
Drug Repurposing Based on Protozoan Proteome: In Vitro Evaluation of In Silico Screened Compounds against Toxoplasma gondii



 and
and 
Abstract
1. Introduction
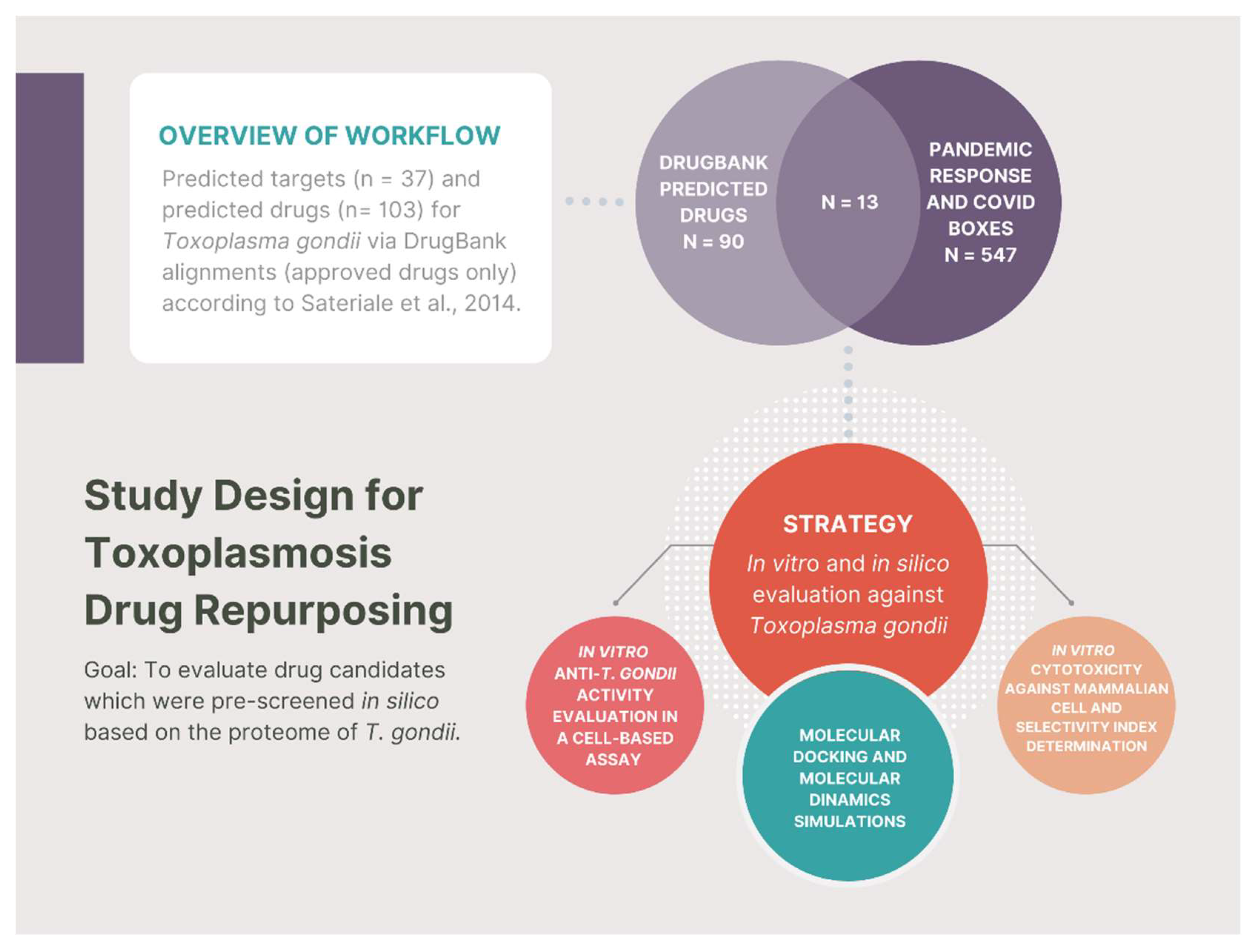
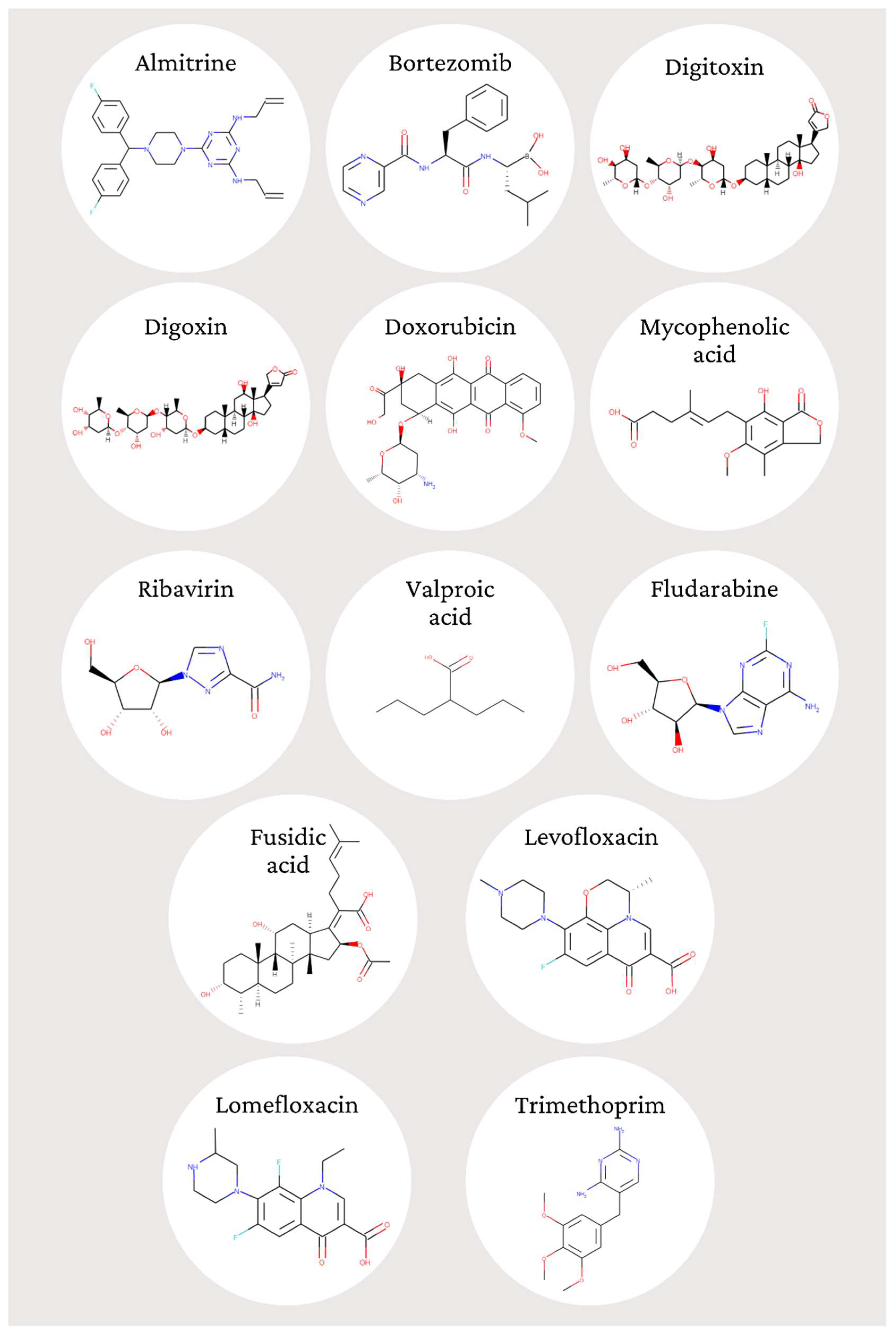
- To demonstrate the in vitro anti-T. gondii activity of the 13 pre-screened compounds: almitrine, bortezomib, digitoxin, digoxin, doxorubicin, fludarabine, fusidic acid, levofloxacin, lomefloxacin, mycophenolic acid, ribavirin, trimethoprim, and valproic acid;
- To demonstrate the in vitro cytotoxic of almitrine, bortezomib and fludarabine against human foreskin fibroblasts;
- To investigate the mechanism of action of the 13 compounds using Molecular Docking through the binding affinity of the compound and the predicted molecular target;
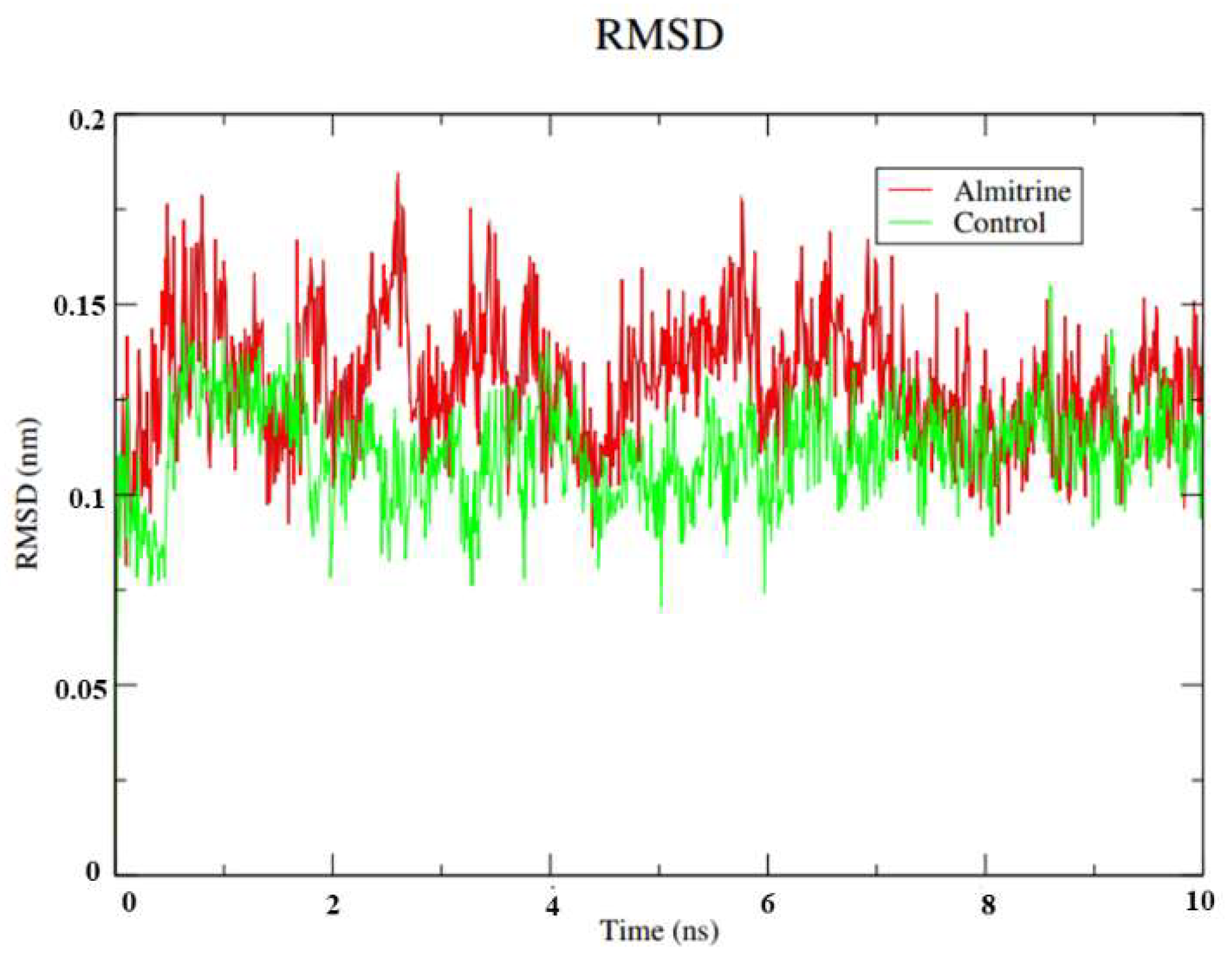
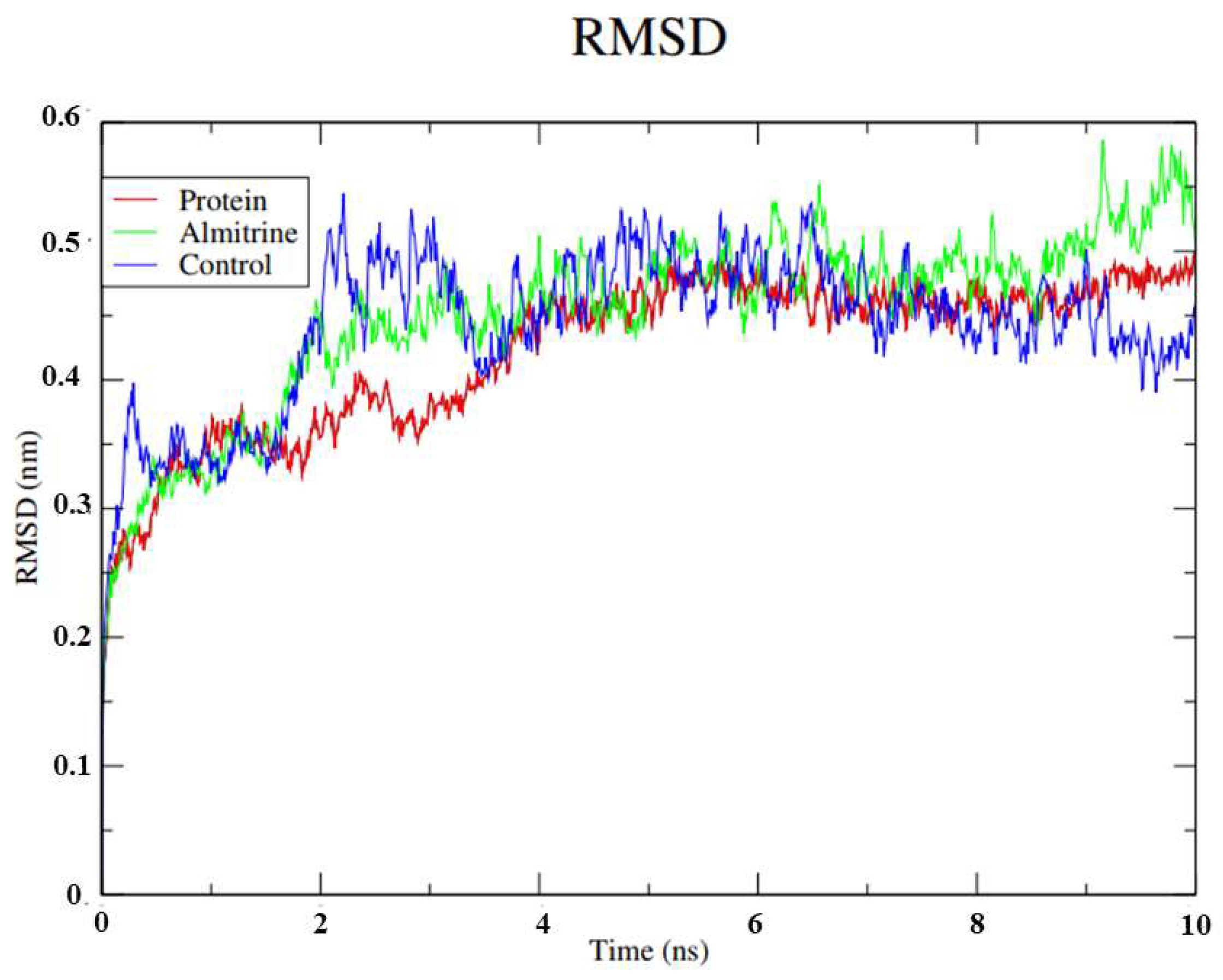
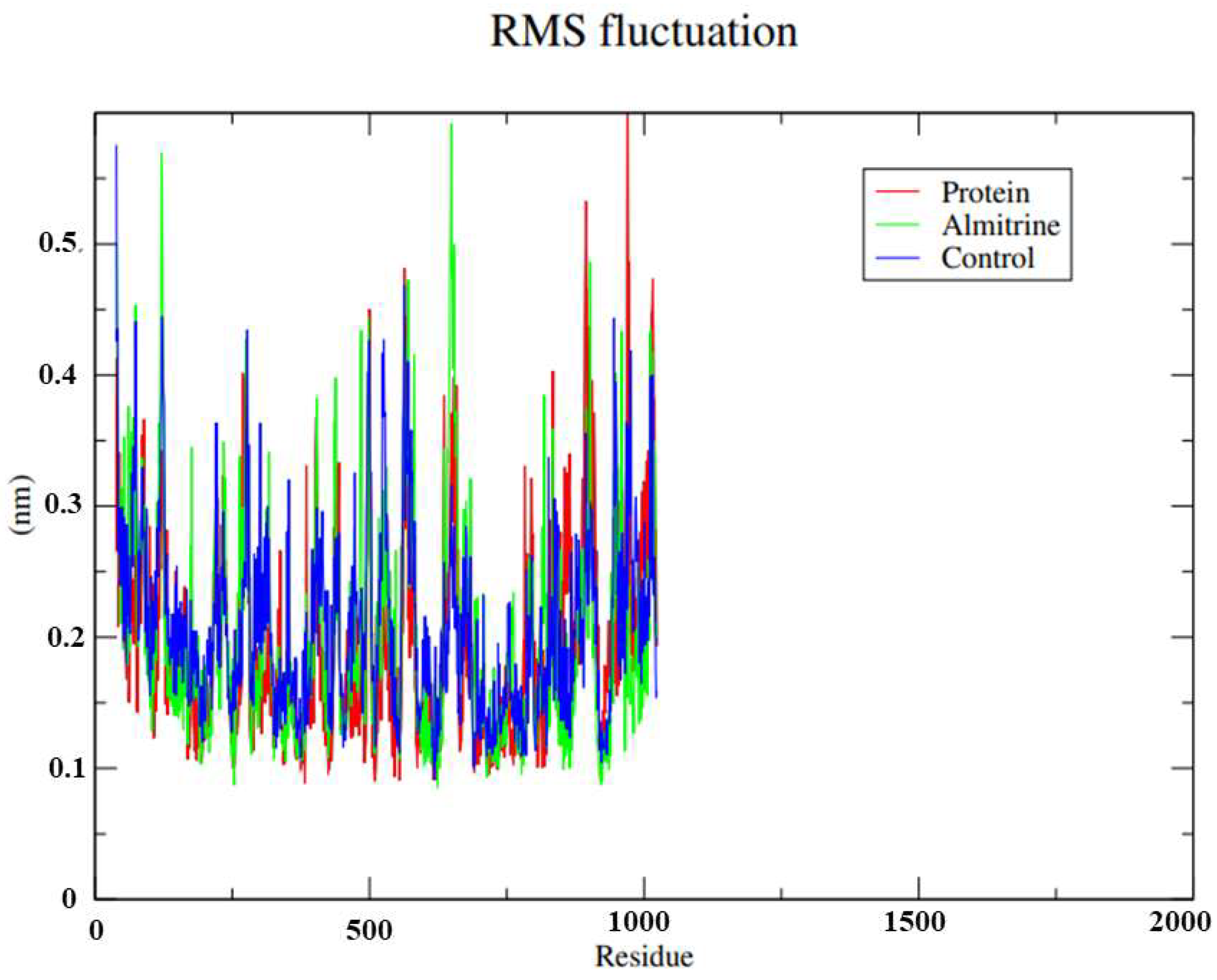
- To carry out Molecular Dynamics simulations of almitrine to assess the flexibility of the transporting ATPase alpha 1 and the stability of the enzyme interactions in the presence of factors such as solvent, ions, pressure and temperature.
2. Materials and Methods
2.1. Drugs and Chemicals
2.2. Cell Culture and Parasite Propagation
2.3. β-Galactosidase-Based Growth Inhibition Assays
2.4. Cytotoxicity in Mammalian Cells
2.5. Molecular Docking
Docking Consensus
2.6. Alignment of Protein Sequences
2.7. Modeling by Homology
2.8. Molecular Dynamics Simulations
3. Results
3.1. In Vitro Anti-T. gondii Activity and Cytotoxicity against HFF
3.2. In Silico Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Kochanowsky, J.A.; Koshy, A.A. Toxoplasma Gondii. Curr. Biol. 2018, 28, R770–R771. [Google Scholar] [CrossRef] [PubMed]
- Radke, J.B.; Burrows, J.N.; Goldberg, D.E.; Sibley, L.D. Evaluation of Current and Emerging Antimalarial Medicines for Inhibition of Toxoplasma gondii Growth in Vitro. ACS Infect. Dis. 2018, 4, 1264–1274. [Google Scholar] [CrossRef] [PubMed]
- CDC. Toxoplasmosis. Available online: https://www.cdc.gov/parasites/toxoplasmosis/ (accessed on 16 March 2022).
- Garcia Bahia-Oliveira, L.M.; Jones, J.L.; Azevedo-Silva, J.; Alves, C.C.F.; Oréfice, F.; Addiss, D.G. Highly Endemic, Waterborne Toxoplasmosis in North Rio de Janeiro State, Brazil. Emerg. Infect. Dis. 2003, 9, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Aspinall, T.V.; Joynson, D.H.M.; Guy, E.; Hyde, J.E.; Sims, P.F.G. The Molecular Basis of Sulfonamide Resistance in Toxoplasma gondii and Implications for the Clinical Management of Toxoplasmosis. J. Infect. Dis. 2002, 185, 1637–1643. [Google Scholar] [CrossRef]
- Dittmar, A.J.; Drozda, A.A.; Blader, I.J. Drug Repurposing Screening Identifies Novel Compounds That Effectively Inhibit Toxoplasma gondii Growth. mSphere 2016, 1, e00042-15. [Google Scholar] [CrossRef]
- Blader, I.J.; Saeij, J.P. Communication between Toxoplasma gondii and Its Host: Impact on Parasite Growth, Development, Immune Evasion, and Virulence. Apmis 2009, 117, 458–476. [Google Scholar] [CrossRef]
- Alday, P.H.; Doggett, J.S. Drugs in development for toxoplasmosis: Advances, challenges, and current status. Drug Des. Dev. Ther. 2017, 25, 273–293. [Google Scholar] [CrossRef]
- Buckle, D.R.; Erhardt, P.W.; Ganellin, C.R.; Kobayashi, T.; Perun, T.J.; Proudfoot, J.; Senn-Bilfinger, J. Glossary of Terms Used in Medicinal Chemistry Part II (IUPAC Recommendations 2013). Annu. Rep. Med. Chem. 2013, 48, 387–418. [Google Scholar] [CrossRef]
- MMV. The Pandemic Response Box|Medicines for Malaria Venture. Available online: https://www.mmv.org/mmv-open/pandemic-response-box (accessed on 16 March 2022).
- Duffy, S.; Sykes, M.L.; Jones, A.J.; Shelper, T.B.; Simpson, M.; Lang, R.; Poulsen, S.A.; Sleebs, B.E.; Avery, V.M. Screening the Medicines for Malaria Venture Pathogen Box across Multiple Pathogens Reclassifies Starting Points for Open-Source Drug Discovery. Antimicrob. Agents Chemother. 2017, 61, e00379-17. [Google Scholar] [CrossRef]
- Spalenka, J.; Escotte-Binet, S.; Bakiri, A.; Hubert, J.; Renault, J.H.; Velard, F.; Duchateau, S.; Aubert, D.; Huguenin, A.; Villena, I. Discovery of New Inhibitors of Toxoplasma gondii via the Pathogen Box. Antimicrob. Agents Chemother. 2018, 62, e01640-17. [Google Scholar] [CrossRef]
- Sateriale, A.; Bessoff, K.; Sarkar, I.N.; Huston, C.D. Drug Repurposing: Mining Protozoan Proteomes for Targets of Known Bioactive Compounds. J. Am. Med. Inform. Assoc. 2014, 21, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.D.; Teixeira, C.; Gomes, P.; Borges, M. Promising Drug Targets and Compounds with Anti-Toxoplasma gondii Activity. Microorganisms 2021, 15, 1960. [Google Scholar] [CrossRef] [PubMed]
- Castro, A.S.; Alves, C.M.O.S.; Angeloni, M.B.; Gomes, A.O.; Barbosa, B.F.; Franco, P.S.; Silva, D.A.O.; Martins-Filho, O.A.; Mineo, J.R.; Mineo, T.W.P.; et al. Trophoblast Cells Are Able to Regulate Monocyte Activity to Control Toxoplasma gondii Infection. Placenta 2013, 34, 240–247. [Google Scholar] [CrossRef]
- Wang, Q.; Sibley, L.D. Assays for Monitoring Toxoplasma gondii Infectivity in the Laboratory Mouse. Methods Mol. Biol. 2020, 2071, 99–116. [Google Scholar] [CrossRef] [PubMed]
- Chin, F.T.; Xing, W.Z.; Bogyo, M.; Carruthers, V.B. Cysteine Protease Inhibitors Block Toxoplasma gondii Microneme Secretion and Cell Invasion. Antimicrob. Agents Chemother. 2007, 51, 679–688. [Google Scholar] [CrossRef]
- Reimão, J.Q.; Mesquita, J.T.; Ferreira, D.D.; Tempone, A.G. Investigation of Calcium Channel Blockers as Antiprotozoal Agents and Their Interference in the Metabolism of Leishmania (L.) Infantum. Evid.-Based Complementary Altern. Med. 2016, 2016, 1523691. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, F.C.; Koetzle, T.F.; Williams, G.J.B.; Meyer, E.F., Jr.; Brice, M.D.; Rodgers, J.R.; Kennard, O.; Shimanouchi, T.; Tasumi, M. NoA Computer-based Archival File for Macromolecular Structures Title. Eur. J. Biochem. 1977, 112, 535–542. [Google Scholar]
- Yusuf, D.; Davis, A.M.; Kleywegt, G.J.; Schmitt, S. An Alternative Method for the Evaluation of Docking Performance: RSR vs. RMSD. J. Chem. Inf. Model. 2008, 48, 1411–1422. [Google Scholar] [CrossRef]
- Thomsen, R.; Christensen, M.H. MolDock: A New Technique for High-Accuracy Molecular Docking. J. Med. Chem. 2006, 49, 3315–3321. [Google Scholar] [CrossRef]
- Da Silva Calixto, P.; de Almeida, R.N.; Salvadori, M.G.S.S.; Dos Santos Maia, M.; Filho, J.M.B.; Scotti, M.T.; Scotti, L. In Silico Study Examining New Phenylpropanoids Targets with Antidepressant Activity. Curr. Drug Targets 2021, 22, 539–554. [Google Scholar] [CrossRef]
- De Azevedo, W., Jr. MolDock Applied to Structure-Based Virtual Screening. Curr. Drug Targets 2010, 11, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Goodsell, D.S.; Huey, R.; Lindstrom, W.; Hart, W.E.; Kurowski, S.; Halliday, S.; Belew, R.; Olson, A.J. AutoDock Tolls 4.2. Available online: https://onlinelibrary.wiley.com/doi/full/10.1002/jcc.21256 (accessed on 14 March 2022).
- Solis-Vasquez, L.; Santos-Martins, D.; Koch, A.; Forli, S. Evaluating the Energy Efficiency of Opencl-Accelerated Autodock Molecular Docking. In Proceedings of the 2020 28th Euromicro International Conference on Parallel, Distributed and Network-Based Processing (PDP), IEEE, Västerås, Sweden, 11–13 March 2020; pp. 162–166. [Google Scholar]
- Trott, O.; Olson, A.J. AutoDock Vina 1.1.1. Available online: https://pubmed.ncbi.nlm.nih.gov/19499576/ (accessed on 14 March 2022).
- Trott, O.; Olson, A.J. Software News and Update AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function. Effic. Optim. Multithreading 2009, 31, 455–461. [Google Scholar]
- Zaware, N.; Sharma, H.; Yang, J.; Devambatla, R.K.V.; Queener, S.F.; Anderson, K.S.; Gangjee, A. Discovery of Potent and Selective Inhibitors of Toxoplasma gondii Thymidylate Synthase for Opportunistic Infections. ACS Med. Chem. Lett. 2013, 4, 1148–1151. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, T.M.; Cassera, M.B.; Ho, M.C.; Zhan, C.; Merino, E.F.; Evans, G.B.; Tyler, P.C.; Almo, S.C.; Schramm, V.L.; Kim, K. Inhibition and Structure of Toxoplasma gondii Purine Nucleoside Phosphorylase. Eukaryot. Cell 2014, 13, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Muench, S.P.; Prigge, S.T.; McLeod, R.; Rafferty, J.B.; Kirisits, M.J.; Roberts, C.W.; Mui, E.J.; Rice, D.W. Studies of Toxoplasma gondii and Plasmodium Falciparum Enoyl Acyl Carrier Protein Reductase and Implications for the Development of Antiparasitic Agents. Acta Crystallogr. Sect. D Biol. Crystallogr. 2007, 63, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Ojo, K.K.; Vidadala, R.; Huang, W.; Geiger, J.A.; Scheele, S.; Choi, R.; Reid, M.C.; Keyloun, K.R.; Rivas, K.; et al. Potent and Selective Inhibitors of CDPK1 from T. Gondii and C. Parvum Based on a 5-Aminopyrazole-4-Carboxamide Scaffold. ACS Med. Chem. Lett. 2014, 5, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Liang, L.J.; Zhang, B.B.; Lou, Z.L.; Zhang, H.F.; Shen, X.; Wu, Y.Q.; Wang, Z.M.; Tang, R.X.; Fu, L.L.; et al. Prevalence and Genotyping of Toxoplasma gondii in Naturally-Infected Synanthropic Rats (Rattus Norvegicus) and Mice (Mus Musculus) in Eastern China. Parasites Vectors 2014, 7, 591. [Google Scholar] [CrossRef]
- Xue, Q.; Liu, X.; Russell, P.; Li, J.; Pan, W.; Fu, J.; Zhang, A. Evaluation of the Binding Performance of Flavonoids to Estrogen Receptor Alpha by Autodock, Autodock Vina and Surflex-Dock. Ecotoxicol. Environ. Saf. 2022, 233, 113323. [Google Scholar] [CrossRef]
- Dos Santos Maia, M.; de Sousa, N.F.; Rodrigues, G.C.S.; Monteiro, A.F.M.; Scotti, M.T.; Scotti, L. Lignans and Neolignans Anti-Tuberculosis Identified by QSAR and Molecular Modeling. Comb. Chem. High Throughput Screen. 2020, 23, 504–516. [Google Scholar] [CrossRef]
- Mito, T.; Kuwahara, S.; Delamere, N.A. The Influence of Thapsigargin on Na,k-ATPase Activity in Cultured Nonpigmented Ciliary Epithelial Cells. Curr. Eye Res. 1995, 14, 651–657. [Google Scholar] [CrossRef]
- RCSB. Protein Data Bank. Available online: https://www.rcsb.org/.2022 (accessed on 16 March 2022). [CrossRef]
- Engel. NCBI. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 16 March 2022).
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; De Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology Modelling of Protein Structures and Complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A Program to Check the Stereochemical Quality of Protein Structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef]
- Berendsen, H.J.C.; van der Spoel, D.; van Drunen, R. GROMACS: A message-passing parallel molecular dynamics implementation. Comput. Phys. Commun. 1995, 91, 43–56. [Google Scholar] [CrossRef]
- Bondi, A. Van der Waals Volumes and Radii. J. Phys. Chem. 1964, 68, 441–451. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Caplan, M.; Goutay, J.; Bignon, A.; Jaillette, E.; Favory, R.; Mathieu, D.; Parmentier-Decrucq, E.; Poissy, J.; Duburcq, T. Lille Intensive Care COVID-19 Group. Almitrine Infusion in Severe Acute Respiratory Syndrome Coronavirus 2-Induced Acute Respiratory Distress Syndrome: A Single-Center Observational Study. Crit. Care Med. 2021, 1, e191–e198. [Google Scholar] [CrossRef]
- Basco, L.K.; Le Bras, J. In Vitro Activity of Mitochondrial ATP Synthetase Inhibitors Against Plasmodium Falciparum. J. Eukaryot. Microbiol. 1994, 41, 179–183. [Google Scholar] [CrossRef]
- MacLeod, C.N.; Thomas, R.W.; Bartley, E.A.; Parkhurst, G.W.; Bachand, R.T. Effects and handling of almitrine bismesylate in healthy subjects. Eur. J. Respir. Dis. Suppl. 1983, 126, 275–289. [Google Scholar]
- Jung, L.; Holle, L.; Dalton, W.S. Discovery, Development, and clinical applications of bortezomib. Oncology 2004, 18 (14 Suppl. 11), 4–13. [Google Scholar]
- Reynolds, J.M.; El Bissati, K.; Brandenburg, J.; Günzl, A.; Mamoun, C.B. Antimalarial Activity of the Anticancer and Proteasome Inhibitor Bortezomib and Its Analog ZL3B. BMC Clin. Pharmacol. 2007, 7, 13. [Google Scholar] [CrossRef] [PubMed]
- LiverTox: Clinical and Research Information on Drug-Induced Liver Injury. Fludarabine. 1 February 2018; National Institute of Diabetes and Digestive and Kidney Diseases: Bethesda, MD, USA, 2012.
- Octavia, Y.; Tocchetti, C.G.; Gabrielson, K.L.; Janssens, S.; Crijns, H.J.; Moens, A.L. Doxorubicin-induced cardiomyopathy: From molecular mechanisms to therapeutic strategies. J. Mol. Cell. Cardiol. 2012, 52, 1213–1225. [Google Scholar] [CrossRef] [PubMed]
- Andrews, K.T.; Fisher, G.; Skinner-Adams, T.S. Drug repurposing and human parasitic protozoan diseases. Int. J. Parasitol. Drugs Drug Resist. 2014, 4, 95–111. [Google Scholar] [CrossRef] [PubMed]
- Rajapakse, S.; Shivanthan, M.C.; Samaranayake, N.; Rodrigo, C.; Fernando, S.D. Antibiotics for human toxoplasmosis: A systematic review of randomized trials. Pathog. Glob. Health 2013, 107, 162–169. [Google Scholar] [CrossRef]
- Castro-Elizalde, K.N.; Hernández-Contreras, P.; Ramírez-Flores, C.J.; González-Pozos, S.; Gómez de León, C.T.; Mondragón-Castelán, M.; Mondragón-Flores, R. Mycophenolic acid induces differentiation of Toxoplasma gondii RH strain tachyzoites into bradyzoites and formation of cyst-like structure in vitro. Parasitol. Res. 2018, 117, 547–563. [Google Scholar] [CrossRef] [PubMed]
- Payne, A.J.; Neal, L.M.; Knoll, L.J. Fusidic acid is an effective treatment against Toxoplasma gondii and Listeria monocytogenes in vitro, but not in mice. Parasitol. Res. 2013, 112, 3859–3863. [Google Scholar] [CrossRef]
- Kemnic, T.R.; Coleman, M. Trimethoprim Sulfamethoxazole. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Al-Wabli, R.I. Lomefloxacin. In Profiles of Drug Substances, Excipients and Related Methodology; Academic Press: Cambridge, MA, USA, 2017; Volume 42, pp. 193–240. [Google Scholar] [CrossRef]
- Tunitskaya, V.L.; Khomutov, A.R.; Kochetkov, S.N.; Kotovskaya, S.K.; Charushin, V.N. Inhibition of DNA gyrase by levofloxacin and related fluorine-containing heterocyclic compounds. Acta Nat. 2011, 3, 94–99. [Google Scholar] [CrossRef]
- Elbaz, H.A.; Stueckle, T.A.; Tse, W.; Rojanasakul, Y.; Dinu, C.Z. Digitoxin and its analogs as novel cancer therapeutics. Exp. Hematol. Oncol. 2012, 1, 4. [Google Scholar] [CrossRef]
- Ren, Y.; Ribas, H.T.; Heath, K.; Wu, S.; Ren, J.; Shriwas, P.; Chen, X.; Johnson, M.E.; Cheng, X.; Burdette, J.E.; et al. Na+/K+-ATPase-Targeted Cytotoxicity of (+)-Digoxin and Several Semisynthetic Derivatives. J. Nat. Prod. 2020, 83, 638–648. [Google Scholar] [CrossRef]
- Hofmann, W.P.; Herrmann, E.; Sarrazin, C.; Zeuzem, S. Ribavirin mode of action in chronic hepatitis C: From clinical use back to molecular mechanisms. Liver Int. 2008, 28, 1332–1343. [Google Scholar] [CrossRef]
- Enshaeieh, M.; Saadatnia, G.; Babaie, J.; Golkar, M.; Choopani, S.; Sayyah, M. Valproic Acid Inhibits Chronic Toxoplasma Infection and Associated Brain Inflammation in Mice. Antimicrob. Agents Chemother. 2021, 65, e0100321. [Google Scholar] [CrossRef] [PubMed]
- Lovell, S.C.; Davis, I.W.; Arendall, W.B.; De Bakker, P.I.W.; Word, J.M.; Prisant, M.G.; Richardson, J.S.; Richardson, D.C. Structure Validation by Cα Geometry: φ,ψ and Cβ Deviation. Proteins Struct. Funct. Genet. 2003, 50, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Hilge, M.; Siegal, G.; Vuister, G.W.; Güntert, P.; Gloor, S.M.; Abrahams, J.P. ATP-Induced Conformational Changes of the Nucleotide-Binding Domain of Na, K-ATPase. Nat. Struct. Mol. Biol. 2003, 10, 468–474. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MMV Code a | Compound (Trivial Name) | Molecular Formula b | Mol wt b | aLogP b | Rule of Five b |
|---|---|---|---|---|---|
| MMV1804175 | Almitrine | C26H29F2N7 | 477.5 | 6.09 | 3 |
| MMV009415 | Bortezomib | C19H25BN4O4 | 384.2 | 2.14 | 4 |
| MMV002436 | Digitoxin | C41H64O13 | 764.9 | 3.11 | 2 |
| MMV002832 | Digoxin | C41H64O14 | 780.9 | 2 | 1 |
| MMV004066 | Doxorubicin | C27H29NO11 | 543.5 | −0.05 | 1 |
| MMV003219 | Mycophenolic acid | C17H20O6 | 320.3 | 3.16 | 4 |
| MMV001439 | Ribavirin | C8H12N4O5 | 244.2 | −2.75 | 4 |
| MMV003305 | Valproic acid | C8H16O2 | 144.2 | 2.75 | 4 |
| MMV637413 | Fludarabine | C10H12FN5O4 | 285.2 | −1.32 | 4 |
| MMV1578575 | Fusidic acid | C31H48O6 | 516.7 | 5.1 | 2 |
| MMV687798 | Levofloxacin | C18H20FN3O4 | 361.4 | −1.38 | 4 |
| MMV002350 | Lomefloxacin | C17H19F2N3O3 | 387.8 | −0.83 | 4 |
| MMV000028 | Trimethoprim | C14H18N4O3 | 290.3 | 1.55 | 4 |
| Compound | EC50 (µM) a | CC50 (µM) b | SI c |
|---|---|---|---|
| Almitrine | 0.424 | >20 | >47 |
| Bortezomib | 0.223 | 0.079 | 0.35 |
| Digitoxin | 5.66 | n.d. | n.d. |
| Digoxin | 42.59 | n.d. | n.d. |
| Doxorubicin | 2.39 | n.d. | n.d. |
| Mycophenolic acid | 8.06 | n.d. | n.d. |
| Ribavirin | 83.31 | n.d. | n.d. |
| Valproic acid | 99.61 | n.d. | n.d. |
| Fludarabine | 0.75 | 2.140 | 2.85 |
| Fusidic acid | 16.70 | n.d. | n.d. |
| Levofloxacin | 70.58 | n.d. | n.d. |
| Lomefloxacin | 7.32 | n.d. | n.d. |
| Trimethoprim | 7.36 | n.d. | n.d. |
| Pyrimethamine | 0.121 | n.d. | n.d. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cajazeiro, D.C.; Toledo, P.P.M.; de Sousa, N.F.; Scotti, M.T.; Reimão, J.Q. Drug Repurposing Based on Protozoan Proteome: In Vitro Evaluation of In Silico Screened Compounds against Toxoplasma gondii. Pharmaceutics 2022, 14, 1634. https://doi.org/10.3390/pharmaceutics14081634
Cajazeiro DC, Toledo PPM, de Sousa NF, Scotti MT, Reimão JQ. Drug Repurposing Based on Protozoan Proteome: In Vitro Evaluation of In Silico Screened Compounds against Toxoplasma gondii. Pharmaceutics. 2022; 14(8):1634. https://doi.org/10.3390/pharmaceutics14081634
Chicago/Turabian StyleCajazeiro, Débora Chaves, Paula Pereira Marques Toledo, Natália Ferreira de Sousa, Marcus Tullius Scotti, and Juliana Quero Reimão. 2022. "Drug Repurposing Based on Protozoan Proteome: In Vitro Evaluation of In Silico Screened Compounds against Toxoplasma gondii" Pharmaceutics 14, no. 8: 1634. https://doi.org/10.3390/pharmaceutics14081634
APA StyleCajazeiro, D. C., Toledo, P. P. M., de Sousa, N. F., Scotti, M. T., & Reimão, J. Q. (2022). Drug Repurposing Based on Protozoan Proteome: In Vitro Evaluation of In Silico Screened Compounds against Toxoplasma gondii. Pharmaceutics, 14(8), 1634. https://doi.org/10.3390/pharmaceutics14081634