
Prospective Application of Partially Digested Autologous Chondrocyte for Meniscus Tissue Engineering

 ,
,
Abstract
1. Introduction
2. Materials and Methods
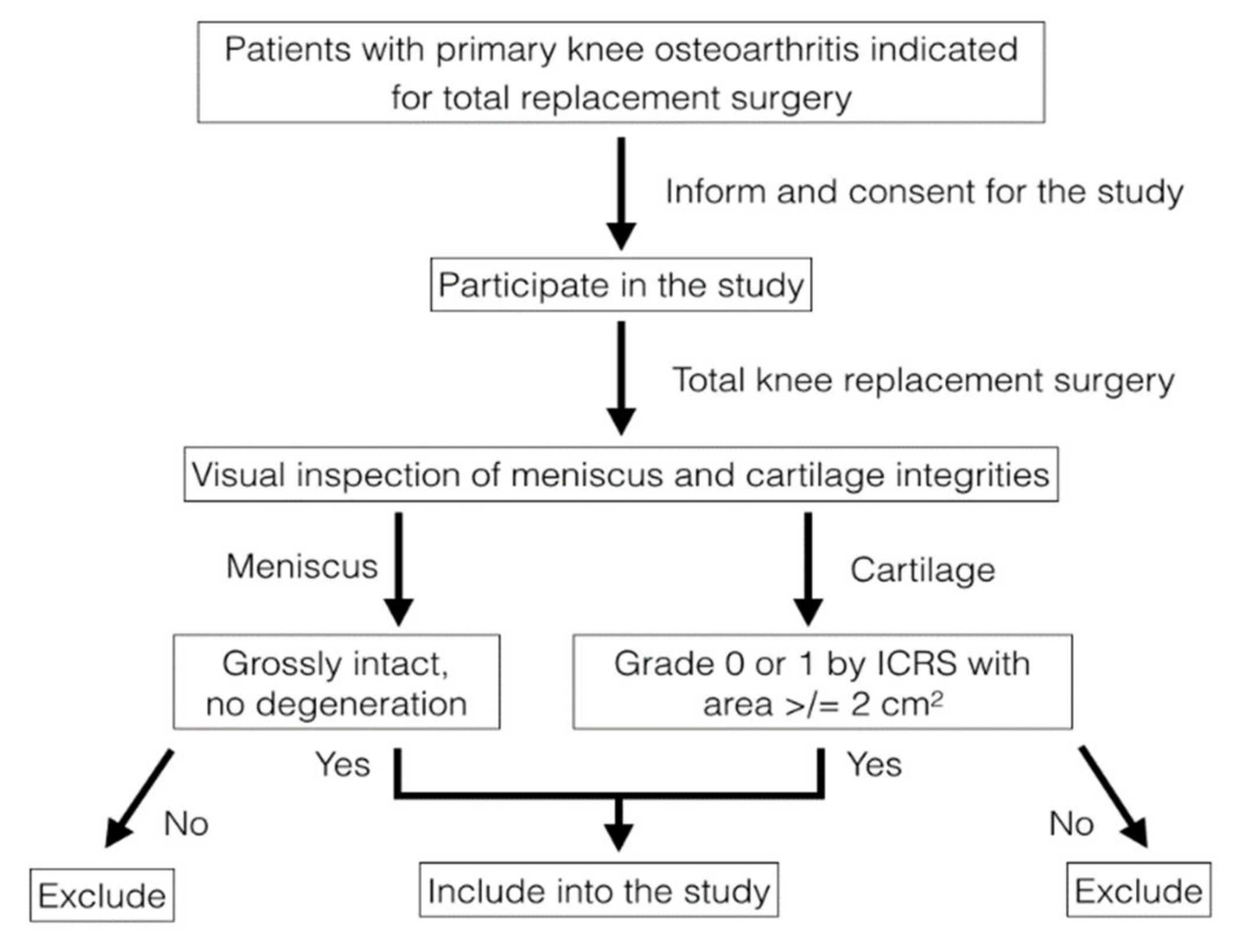
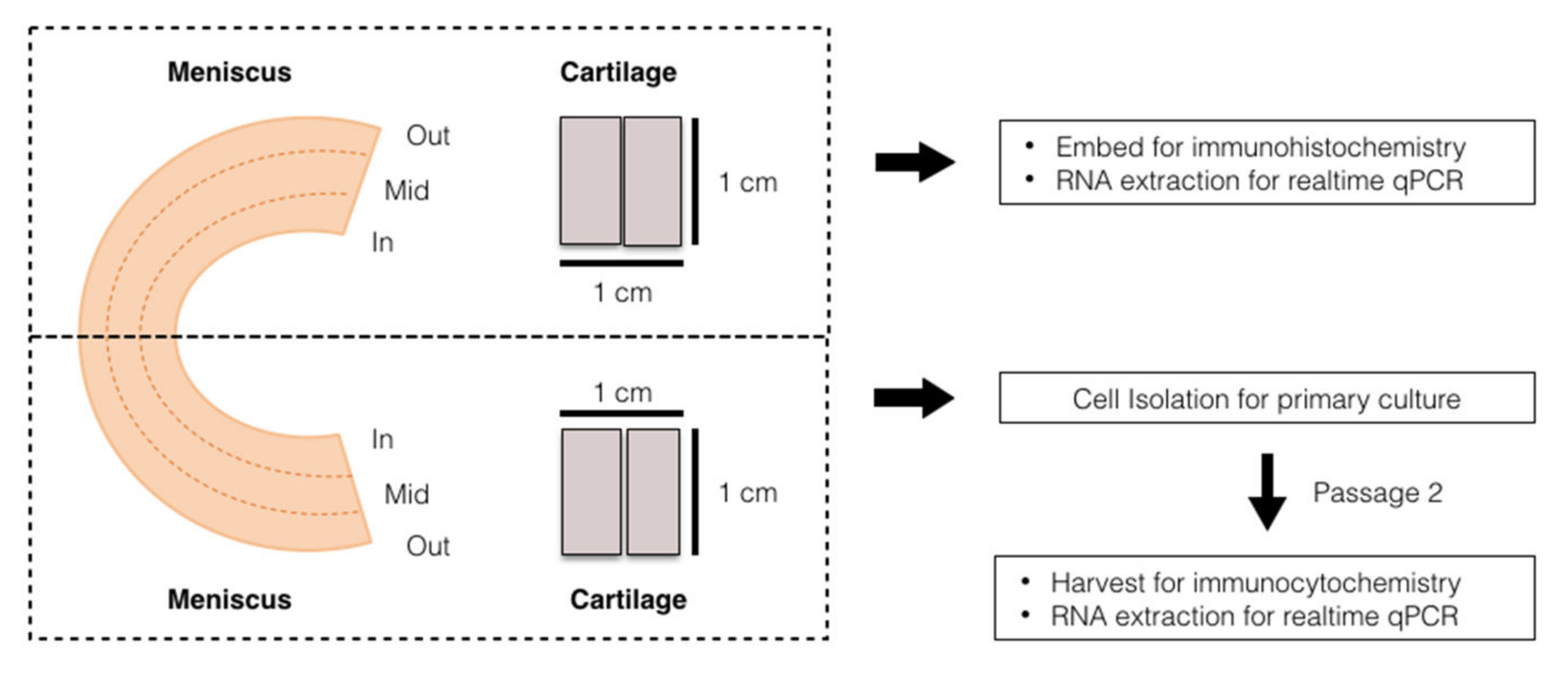
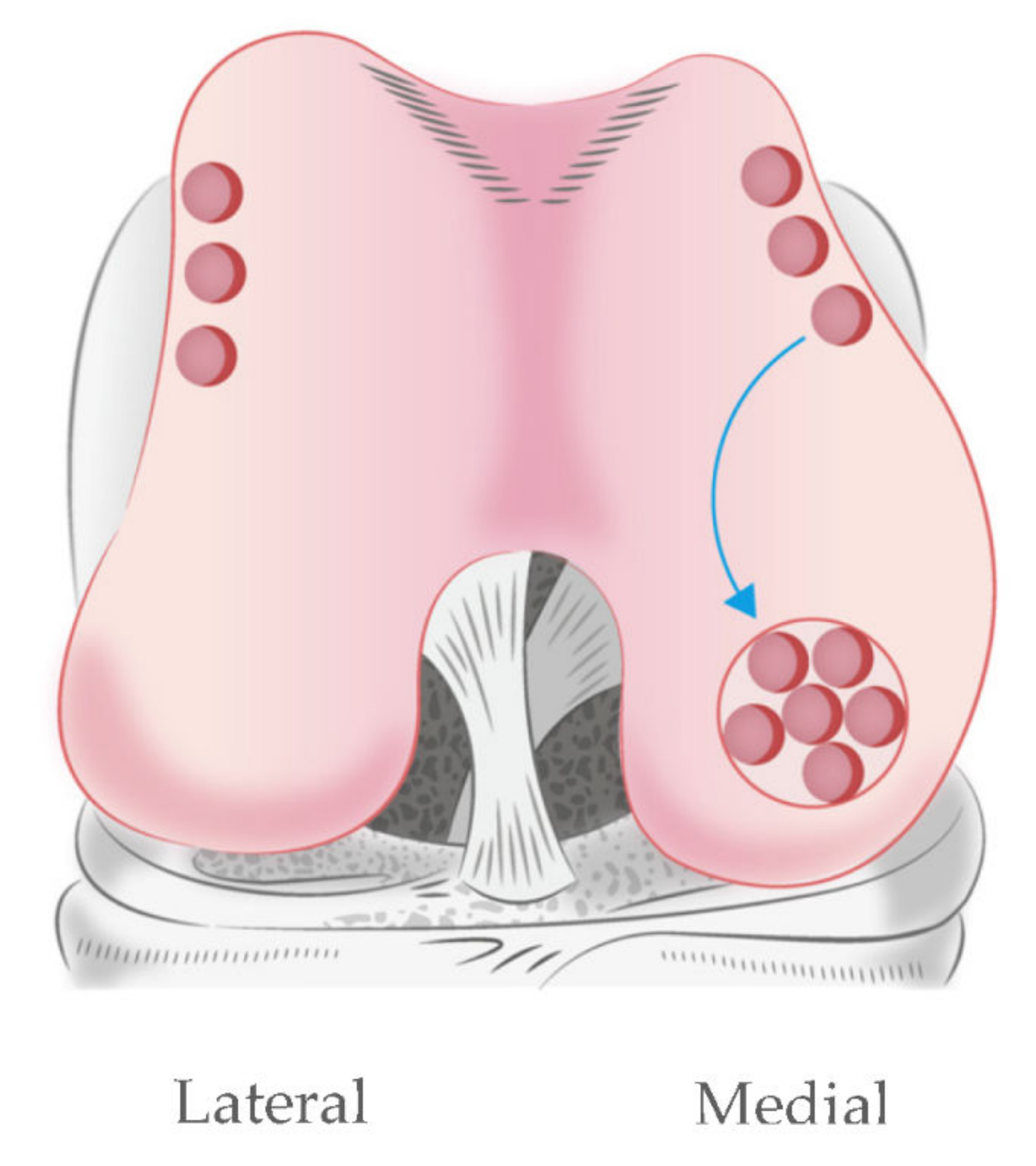
2.1. Sample Collection and Preparation
2.2. Histology and Immunofluorescent Analysis
2.3. Cell Isolation, Culture and Immunocytofluorescent Analysis
2.4. Quantitative Analysis for Gene Expression
2.5. Statistical Analysis
3. Results
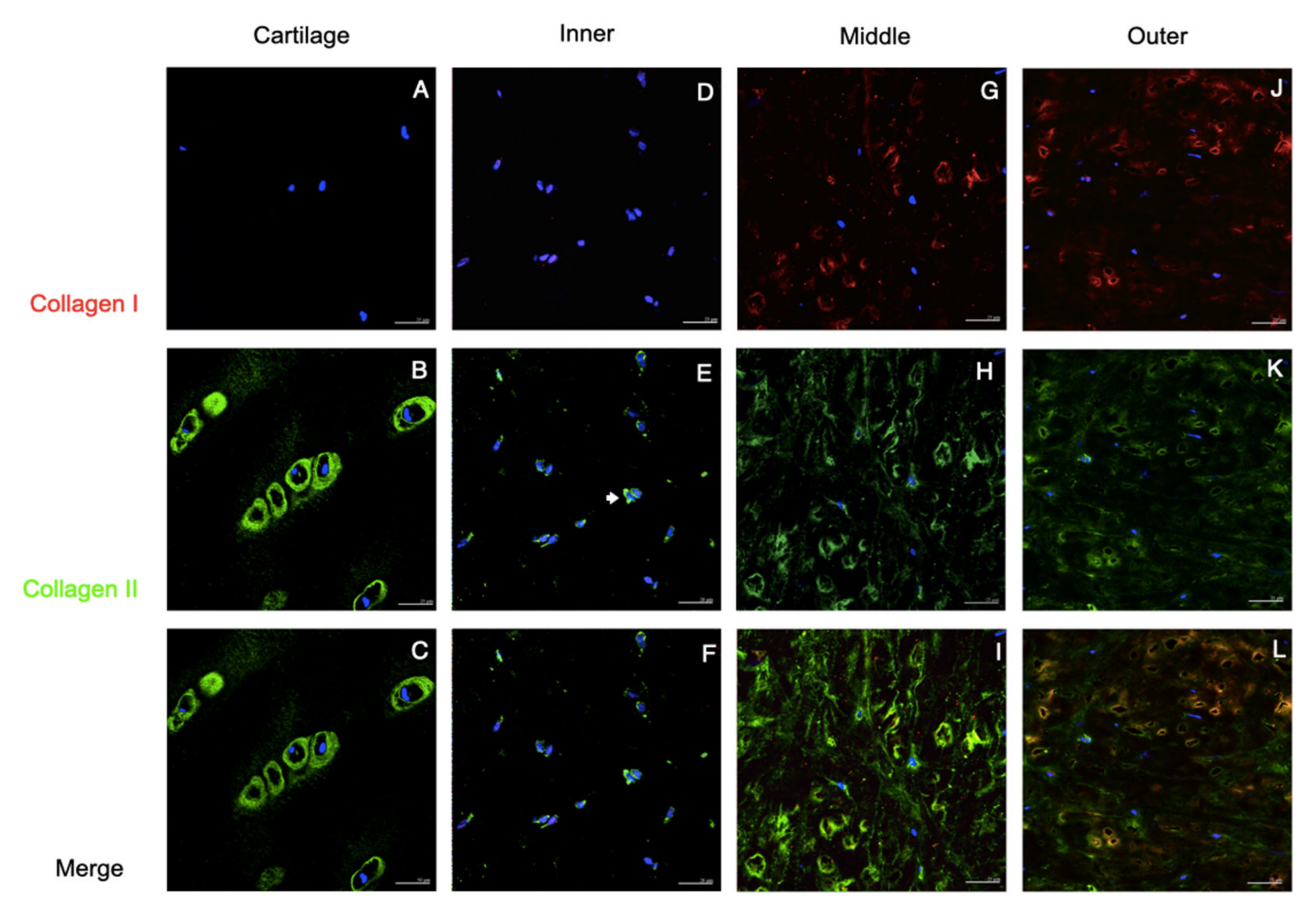
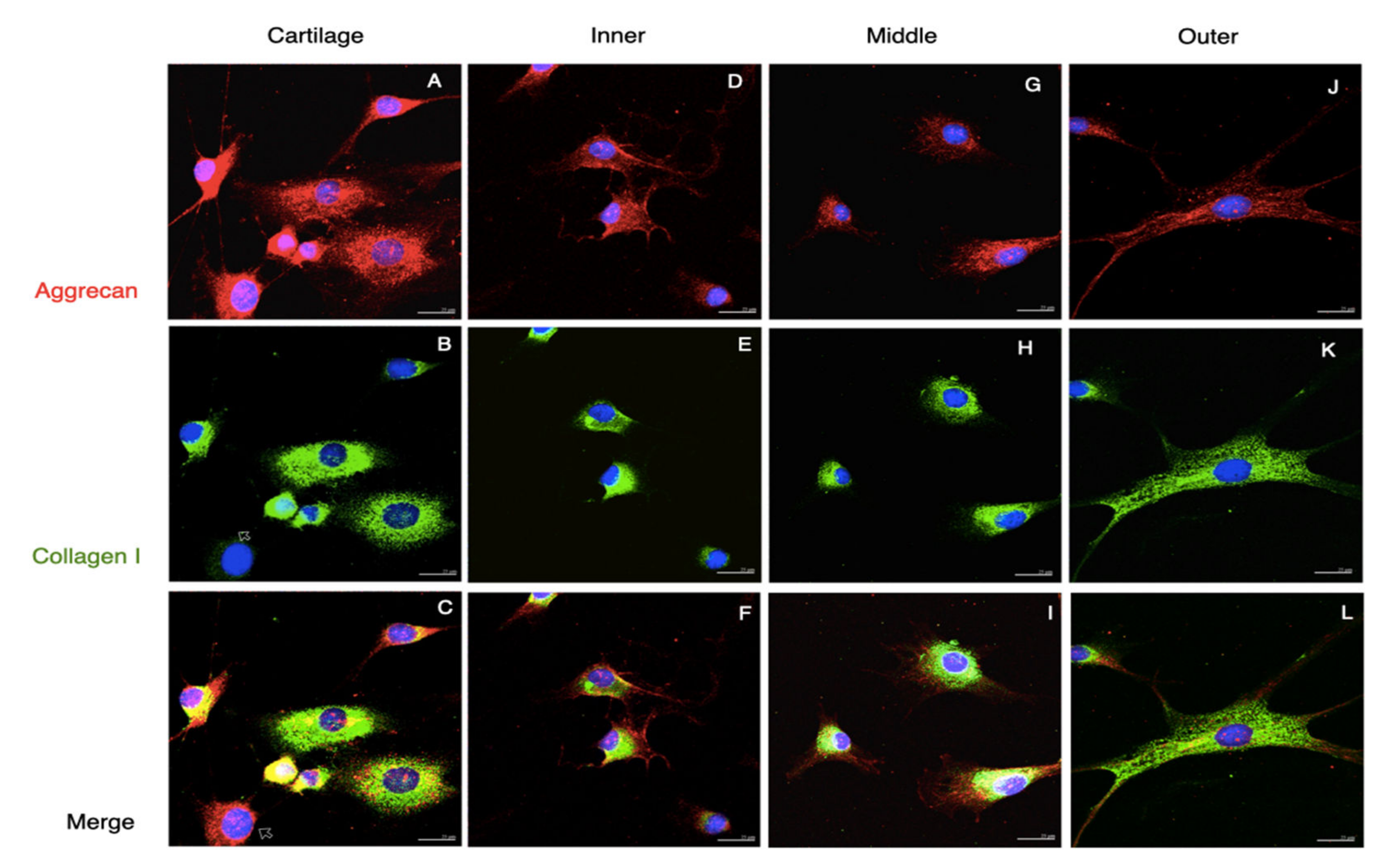
3.1. Tissues and Native Cells
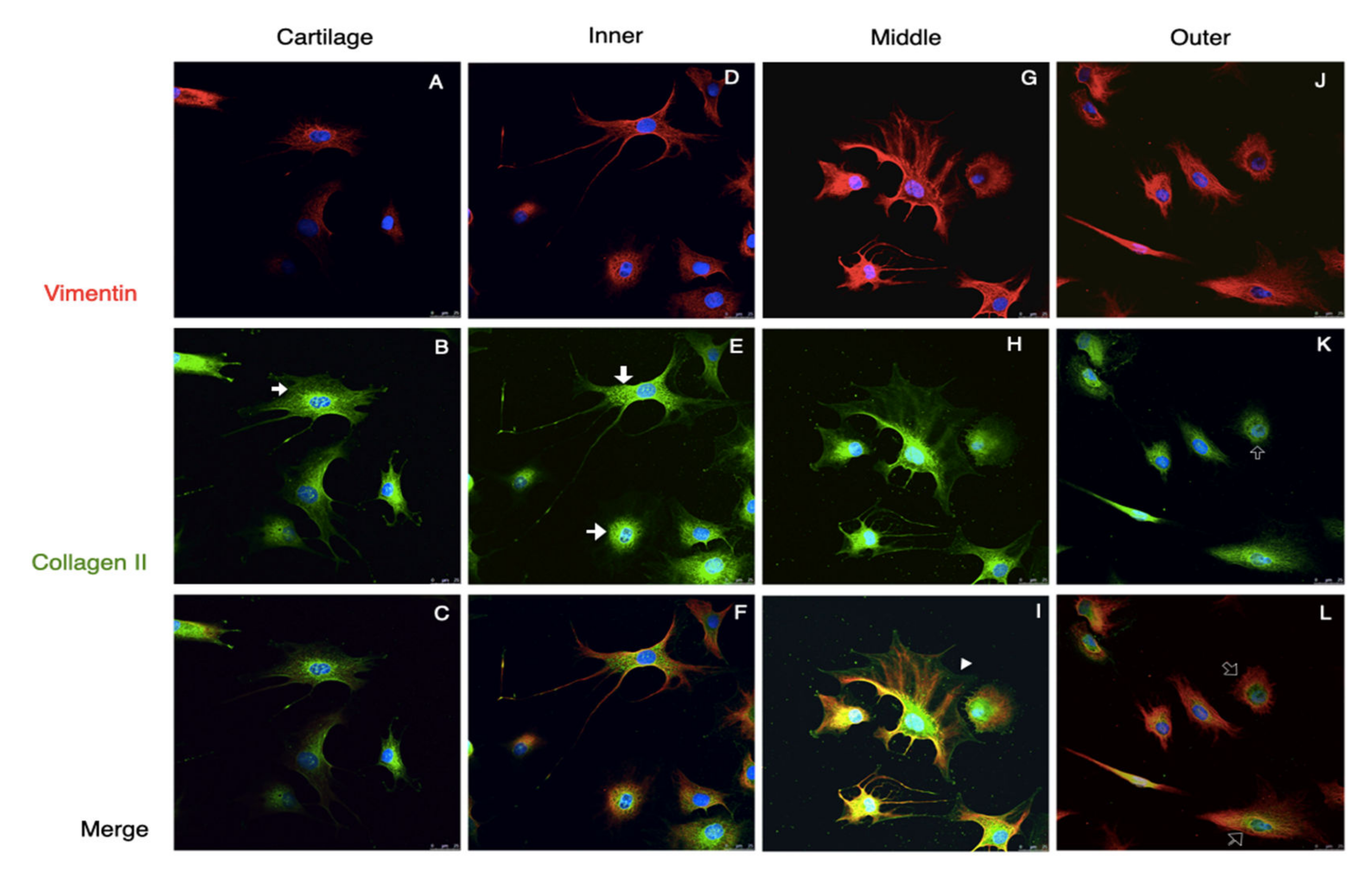
3.2. The Passage 2 Cultured Cells
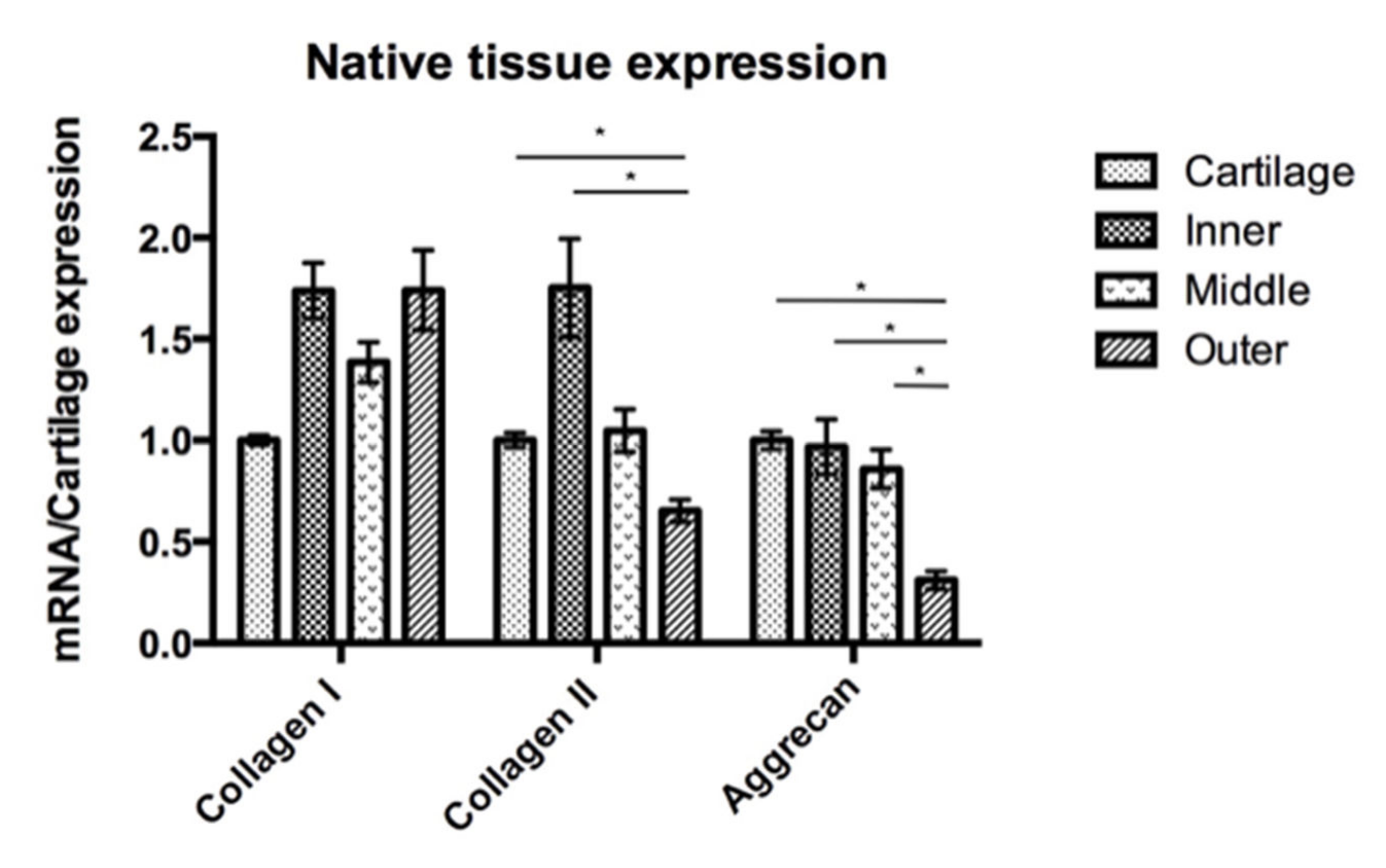
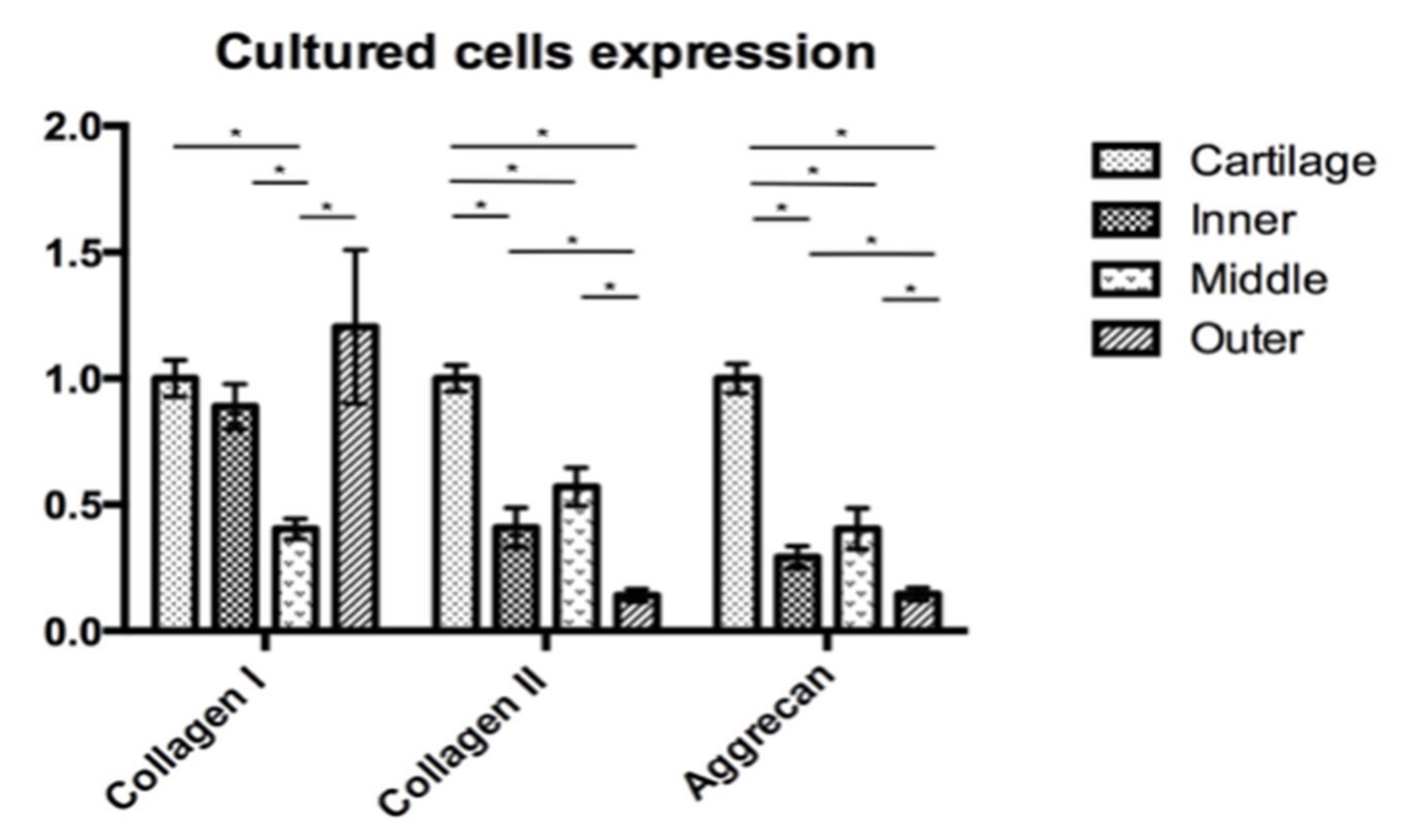
3.3. Gene Expression Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACI | Autologous Chondrocyte Implantation |
| KOOS | Knee injury and Osteoarthritis Outcomes Score |
| PBS | Phosphate-buffered Saline |
| RNA | Ribonucleic Acid |
| BSA | Bovine Serum Albumin |
| FBS | Fetal Bovine Serum |
| COL1A1 | Collagen type 1 alpha 1 gene |
| COL2A1 | Collagen type 2 alpha 1 gene |
| ACAN | Aggrecan gene |
| 18S rRNA | 18S ribosomal RNA |
| MSCs | Mesenchymal Stem Cells |
References
- Spindler, K.P.; Miller, R.R.; Andrish, J.T.; McDevitt, C.A. Comparison of collagen synthesis in the peripheral and central region of the canine meniscus. Clin. Orthop. Relat. Res. 1994, 303, 256–263. [Google Scholar] [CrossRef]
- Newman, A.P.; Daniels, A.U.; Burks, R.T. Principles and decision making in meniscal surgery. Arthroscopy 1993, 9, 33–51. [Google Scholar] [CrossRef]
- Rubman, M.H.; Noyes, F.R.; Barber-Westin, S.D. Arthroscopic repair of meniscal tears that extend into the avascular zone. A review of 198 single and complex tears. Am. J. Sports Med. 1998, 26, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Rath, E.; Richmond, J.C. The menisci: Basic science and advances in treatment. Br. J. Sports Med. 2000, 34, 252–257. [Google Scholar] [CrossRef]
- McDevitt, C.A.; Webber, R.J. The ultrastructure and biochemistry of meniscal cartilage. Clin. Orthop. Relat. Res. 1990, 252, 8–18. [Google Scholar] [CrossRef]
- Noyes, F.R.; Barber-Westin, S.D. Arthroscopic repair of meniscus tears extending into the avascular zone with or without anterior cruciate ligament reconstruction in patients 40 years of age and older. Arthroscopy 2000, 16, 822–829. [Google Scholar] [CrossRef]
- Eggli, S.; Wegmüller, H.; Kosina, J.; Huckell, C.; Jakob, R.P. Long-term results of arthroscopic meniscal repair. An analysis of isolated tears. Am. J. Sports Med. 1995, 23, 715–720. [Google Scholar] [CrossRef]
- Griffin, J.W.; Hadeed, M.M.; Werner, B.C.; Diduch, D.R.; Carson, E.W.; Miller, M.D. Platelet-rich plasma in meniscal repair: Does augmentation improve surgical outcomes? Clin. Orthop. Relat. Res. 2015, 473, 1665–1672. [Google Scholar] [CrossRef]
- Verdonk, P.C.; Forsyth, R.G.; Wang, J.; Almqvist, K.F.; Verdonk, R.; Veys, E.M.; Verbruggen, G. Characterisation of human knee meniscus cell phenotype. Osteoarthr. Cartil. 2005, 13, 548–560. [Google Scholar] [CrossRef]
- Vanderploeg, E.J.; Wilson, C.G.; Imler, S.M.; Ling, C.H.; Levenston, M.E. Regional variations in the distribution and colocalization of extracellular matrix proteins in the juvenile bovine meniscus. J. Anat. 2012, 221, 174–186. [Google Scholar] [CrossRef]
- Scotti, C.; Pozzi, A.; Mangiavini, L.; Vitari, F.; Boschetti, F.; Domeneghini, C.; Fraschini, G.; Peretti, G.M. Healing of meniscal tissue by cellular fibrin glue: An in vivo study. Knee Surg. Sports Traumatol. Arthrosc. 2009, 17, 645–651. [Google Scholar] [CrossRef]
- Fisher, M.B.; Henning, E.A.; Söegaard, N.; Bostrom, M.; Esterhai, J.L.; Mauck, R.L. Engineering meniscus structure and function via multi-layered mesenchymal stem cell-seeded nanofibrous scaffolds. J. Biomech. 2015, 48, 1412–1419. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Yasuda, K.; Kondo, E.; Katsura, T.; Tanabe, Y.; Kimura, M.; Tohyama, H. Implantation of autogenous meniscal fragments wrapped with a fascia sheath enhances fibrocartilage regeneration in vivo in a large harvest site defect. Am. J. Sports Med. 2010, 38, 740–748. [Google Scholar] [CrossRef]
- Shimomura, K.; Bean, A.C.; Lin, H.; Nakamura, N.; Tuan, R.S. In Vitro Repair of Meniscal Radial Tear Using Aligned Electrospun Nanofibrous Scaffold. Tissue Eng. Part A 2015, 21, 2066–2075. [Google Scholar] [CrossRef]
- Ochi, M.; Uchio, Y.; Tobita, M.; Kuriwaka, M. Current concepts in tissue engineering technique for repair of cartilage defect. Artif. Organs 2001, 25, 172–179. [Google Scholar] [CrossRef]
- Cheng, T.; Maddox, N.C.; Wong, A.W.; Rahnama, R.; Kuo, A.C. Comparison of gene expression patterns in articular cartilage and dedifferentiated articular chondrocytes. J. Orthop. Res. 2012, 30, 234–245. [Google Scholar] [CrossRef]
- Chiang, H.; Jiang, C.C. Repair of articular cartilage defects: Review and perspectives. J. Formos Med. Assoc. 2009, 108, 87–101. [Google Scholar] [CrossRef]
- Chang, C.H.; Kuo, T.F.; Lin, C.C.; Chou, C.H.; Chen, K.H.; Lin, F.H.; Liu, H.C. Tissue engineering-based cartilage repair with allogenous chondrocytes and gelatin-chondroitin-hyaluronan tri-copolymer scaffold: A porcine model assessed at 18, 24, and 36 weeks. Biomaterials 2006, 27, 1876–1888. [Google Scholar] [CrossRef]
- Bekkers, J.E.; Bartels, L.W.; Vincken, K.L.; Dhert, W.J.; Creemers, L.B.; Saris, D.B. Articular cartilage evaluation after TruFit plug implantation analyzed by delayed gadolinium-enhanced MRI of cartilage (dGEMRIC). Am. J. Sports Med. 2013, 41, 1290–1295. [Google Scholar] [CrossRef]
- Chiang, H.; Liao, C.J.; Hsieh, C.H.; Shen, C.Y.; Huang, Y.Y.; Jiang, C.C. Clinical feasibility of a novel biphasic osteochondral composite for matrix-associated autologous chondrocyte implantation. Osteoarthr. Cartil. 2013, 21, 589–598. [Google Scholar] [CrossRef]
- Hoemann, C.D.; A Kandel, R.; Roberts, S.; Saris, D.B.; Creemers, L.; Mainil-Varlet, P.; Méthot, S.; Hollander, A.P.; Buschmann, M. International Cartilage Repair Society (ICRS) Recommended Guidelines for Histological Endpoints for Cartilage Repair Studies in Animal Models and Clinical Trials. Cartilage 2011, 2, 153–172. [Google Scholar] [CrossRef]
- Wilusz, R.E.; Sanchez-Adams, J.; Guilak, F. The structure and function of the pericellular matrix of articular cartilage. Matrix Biol. 2014, 39, 25–32. [Google Scholar] [CrossRef]
- Chevrier, A.; Nelea, M.; Hurtig, M.B.; Hoemann, C.D.; Buschmann, M.D. Meniscus structure in human, sheep, and rabbit for animal models of meniscus repair. J. Orthop. Res. 2009, 27, 1197–1203. [Google Scholar] [CrossRef]
- Chandran, P.L.; Horkay, F. Aggrecan, an unusual polyelectrolyte: Review of solution behavior and physiological implications. Acta Biomater. 2012, 8, 3–12. [Google Scholar] [CrossRef]
- Rodeo, S.; Kawamura, S.; Einhorn, T.; O’Keefe, R.; Buckwalter, J. Form and function of the meniscus. In Orthopaedic Basic Science, 3rd ed.; American Academy of Orthopaedic Surgeons: Rosemont, IL, USA, 2007; pp. 175–189. [Google Scholar]
- Kambic, H.E.; McDevitt, C.A. Spatial organization of types I and II collagen in the canine meniscus. J. Orthop. Res. 2005, 23, 142–149. [Google Scholar] [CrossRef]
- Furumatsu, T.; Kanazawa, T.; Yokoyama, Y.; Abe, N.; Ozaki, T. Inner meniscus cells maintain higher chondrogenic phenotype compared with outer meniscus cells. Connect. Tissue Res. 2011, 52, 459–465. [Google Scholar] [CrossRef]
- Mauck, R.L.; Burdick, J.A. From repair to regeneration: Biomaterials to reprogram the meniscus wound microenvironment. Ann. Biomed. Eng. 2015, 43, 529–542. [Google Scholar] [CrossRef][Green Version]
- Mauck, R.L.; Martinez-Diaz, G.J.; Yuan, X.; Tuan, R.S. Regional multilineage differentiation potential of meniscal fibrochondrocytes: Implications for meniscus repair. Anat. Rec. 2007, 290, 48–58. [Google Scholar] [CrossRef]
- Kuo, C.K.; Li, W.J.; Mauck, R.L.; Tuan, R.S. Cartilage tissue engineering: Its potential and uses. Curr. Opin. Rheumatol. 2006, 18, 64–73. [Google Scholar] [CrossRef]
- Guilak, F.; Alexopoulos, L.G.; Upton, M.L.; Youn, I.; Choi, J.B.; Cao, L.; Setton, L.A.; Haider, M.A. The pericellular matrix as a transducer of biomechanical and biochemical signals in articular cartilage. Ann. N. Y. Acad. Sci. 2006, 1068, 498–512. [Google Scholar] [CrossRef]
- Ruoslahti, E.; Yamaguchi, Y. Proteoglycans as modulators of growth factor activities. Cell 1991, 64, 867–869. [Google Scholar] [CrossRef]
- Larson, C.M.; Kelley, S.S.; Blackwood, A.D.; Banes, A.J.; Lee, G.M. Retention of the native chondrocyte pericellular matrix results in significantly improved matrix production. Matrix Biol. 2002, 21, 349–359. [Google Scholar] [CrossRef]
- Garretson, R.B., III; Katolik, L.I.; Verma, N.; Beck, P.R.; Bach, B.R.; Cole, B.J. Contact pressure at osteochondral donor sites in the patellofemoral joint. Am. J. Sports Med. 2004, 32, 967–974. [Google Scholar] [CrossRef] [PubMed]
- Hangody, L.; Vásárhelyi, G.; Hangody, L.R.; Sükösd, Z.; Tibay, G.; Bartha, L.; Bodó, G. Autologous osteochondral grafting--technique and long-term results. Injury 2008, 39 (Suppl. 1), S32–S39. [Google Scholar] [CrossRef]
- Ghadially, F.N.; Thomas, I.; Yong, N.; Lalonde, J.M. Ultrastructure of rabbit semilunar cartilages. J. Anat. 1978, 125, 499–517. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Antiserum | Cell Type | Titer |
|---|---|---|---|
| Aggrecan | Chondrocyte | 1:100 | |
| Collagen I | Fibroblast | 1:200 | |
| Collagen II | Chondrocyte | 1:100 | |
| Vimentin | Fibroblast | 1:100 | |
| Mouse IgG | 1:200 | ||
| Rabbit IgG | 1:200 |
| Genes | Pairs | Primer Sequence (5′ to 3′) |
|---|---|---|
| Type I collagen (COL1A1) | Sense Antisense | GGAGGAGAGTCAGGAAGG GCAACACAGTTACACAAGG |
| Type II collagen (COL2A1) | Sense Antisense | GGCAGAGGTATAATGATAAG ATGTCGTCGCAGAGG |
| Aggrecan (ACAN) | Sense Antisense | ATACCGTCGTAGTTCC TCCTTGTCTCCATAGC |
| 18S rRNA | Sense Antisense | ATACCGTCGTAGTTCC GTCTCGTTCGTTATCG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Numpaisal, P.-o.; Jiang, C.-C.; Hsieh, C.-H.; Chiang, H.; Chien, C.-L. Prospective Application of Partially Digested Autologous Chondrocyte for Meniscus Tissue Engineering. Pharmaceutics 2022, 14, 605. https://doi.org/10.3390/pharmaceutics14030605
Numpaisal P-o, Jiang C-C, Hsieh C-H, Chiang H, Chien C-L. Prospective Application of Partially Digested Autologous Chondrocyte for Meniscus Tissue Engineering. Pharmaceutics. 2022; 14(3):605. https://doi.org/10.3390/pharmaceutics14030605
Chicago/Turabian StyleNumpaisal, Piya-on, Ching-Chuan Jiang, Chang-Hsun Hsieh, Hongsen Chiang, and Chung-Liang Chien. 2022. "Prospective Application of Partially Digested Autologous Chondrocyte for Meniscus Tissue Engineering" Pharmaceutics 14, no. 3: 605. https://doi.org/10.3390/pharmaceutics14030605
APA StyleNumpaisal, P.-o., Jiang, C.-C., Hsieh, C.-H., Chiang, H., & Chien, C.-L. (2022). Prospective Application of Partially Digested Autologous Chondrocyte for Meniscus Tissue Engineering. Pharmaceutics, 14(3), 605. https://doi.org/10.3390/pharmaceutics14030605