.; Bereswill, S. Garlic Essential Oil as Promising Option for the Treatment of Acute Campylobacteriosis—Results from a Preclinical Placebo-Controlled Intervention Study. Microorganisms 2021, 9, 1140. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, S.; Weschka, D.; Bereswill, S.; Heimesaat, M.M. Immune-Modulatory Effects upon Oral Application of Cumin-Essential-Oil to Mice Suffering from Acute Campylobacteriosis. Pathogens 2021, 10, 818. [Google Scholar] [CrossRef] [PubMed]
- Heimesaat, M.M.; Mousavi, S.; Weschka, D.; Bereswill, S. Anti-Pathogenic and Immune-Modulatory Effects of Peroral Treatment with Cardamom Essential Oil in Acute Murine Campylobacteriosis. Microorganisms 2021, 9, 169. [Google Scholar] [CrossRef] [PubMed]
- Heimesaat, M.M.; Mousavi, S.; Escher, U.; Lobo de Sá, F.D.; Peh, E.; Schulzke, J.-D.; Kittler, S.; Bücker, R.; Bereswill, S. Resveratrol Alleviates Acute Campylobacter jejuni Induced Enterocolitis in a Preclinical Murine Intervention Study. Microorganisms 2020, 8, 1858. [Google Scholar] [CrossRef] [PubMed]
- Lobo de Sa, F.D.; Butkevych, E.; Nattramilarasu, P.K.; Fromm, A.; Mousavi, S.; Moos, V.; Golz, J.C.; Stingl, K.; Kittler, S.; Seinige, D.; et al. Curcumin Mitigates Immune-Induced Epithelial Barrier Dysfunction by Campylobacter jejuni. Int. J. Mol. Sci. 2019, 20, 4830. [Google Scholar] [CrossRef] [PubMed]
- Bunse, M.; Daniels, R.; Gründemann, C.; Heilmann, J.; Kammerer, D.R.; Keusgen, M.; Lindequist, U.; Melzig, M.F.; Morlock, G.E.; Schulz, H.; et al. Essential Oils as Multicomponent Mixtures and Their Potential for Human Health and Well-Being. Front. Pharmacol. 2022, 13, 956541. [Google Scholar] [CrossRef] [PubMed]
- Tomczyk, M.; Latté, K.P. Potentilla—A review of its phytochemical and pharmacological profile. J. Ethnopharmacol. 2009, 122, 184–204. [Google Scholar] [CrossRef]
- Watkins, F.; Pendry, B.; Sanchez-Medina, A.; Corcoran, O. Antimicrobial assays of three native British plants used in Anglo-Saxon medicine for wound healing formulations in 10th century England. J. Ethnopharmacol. 2012, 144, 408–415. [Google Scholar] [CrossRef]
- Dróżdż, P.; Sentkowska, A.; Pyrzynska, K. Potentilla erecta (L.) rhizomes as a source of phenolic acids. Nat. Prod. Res. 2019, 33, 2128–2131. [Google Scholar] [CrossRef]
- Paduch, R.; Wiater, A.; Locatelli, M.; Pleszczyńska, M.; Tomczyk, M. Aqueous Extracts of Selected Potentilla Species Modulate Biological Activity of Human Normal Colon Cells. Curr. Drug Targets 2015, 16, 1495–1502. [Google Scholar] [CrossRef] [PubMed]
- Kaltalioglu, K.; Balabanli, B.; Coskun-Cevher, S. Phenolic, Antioxidant, Antimicrobial, and In-vivo Wound Healing Properties of Potentilla erecta L. Root Extract in Diabetic Rats. Iran. J. Pharm. Res. 2020, 19, 264–274. [Google Scholar] [CrossRef]
- Farnaghi, S.; Braniff, K. Survey of Raspberry Leaf Tea in pregnancy. Aust. New Zealand J. Obstet. Gynaecol. 2022, 62, 506–510. [Google Scholar] [CrossRef] [PubMed]
- Ferlemi, A.V.; Lamari, F.N. Berry Leaves: An Alternative Source of Bioactive Natural Products of Nutritional and Medicinal Value. Antioxidants 2016, 5, 17. [Google Scholar] [CrossRef] [PubMed]
- Piwowarski, J.P.; Granica, S.; Zwierzyńska, M.; Stefańska, J.; Schopohl, P.; Melzig, M.F.; Kiss, A.K. Role of human gut microbiota metabolism in the anti-inflammatory effect of traditionally used ellagitannin-rich plant materials. J. Ethnopharmacol. 2014, 155, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Staszowska-Karkut, M.; Materska, M. Phenolic Composition, Mineral Content, and Beneficial Bioactivities of Leaf Extracts from Black Currant (Ribes nigrum L.), Raspberry (Rubus idaeus), and Aronia (Aronia melanocarpa). Nutrients 2020, 12, 463. [Google Scholar] [CrossRef] [PubMed]
- Dudzinska, D.; Bednarska, K.; Boncler, M.; Luzak, B.; Watala, C. The influence of Rubus idaeus and Rubus caesius leaf extracts on platelet aggregation in whole blood. Cross-talk of platelets and neutrophils. Platelets 2016, 27, 433–439. [Google Scholar] [CrossRef]
- Krzepiłko, A.; Prażak, R.; Święciło, A. Chemical Composition, Antioxidant and Antimicrobial Activity of Raspberry, Blackberry and Raspberry-Blackberry Hybrid Leaf Buds. Molecules 2021, 26, 327. [Google Scholar] [CrossRef]
- Nikitina, V.S.; Kuz’mina, L.Y.; Melent’ev, A.I.; Shendel’, G.V. Antibacterial activity of polyphenolic compounds isolated from plants of Geraniaceae and Rosaceae families. Appl. Biochem. Microbiol. 2007, 43, 629–634. [Google Scholar] [CrossRef]
- Rojas-Vera, J.; Patel, A.V.; Dacke, C.G. Relaxant activity of raspberry (Rubus idaeus) leaf extract in guinea-pig ileum in vitro. Phytother. Res. 2002, 16, 665–668. [Google Scholar] [CrossRef]
- Puupponen-Pimiä, R.; Nohynek, L.; Hartmann-Schmidlin, S.; Kähkönen, M.; Heinonen, M.; Määttä-Riihinen, K.; Oksman-Caldentey, K.M. Berry phenolics selectively inhibit the growth of intestinal pathogens. J. Appl. Microbiol. 2005, 98, 991–1000. [Google Scholar] [CrossRef] [PubMed]
- Tabashsum, Z.; Peng, M.; Kahan, E.; Rahaman, S.O.; Biswas, D. Effect of conjugated linoleic acid overproducing Lactobacillus with berry pomace phenolic extracts on Campylobacter jejuni pathogenesis. Food Funct. 2019, 10, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Nohynek, L.J.; Alakomi, H.L.; Kähkönen, M.P.; Heinonen, M.; Helander, I.M.; Oksman-Caldentey, K.M.; Puupponen-Pimiä, R.H. Berry phenolics: Antimicrobial properties and mechanisms of action against severe human pathogens. Nutr. Cancer 2006, 54, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Tunalier, Z.; Koşar, M.; Küpeli, E.; Caliş, I.; Başer, K.H. Antioxidant, anti-inflammatory, anti-nociceptive activities and composition of Lythrum salicaria L. extracts. J. Ethnopharmacol. 2007, 110, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Vafi, F.; Bahramsoltani, R.; Abdollahi, M.; Manayi, A.; Hossein Abdolghaffari, A.; Samadi, N.; Amin, G.; Hassanzadeh, G.; Jamalifar, H.; Baeeri, M.; et al. Burn Wound Healing Activity of Lythrum salicaria L. and Hypericum scabrum L. Wounds 2016, WNDS20160929-2. [Google Scholar]
- Local Food-Nutraceuticals Consortium. Understanding local Mediterranean diets: A multidisciplinary pharmacological and ethnobotanical approach. Pharmacol. Res. 2005, 52, 353–366. [Google Scholar] [CrossRef] [PubMed]
- Rauha, J.P.; Remes, S.; Heinonen, M.; Hopia, A.; Kähkönen, M.; Kujala, T.; Pihlaja, K.; Vuorela, H.; Vuorela, P. Antimicrobial effects of Finnish plant extracts containing flavonoids and other phenolic compounds. Int. J. Food Microbiol. 2000, 56, 3–12. [Google Scholar] [CrossRef]
- Piwowarski, J.P.; Kiss, A.K. Contribution of C-glucosidic ellagitannins to Lythrum salicaria L. influence on pro-inflammatory functions of human neutrophils. J. Nat. Med. 2015, 69, 100–110. [Google Scholar] [CrossRef]
- Becker, H.; Scher, J.M.; Speakman, J.B.; Zapp, J. Bioactivity guided isolation of antimicrobial compounds from Lythrum salicaria. Fitoterapia 2005, 76, 580–584. [Google Scholar] [CrossRef]
- Granica, S.; Vahjen, W.; Zentek, J.; Melzig, M.F.; Pawłowska, K.A.; Piwowarski, J.P. Lythrum salicaria Ellagitannins Stimulate IPEC-J2 Cells Monolayer Formation and Inhibit Enteropathogenic Escherichia coli Growth and Adhesion. J. Nat. Prod. 2020, 83, 3614–3622. [Google Scholar] [CrossRef]
- Dadi, T.H.; Vahjen, W.; Zentek, J.; Melzig, M.F.; Granica, S.; Piwowarski, J.P. Lythrum salicaria L. herb and gut microbiota of healthy post-weaning piglets. Focus on prebiotic properties and formation of postbiotic metabolites in ex vivo cultures. J. Ethnopharmacol. 2020, 261, 113073. [Google Scholar] [CrossRef] [PubMed]
- Mahendran, G.; Rahman, L.U. Ethnomedicinal, phytochemical and pharmacological updates on Peppermint (Mentha × piperita L.)-A review. Phytother. Res. 2020, 34, 2088–2139. [Google Scholar] [CrossRef]
- Zhao, H.; Ren, S.; Yang, H.; Tang, S.; Guo, C.; Liu, M.; Tao, Q.; Ming, T.; Xu, H. Peppermint essential oil: Its phytochemistry, biological activity, pharmacological effect and application. Biomed. Pharmacother. 2022, 154, 113559. [Google Scholar] [CrossRef] [PubMed]
- McKemy, D.D.; Neuhausser, W.M.; Julius, D. Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature 2002, 416, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Peier, A.M.; Moqrich, A.; Hergarden, A.C.; Reeve, A.J.; Andersson, D.A.; Story, G.M.; Earley, T.J.; Dragoni, I.; McIntyre, P.; Bevan, S.; et al. A TRP channel that senses cold stimuli and menthol. Cell 2002, 108, 705–715. [Google Scholar] [CrossRef]
- Soković, M.; Glamočlija, J.; Marin, P.D.; Brkić, D.; van Griensven, L.J. Antibacterial effects of the essential oils of commonly consumed medicinal herbs using an in vitro model. Molecules 2010, 15, 7532–7546. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.J.; Baskar, T.B.; Yeo, S.K.; Arasu, M.V.; Al-Dhabi, N.A.; Lim, S.S.; Park, S.U. Composition of volatile compounds and in vitro antimicrobial activity of nine Mentha spp. Springerplus 2016, 5, 1628. [Google Scholar] [CrossRef]
- Ghasemi-Pirbaluti, M.; Motaghi, E.; Bozorgi, H. The effect of menthol on acute experimental colitis in rats. Eur. J. Pharmacol. 2017, 805, 101–107. [Google Scholar] [CrossRef]
- Chang, C.; Miller, J.F. Campylobacter jejuni colonization of mice with limited enteric flora. Infect. Immun. 2006, 74, 5261–5271. [Google Scholar] [CrossRef]
- Heimesaat, M.M.; Mousavi, S.; Bandick, R.; Bereswill, S. Campylobacter jejuni infection induces acute enterocolitis in IL-10−/− mice pretreated with ampicillin plus sulbactam. Eur. J. Microbiol. Immunol. 2022, 12, 73–83. [Google Scholar] [CrossRef]
- Du, K.; Foote, M.S.; Mousavi, S.; Buczkowski, A.; Schmidt, S.; Bereswill, S.; Heimesaat, M.M. Less Pronounced Immunopathological Responses Following Oral Butyrate Treatment of Campylobacter jejuni-Infected Mice. Microorganisms 2022, 10, 1953. [Google Scholar] [CrossRef]
- Erben, U.; Loddenkemper, C.; Doerfel, K.; Spieckermann, S.; Haller, D.; Heimesaat, M.M.; Zeitz, M.; Siegmund, B.; Kühl, A.A. A guide to histomorphological evaluation of intestinal inflammation in mouse models. Int. J. Clin. Exp. Pathol. 2014, 7, 4557–4576. [Google Scholar] [PubMed]
- Heimesaat, M.M.; Giladi, E.; Kühl, A.A.; Bereswill, S.; Gozes, I. The octapetide NAP alleviates intestinal and extra-intestinal anti-inflammatory sequelae of acute experimental colitis. Peptides 2018, 101, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Haag, L.M.; Fischer, A.; Otto, B.; Plickert, R.; Kuhl, A.A.; Gobel, U.B.; Bereswill, S.; Heimesaat, M.M. Campylobacter jejuni induces acute enterocolitis in gnotobiotic IL-10−/− mice via Toll-like-receptor-2 and -4 signaling. PLoS ONE 2012, 7, e40761. [Google Scholar] [CrossRef] [PubMed]
- Heimesaat, M.M.; Bereswill, S.; Fischer, A.; Fuchs, D.; Struck, D.; Niebergall, J.; Jahn, H.K.; Dunay, I.R.; Moter, A.; Gescher, D.M.; et al. Gram-negative bacteria aggravate murine small intestinal Th1-type immunopathology following oral infection with Toxoplasma gondii. J. Immunol. 2006, 177, 8785–8795. [Google Scholar] [CrossRef] [PubMed]
- Bajagai, Y.S.; Petranyi, F.; Yu, S.J.; Lobo, E.; Batacan, R., Jr.; Kayal, A.; Horyanto, D.; Ren, X.; Whitton, M.M.; Stanley, D. Phytogenic supplement containing menthol, carvacrol and carvone ameliorates gut microbiota and production performance of commercial layers. Sci. Rep. 2022, 12, 11033. [Google Scholar] [CrossRef] [PubMed]
- Kovács, J.K.; Felső, P.; Horváth, G.; Schmidt, J.; Dorn, Á.; Ábrahám, H.; Cox, A.; Márk, L.; Emődy, L.; Kovács, T.; et al. Stress Response and Virulence Potential Modulating Effect of Peppermint Essential Oil in Campylobacter jejuni. Biomed. Res. Int. 2019, 2019, 2971741. [Google Scholar] [CrossRef]
- Huber, R.; Ditfurth, A.v.; Amann, F.; Güthlin, C.; Rostock, M.; Trittler, R.; Kümmerer, K.; Merfort, I. Tormentil for Active Ulcerative Colitis: An Open-label, Dose-escalating Study. J. Clin. Gastroenterol. 2007, 41, 834–838. [Google Scholar] [CrossRef]
- Bayat, M.; Kalantar, K.; Amirghofran, Z. Inhibition of interferon-γ production and T-bet expression by menthol treatment of human peripheral blood mononuclear cells. Immunopharmacol. Immunotoxicol. 2019, 41, 267–276. [Google Scholar] [CrossRef]
- Mace, T.A.; King, S.A.; Ameen, Z.; Elnaggar, O.; Young, G.; Riedl, K.M.; Schwartz, S.J.; Clinton, S.K.; Knobloch, T.J.; Weghorst, C.M.; et al. Bioactive compounds or metabolites from black raspberries modulate T lymphocyte proliferation, myeloid cell differentiation and Jak/STAT signaling. Cancer Immunol. Immunother. 2014, 63, 889–900. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Quispe, C.; Castillo, C.M.S.; Caroca, R.; Lazo-Vélez, M.A.; Antonyak, H.; Polishchuk, A.; Lysiuk, R.; Oliinyk, P.; De Masi, L.; et al. Ellagic Acid: A Review on Its Natural Sources, Chemical Stability, and Therapeutic Potential. Oxidative Med. Cell. Longev. 2022, 2022, 3848084. [Google Scholar] [CrossRef] [PubMed]
- Marín, M.; María Giner, R.; Ríos, J.L.; Recio, M.C. Intestinal anti-inflammatory activity of ellagic acid in the acute and chronic dextrane sulfate sodium models of mice colitis. J. Ethnopharmacol. 2013, 150, 925–934. [Google Scholar] [CrossRef] [PubMed]
- Wölfle, U.; Hoffmann, J.; Haarhaus, B.; Rao Mittapalli, V.; Schempp, C.M. Anti-inflammatory and vasoconstrictive properties of Potentilla erecta—A traditional medicinal plant from the northern hemisphere. J. Ethnopharmacol. 2017, 204, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Guo, W.; Jia, Y.; Ye, B.; Liu, S.; Fu, S.; Liu, J.; Hu, G. Menthol Targeting AMPK Alleviates the Inflammatory Response of Bovine Mammary Epithelial Cells and Restores the Synthesis of Milk Fat and Milk Protein. Front. Immunol. 2021, 12, 782989. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Liu, D.; Zhang, X.; Zhou, A.; Su, Y.; He, D.; Fu, S.; Gao, F. Menthol protects dopaminergic neurons against inflammation-mediated damage in lipopolysaccharide (LPS)-Evoked model of Parkinson’s disease. Int. Immunopharmacol. 2020, 85, 106679. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.W.; Echeveste, C.E.; Oshima, K.; Zhang, J.; Yearsley, M.; Yu, J.; Wang, L.S. Anti-colonic Inflammation by Black Raspberries through Regulating Toll-like Receptor-4 Signaling in Interlukin-10 Knockout Mice. J. Cancer Prev. 2020, 25, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Yadav, N.; Chandra, H. Modulation of alveolar macrophage innate response in proinflammatory-, pro-oxidant-, and infection- models by mint extract and chemical constituents: Role of MAPKs. Immunobiology 2018, 223, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Shayya, N.W.; Foote, M.S.; Langfeld, L.Q.; Du, K.; Bandick, R.; Mousavi, S.; Bereswill, S.; Heimesaat, M.M. Human microbiota associated IL-10-/- mice: A valuable enterocolitis model to dissect the interactions of Campylobacter jejuni with host immunity and gut microbiota. Eur. J. Microbiol. Immunol. 2023, 12, 107–122. [Google Scholar] [CrossRef] [PubMed]
- Espín, J.C.; Larrosa, M.; García-Conesa, M.T.; Tomás-Barberán, F. Biological significance of urolithins, the gut microbial ellagic Acid-derived metabolites: The evidence so far. Evid. Based Complement. Altern. Med. 2013, 2013, 270418. [Google Scholar] [CrossRef]
- Ríos, J.-L.; Giner, R.M.; Marín, M.; Recio, M.C. A pharmacological update of ellagic acid. Planta Med. 2018, 84, 1068–1093. [Google Scholar] [CrossRef]
- Rosillo, M.A.; Sánchez-Hidalgo, M.; Cárdeno, A.; Aparicio-Soto, M.; Sánchez-Fidalgo, S.; Villegas, I.; de la Lastra, C.A. Dietary supplementation of an ellagic acid-enriched pomegranate extract attenuates chronic colonic inflammation in rats. Pharmacol. Res. 2012, 66, 235–242. [Google Scholar] [CrossRef] [PubMed]
- De, R.; Sarkar, A.; Ghosh, P.; Ganguly, M.; Karmakar, B.C.; Saha, D.R.; Halder, A.; Chowdhury, A.; Mukhopadhyay, A.K. Antimicrobial activity of ellagic acid against Helicobacter pylori isolates from India and during infections in mice. J. Antimicrob. Chemother. 2018, 73, 1595–1603. [Google Scholar] [CrossRef] [PubMed]
- Avila-Galvez, M.; Gimenez-Bastida, J.; Gonzalez-Sarrias, A.; Espin, J. Tissue deconjugation of urolithin A glucuronide to free urolithin A in systemic inflammation. Food Funct. 2019, 10, 3135–3141. [Google Scholar] [CrossRef] [PubMed]
- Silván, J.M.; Mingo, E.; Hidalgo, M.; de Pascual-Teresa, S.; Carrascosa, A.V.; Martinez-Rodriguez, A.J. Antibacterial activity of a grape seed extract and its fractions against Campylobacter spp. Food Control 2013, 29, 25–31. [Google Scholar] [CrossRef]
- Alejo-Armijo, A.; Glibota, N.; Frías, M.P.; Altarejos, J.; Gálvez, A.; Ortega-Morente, E.; Salido, S. Antimicrobial and antibiofilm activities of procyanidins extracted from laurel wood against a selection of foodborne microorganisms. Int. J. Food Sci. Technol. 2017, 52, 679–686. [Google Scholar] [CrossRef]
- Chen, L.; You, Q.; Hu, L.; Gao, J.; Meng, Q.; Liu, W.; Wu, X.; Xu, Q. The antioxidant procyanidin reduces reactive oxygen species signaling in macrophages and ameliorates experimental colitis in mice. Front. Immunol. 2018, 8, 1910. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Zhang, H.; Li, S.; Xin, D.; Li, S.; Yan, B.; Wang, S.; Liu, C. Procyanidin improves experimental colitis by regulating macrophage polarization. Biomed. Pharmacother. 2023, 165, 115076. [Google Scholar] [CrossRef] [PubMed]
- Pandurangan, A.K.; Mohebali, N.; Esa, N.M.; Looi, C.Y.; Ismail, S.; Saadatdoust, Z. Gallic acid suppresses inflammation in dextran sodium sulfate-induced colitis in mice: Possible mechanisms. Int. Immunopharmacol. 2015, 28, 1034–1043. [Google Scholar] [CrossRef]
- Zhu, L.; Gu, P.; Shen, H. Gallic acid improved inflammation via NF-κB pathway in TNBS-induced ulcerative colitis. Int. Immunopharmacol. 2019, 67, 129–137. [Google Scholar] [CrossRef]
- Gopu, V.; Meena, C.K.; Shetty, P.H. Quercetin influences quorum sensing in food borne bacteria: In-vitro and in-silico evidence. PLoS ONE 2015, 10, e0134684. [Google Scholar] [CrossRef]
- Roy, P.K.; Park, S.-H.; Song, M.G.; Park, S.Y. Antimicrobial Efficacy of Quercetin against Vibrio parahaemolyticus biofilm on food surfaces and downregulation of virulence genes. Polymers 2022, 14, 3847. [Google Scholar] [CrossRef]
- Guazelli, C.F.; Fattori, V.; Colombo, B.B.; Georgetti, S.R.; Vicentini, F.T.; Casagrande, R.; Baracat, M.M.; Verri Jr, W.A. Quercetin-loaded microcapsules ameliorate experimental colitis in mice by anti-inflammatory and antioxidant mechanisms. J. Nat. Prod. 2013, 76, 200–208. [Google Scholar] [CrossRef]
- Ju, S.; Ge, Y.; Li, P.; Tian, X.; Wang, H.; Zheng, X.; Ju, S. Dietary quercetin ameliorates experimental colitis in mouse by remodeling the function of colonic macrophages via a heme oxygenase-1-dependent pathway. Cell Cycle 2018, 17, 53–63. [Google Scholar] [CrossRef]
- Araújo, A.R.; Araújo, A.C.; Reis, R.L.; Pires, R.A. Vescalagin and castalagin present bactericidal activity toward methicillin-resistant bacteria. ACS Biomater. Sci. Eng. 2021, 7, 1022–1030. [Google Scholar] [CrossRef]
- Funatogawa, K.; Hayashi, S.; Shimomura, H.; Yoshida, T.; Hatano, T.; Ito, H.; Hirai, Y. Antibacterial activity of hydrolyzable tannins derived from medicinal plants against Helicobacter pylori. Microbiol. Immunol. 2004, 48, 251–261. [Google Scholar] [CrossRef]
- Fumagalli, M.; Sangiovanni, E.; Vrhovsek, U.; Piazza, S.; Colombo, E.; Gasperotti, M.; Mattivi, F.; De Fabiani, E.; Dell’Agli, M. Strawberry tannins inhibit IL-8 secretion in a cell model of gastric inflammation. Pharmacol. Res. 2016, 111, 703–712. [Google Scholar] [CrossRef] [PubMed]








| Treatment | Daily Dose (mg/kg) | Drinking Solution (mg/L) | Minimal Inhibitory Concentration (mg/L) |
|---|---|---|---|
| Tormentil | 12 | 60 | >3840 |
| Raspberry leaves | 40 | 200 | >12,800 |
| Loosestrife | 40 | 200 | >12,800 |
| Menthol | 100 | 500 | 16,000 |
| Combination | 192 | 960 | >15,360 |
| Treatment | Results |
|---|---|
| TORMENTIL | -  T lymphocytes (colon) vs. PLC T lymphocytes (colon) vs. PLC- basal IFN-γ (MLN) |
| RASPBERRY LEAVES | - basal IL-6 (MLN) |
| LOOSESTRIFE | - no significant effect |
| MENTHOL | - improved clinical outcome vs. PLC - T cells (colon) vs. PLC |
| COMBINATION | - diarrheal symptoms vs. PLC - T lymphocytes (colon) vs. PLC- MLN: basal TNF-α basal IFN-γ basal IL-6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bandick, R.; Busmann, L.V.; Mousavi, S.; Shayya, N.W.; Piwowarski, J.P.; Granica, S.; Melzig, M.F.; Bereswill, S.; Heimesaat, M.M. Therapeutic Effects of Oral Application of Menthol and Extracts from Tormentil (Potentilla erecta), Raspberry Leaves (Rubus idaeus), and Loosestrife (Lythrum salicaria) during Acute Murine Campylobacteriosis. Pharmaceutics 2023, 15, 2410. https://doi.org/10.3390/pharmaceutics15102410
Bandick R, Busmann LV, Mousavi S, Shayya NW, Piwowarski JP, Granica S, Melzig MF, Bereswill S, Heimesaat MM. Therapeutic Effects of Oral Application of Menthol and Extracts from Tormentil (Potentilla erecta), Raspberry Leaves (Rubus idaeus), and Loosestrife (Lythrum salicaria) during Acute Murine Campylobacteriosis. Pharmaceutics. 2023; 15(10):2410. https://doi.org/10.3390/pharmaceutics15102410
Chicago/Turabian StyleBandick, Rasmus, Lia V. Busmann, Soraya Mousavi, Nizar W. Shayya, Jakub P. Piwowarski, Sebastian Granica, Matthias F. Melzig, Stefan Bereswill, and Markus M. Heimesaat. 2023. "Therapeutic Effects of Oral Application of Menthol and Extracts from Tormentil (Potentilla erecta), Raspberry Leaves (Rubus idaeus), and Loosestrife (Lythrum salicaria) during Acute Murine Campylobacteriosis" Pharmaceutics 15, no. 10: 2410. https://doi.org/10.3390/pharmaceutics15102410
APA StyleBandick, R., Busmann, L. V., Mousavi, S., Shayya, N. W., Piwowarski, J. P., Granica, S., Melzig, M. F., Bereswill, S., & Heimesaat, M. M. (2023). Therapeutic Effects of Oral Application of Menthol and Extracts from Tormentil (Potentilla erecta), Raspberry Leaves (Rubus idaeus), and Loosestrife (Lythrum salicaria) during Acute Murine Campylobacteriosis. Pharmaceutics, 15(10), 2410. https://doi.org/10.3390/pharmaceutics15102410