
Encapsulation of Asparaginase as a Promising Strategy to Improve In Vivo Drug Performance

 , and
, and
Abstract
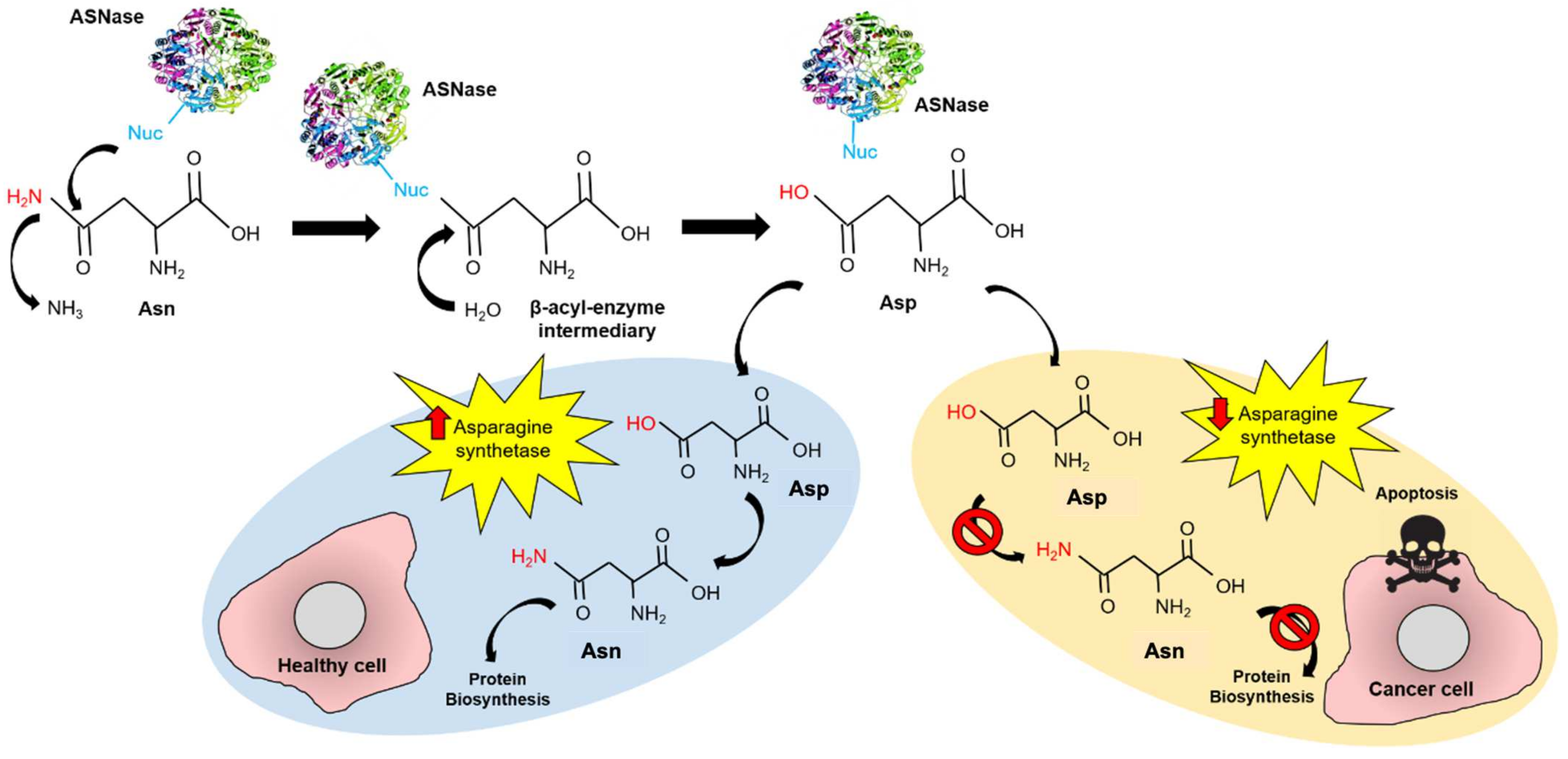
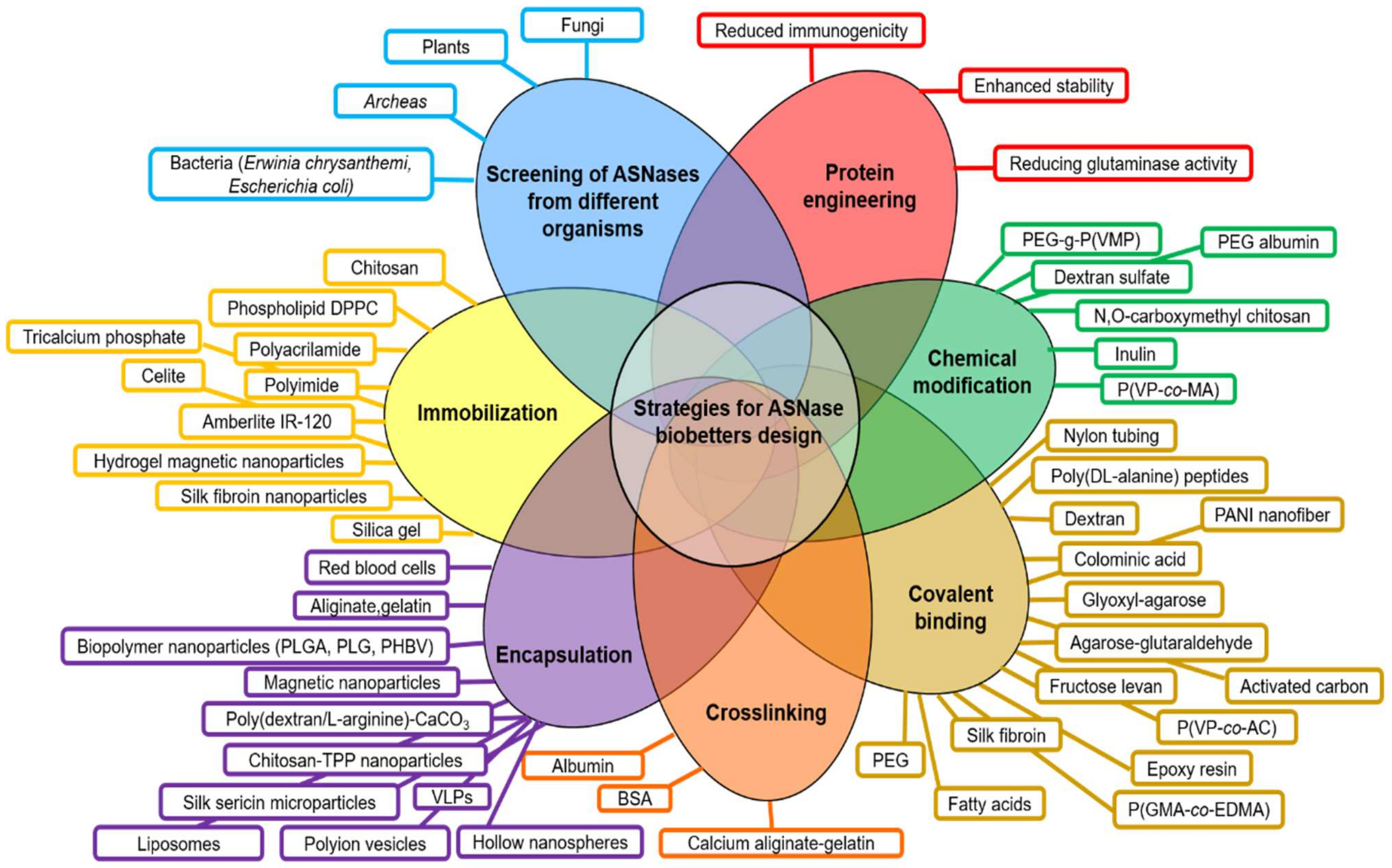
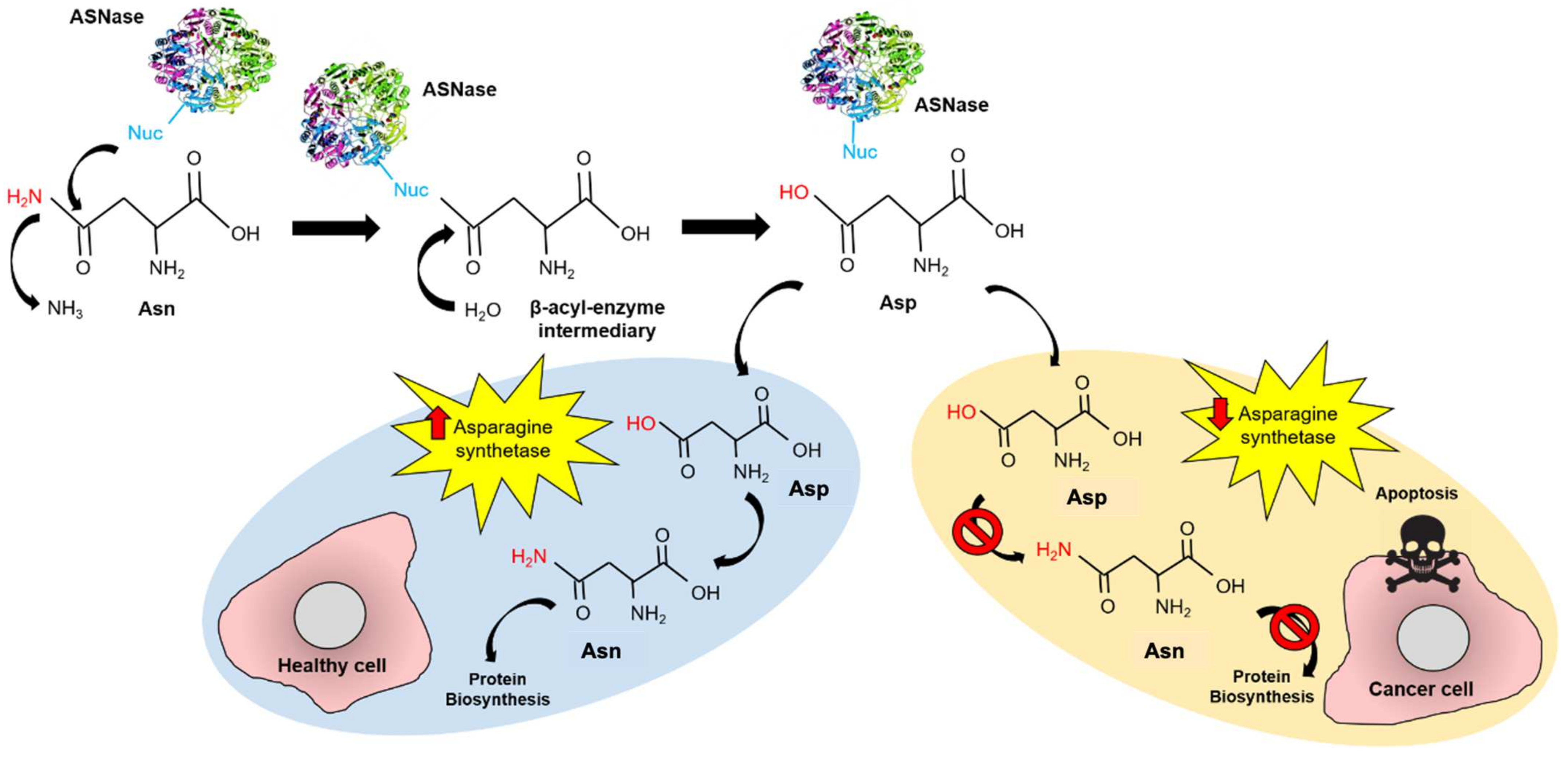
:1. Introduction
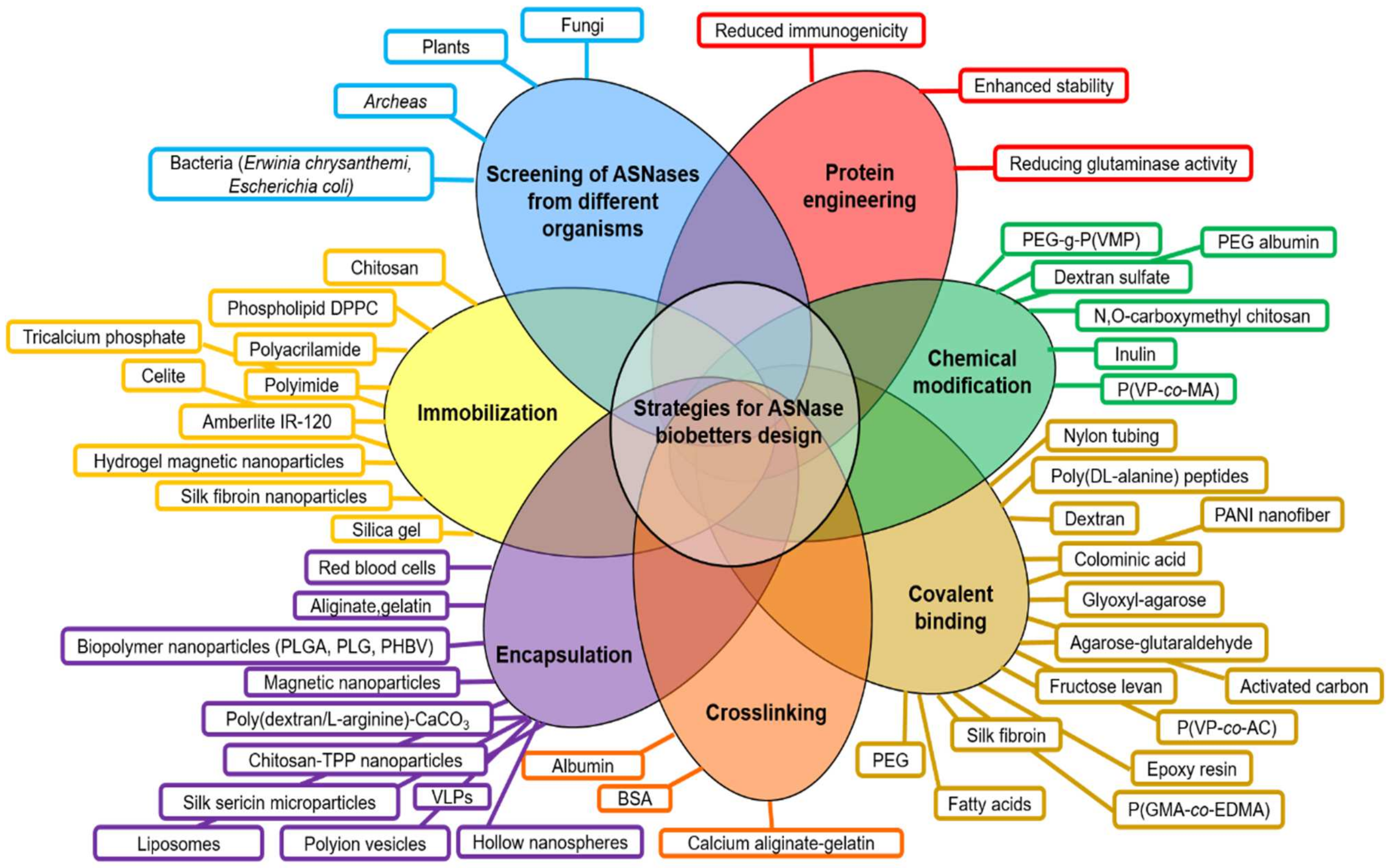
- (a)
- Screening of ASNases from different microorganisms (plant, fungi, bacteria, archaea).
- (b)
- Protein engineering mainly focused on reducing the glutaminase cross-reactivity, enhancement of enzyme stability and reducing immunogenicity.
- (c)
- Chemical modifications (i.e., crosslinking, covalent binding).
- (d)
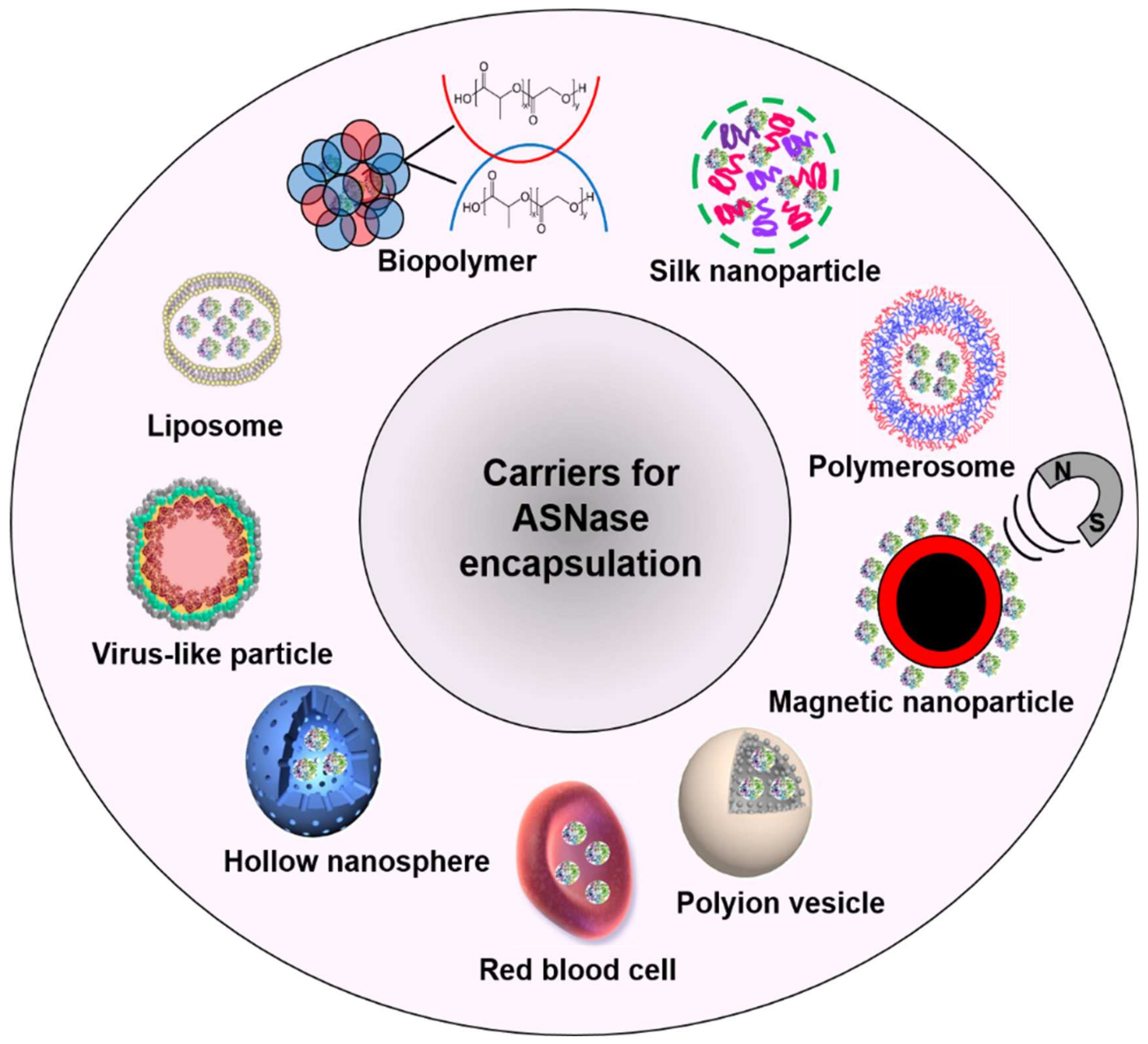
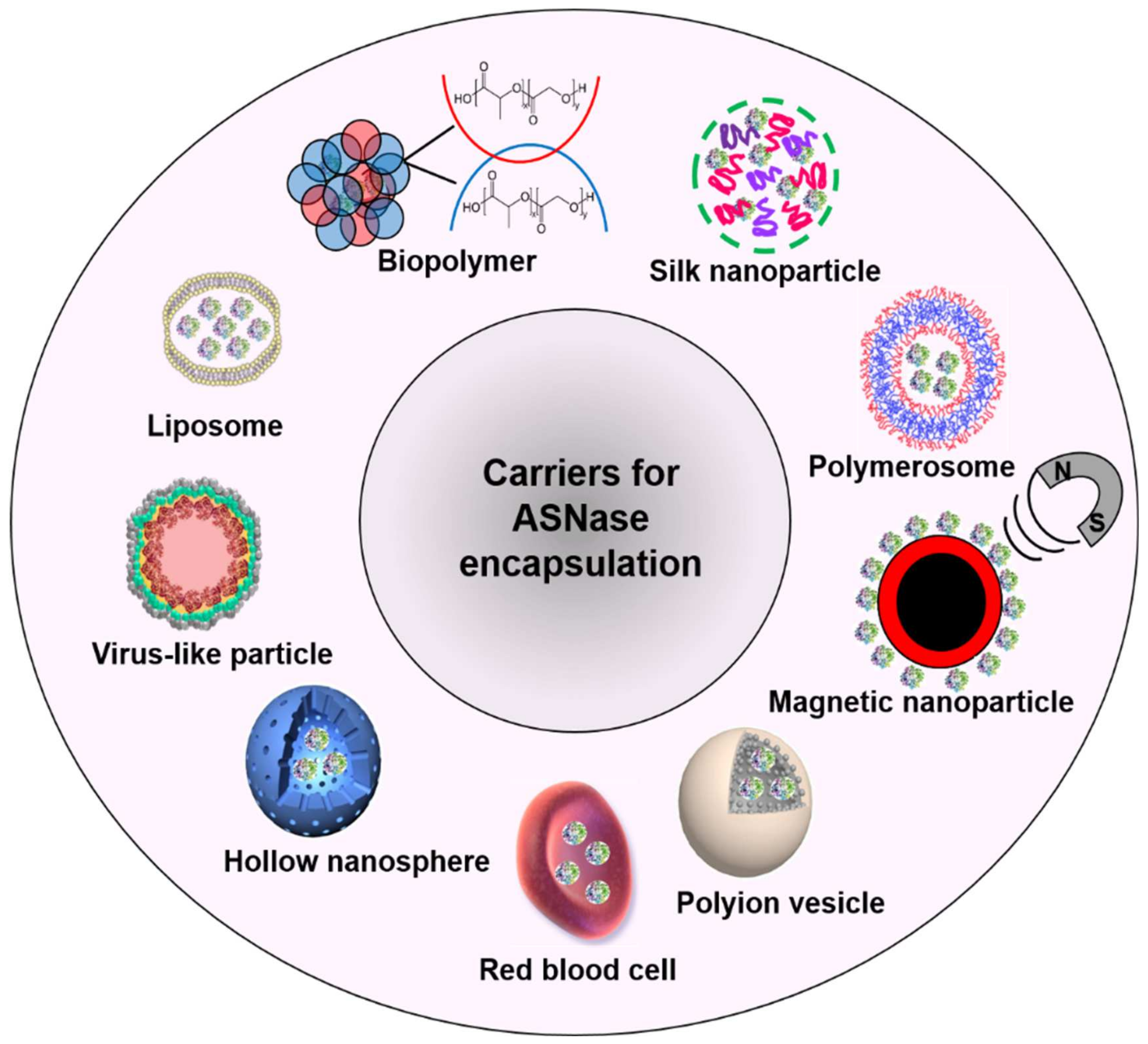
2. Carriers for ASNase Encapsulation
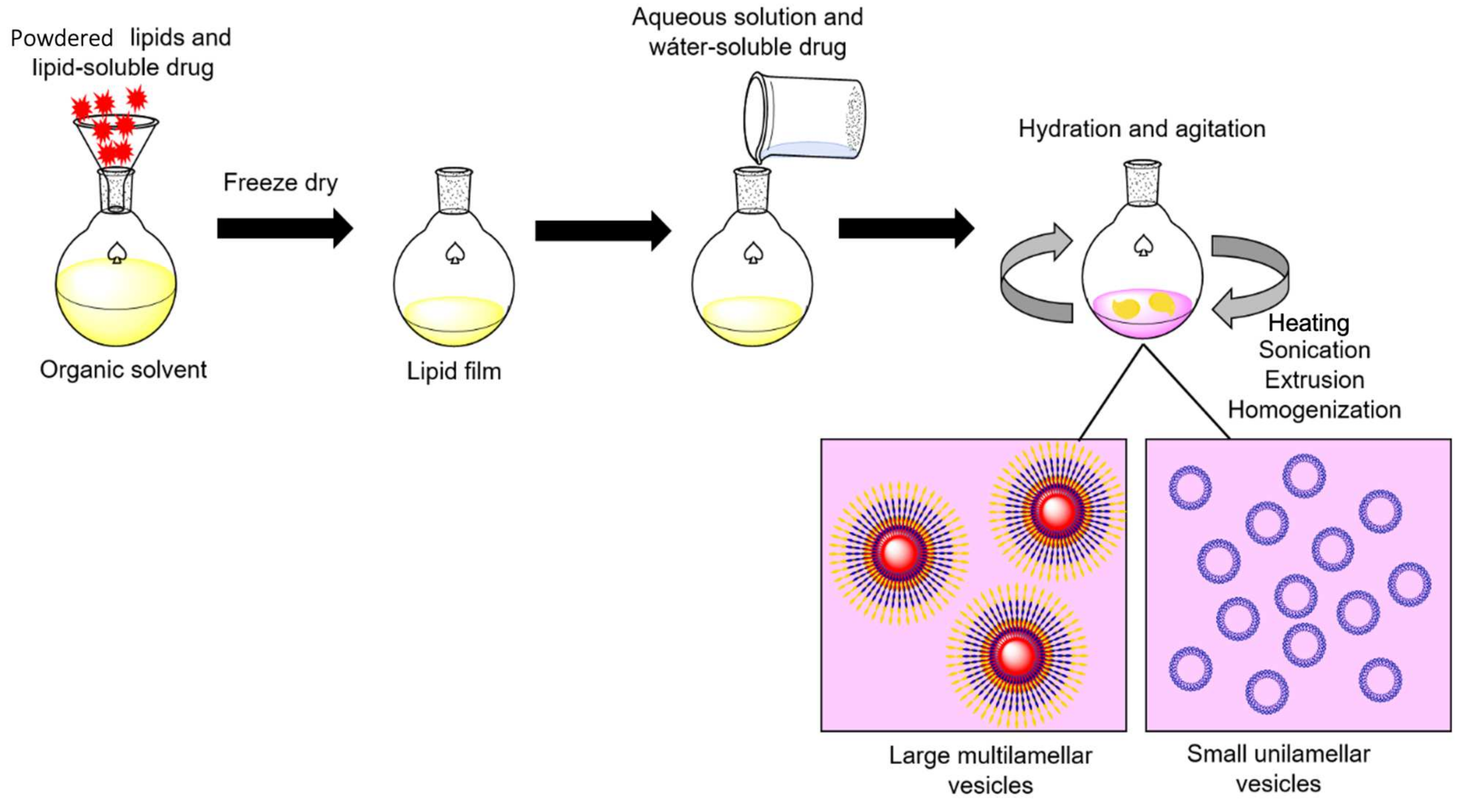
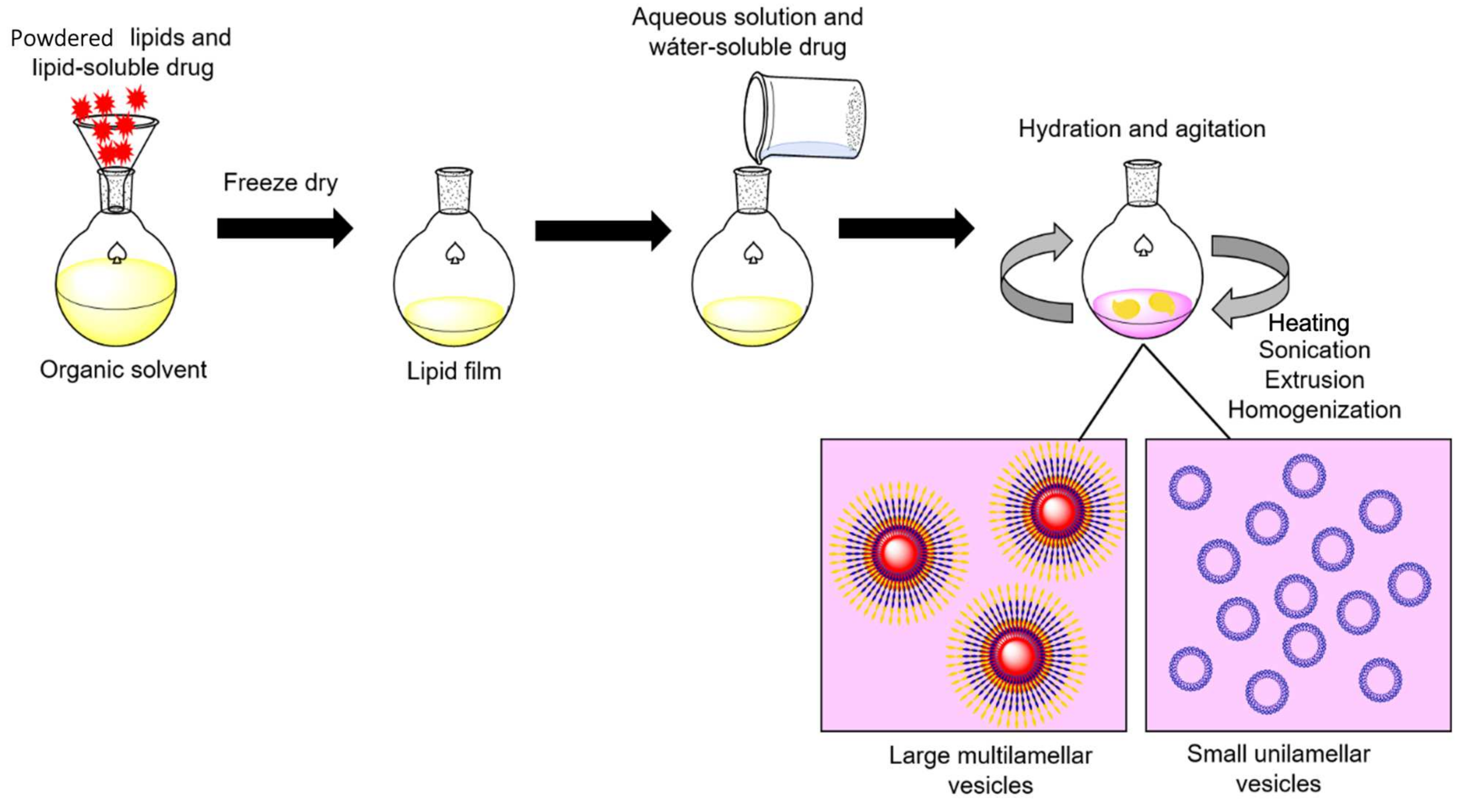
3. Liposome-ASNase
4. Liposome Size Influences Blood-Circulating Time
5. Biopolymer-ASNase
6. What Should Be Considered before Using a Biopolymer-Based Carrier for ASNase Encapsulation?
7. Silk-ASNase
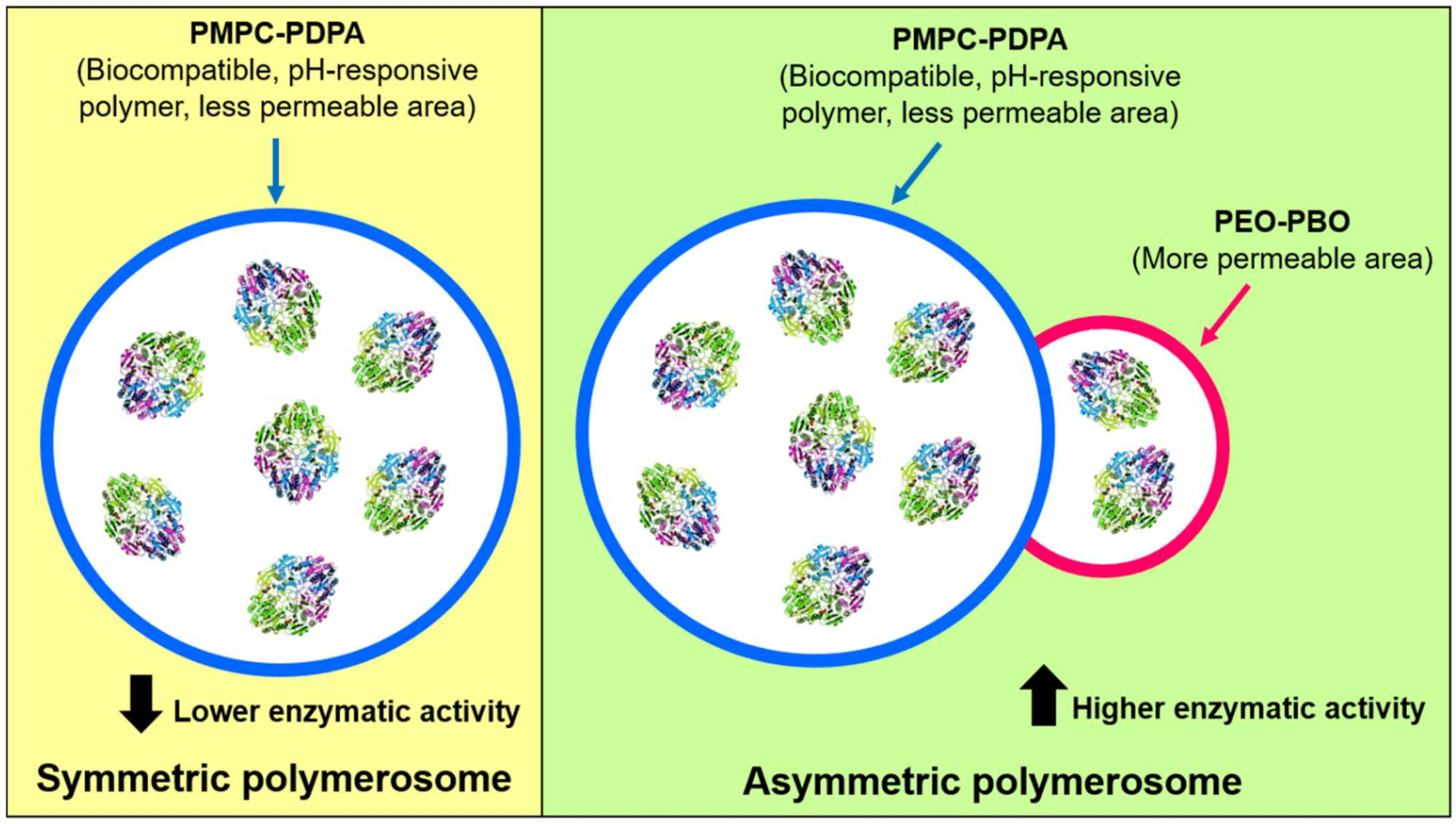
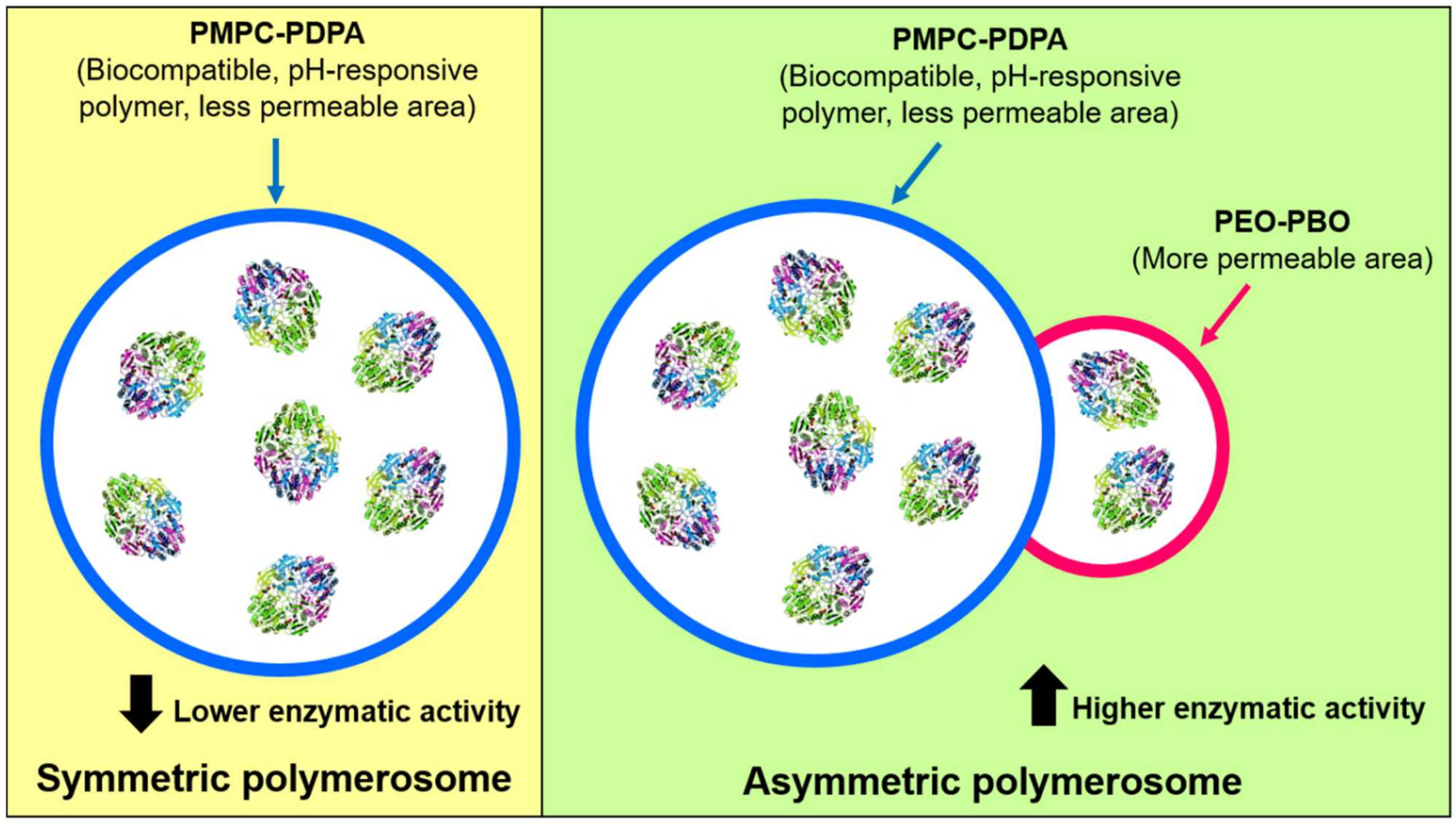
8. Polymerosome-ASNase
9. Magnetic Nanoparticle-ASNase
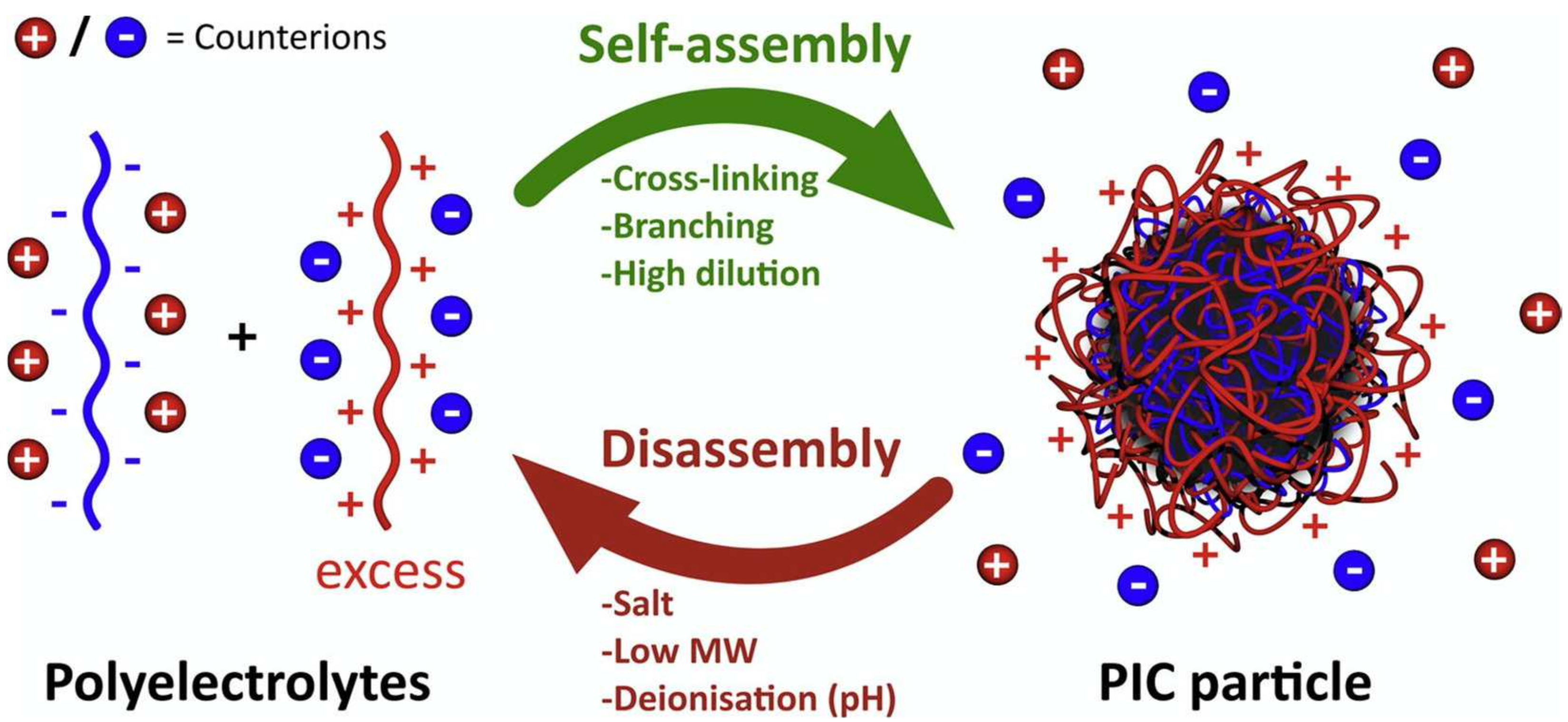
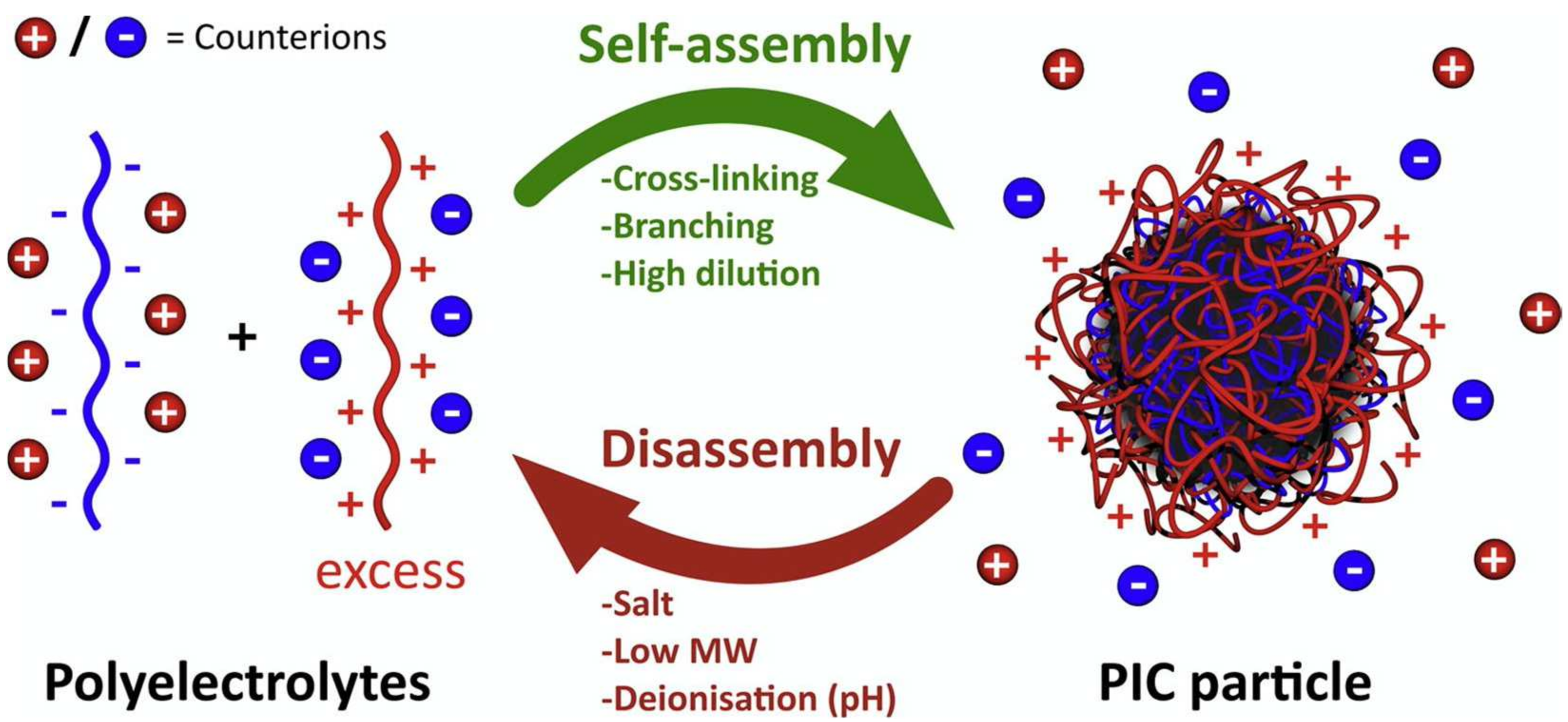
10. Polyion Complex Vesicles-ASNase
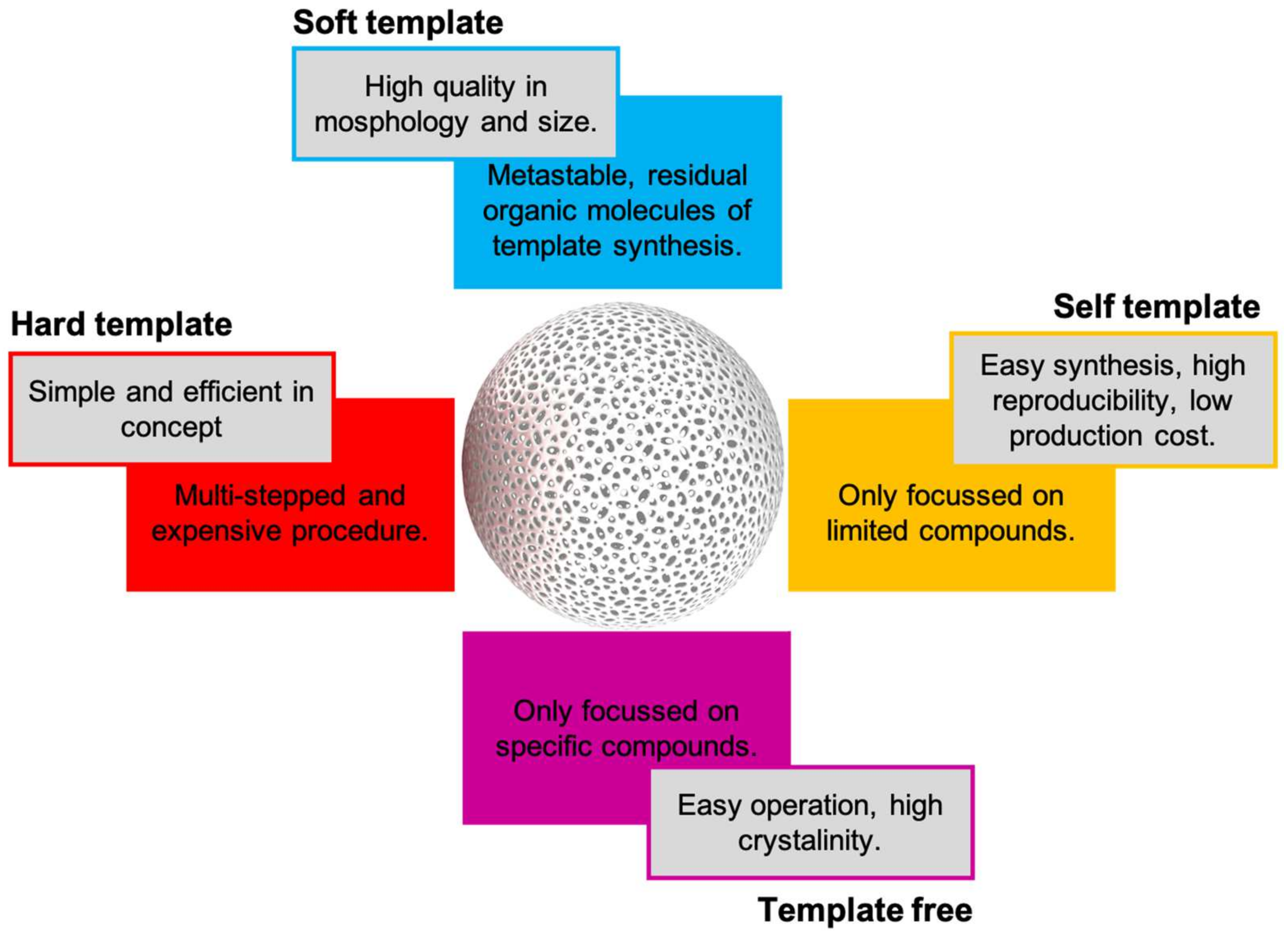
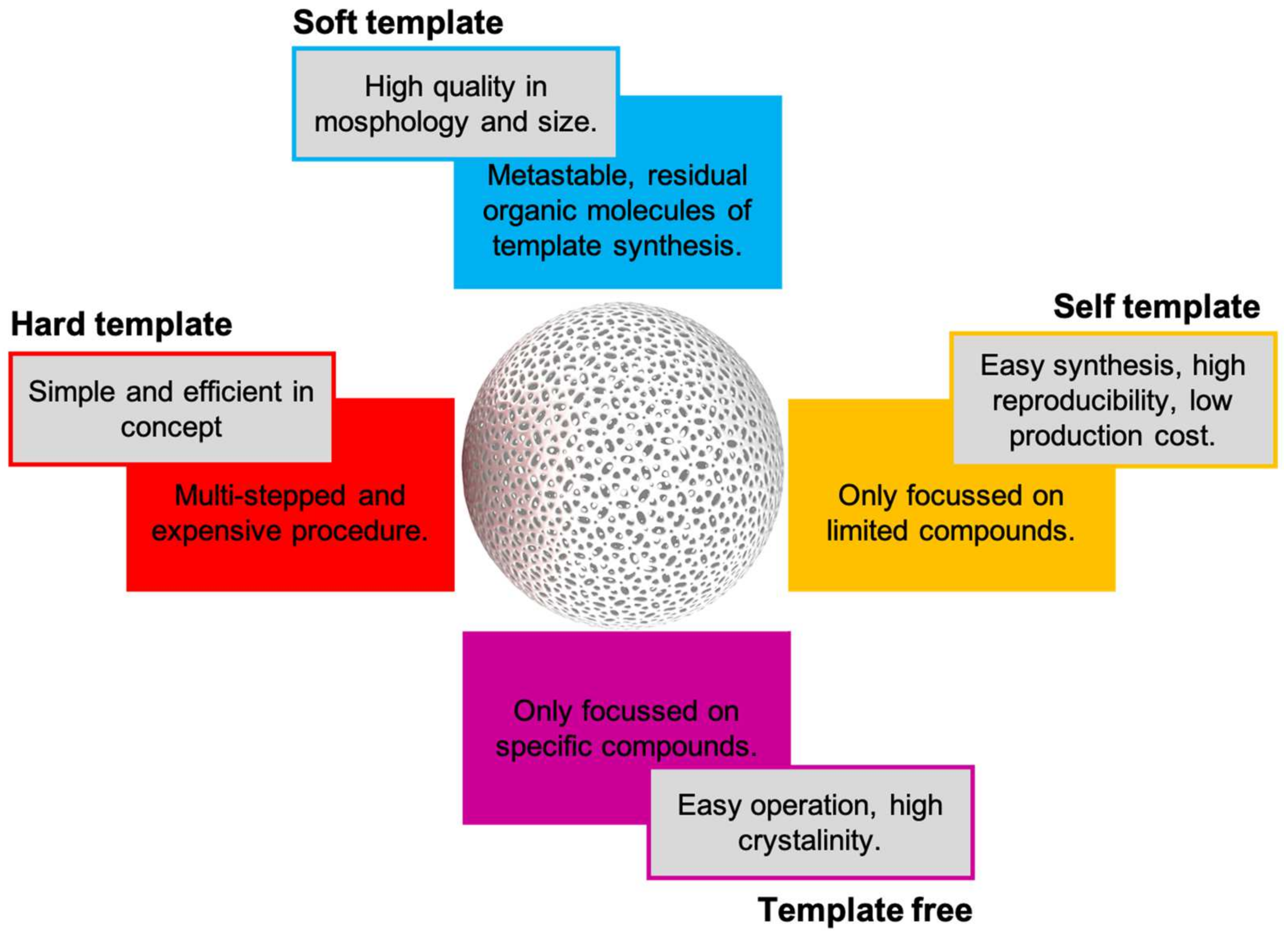
11. Hollow Nanospheres-ASNase
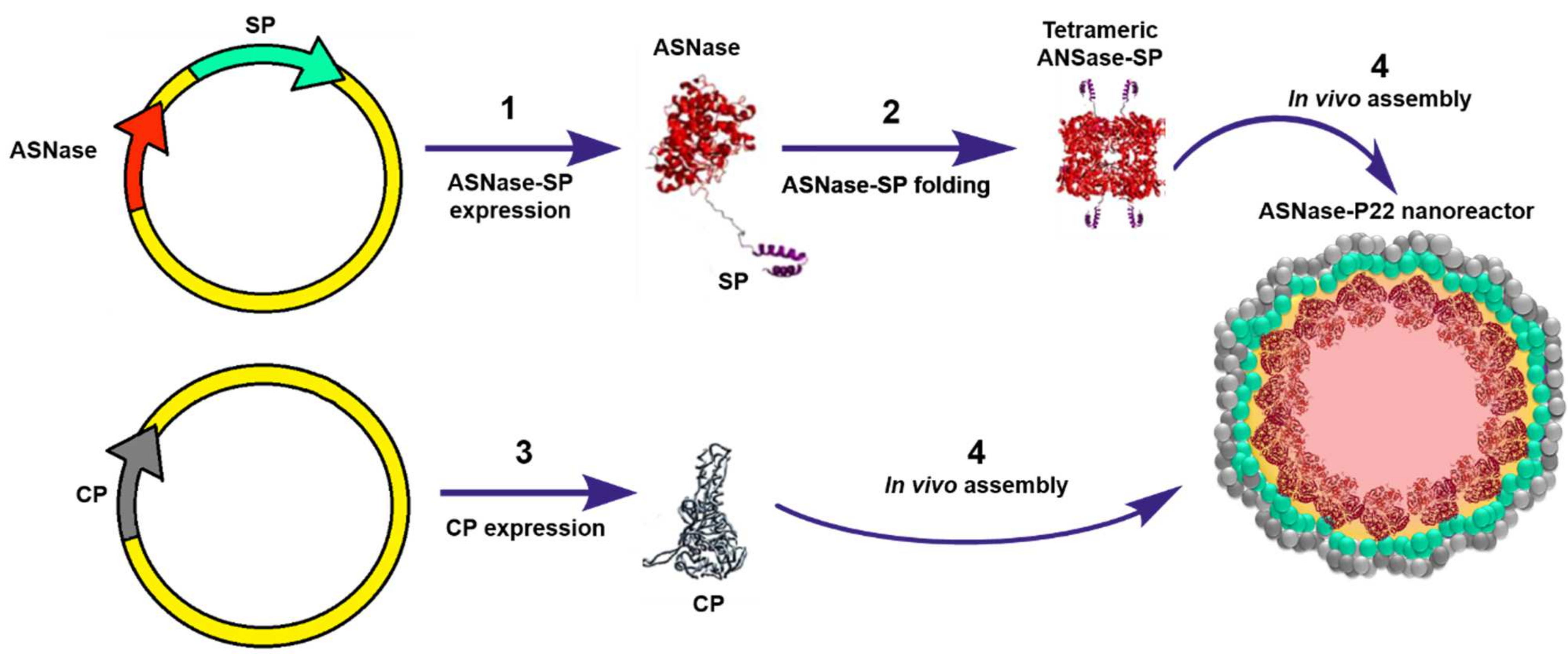
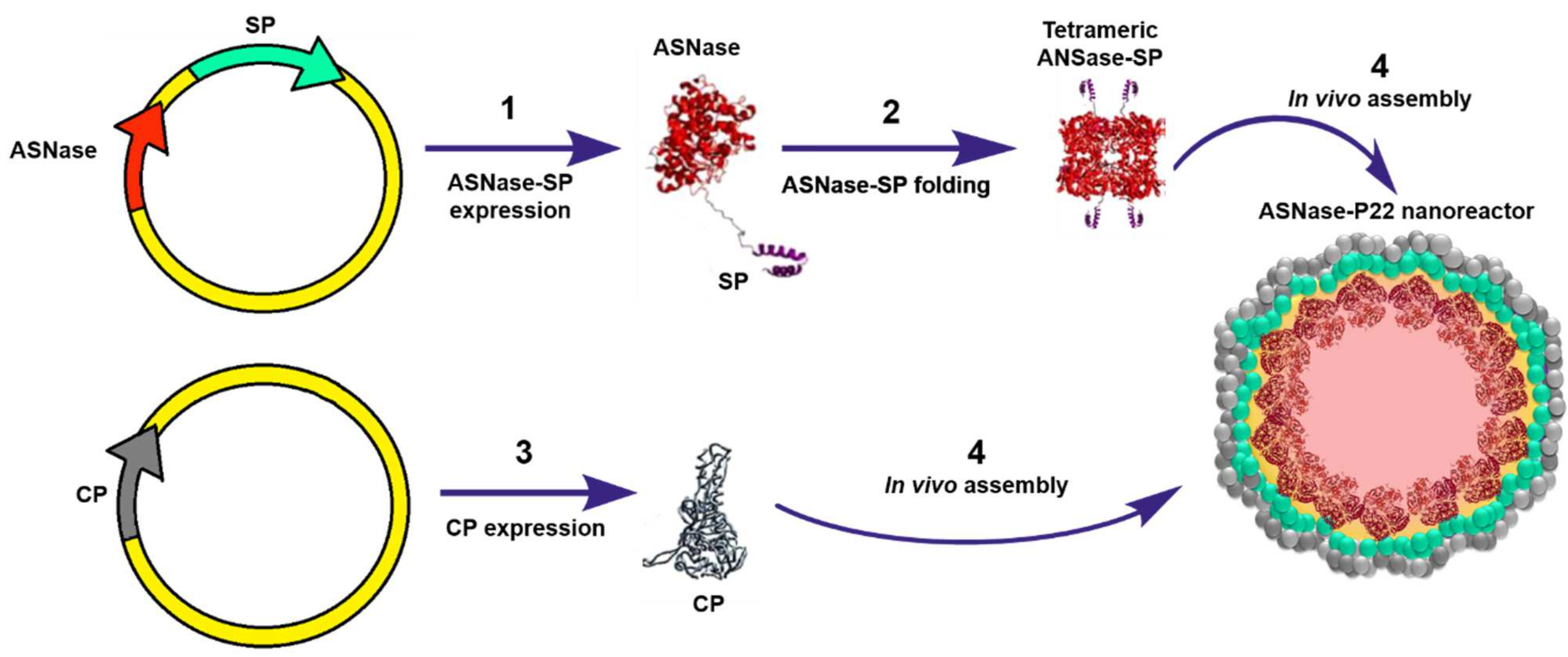
12. Virus-Like Particles-ASNase
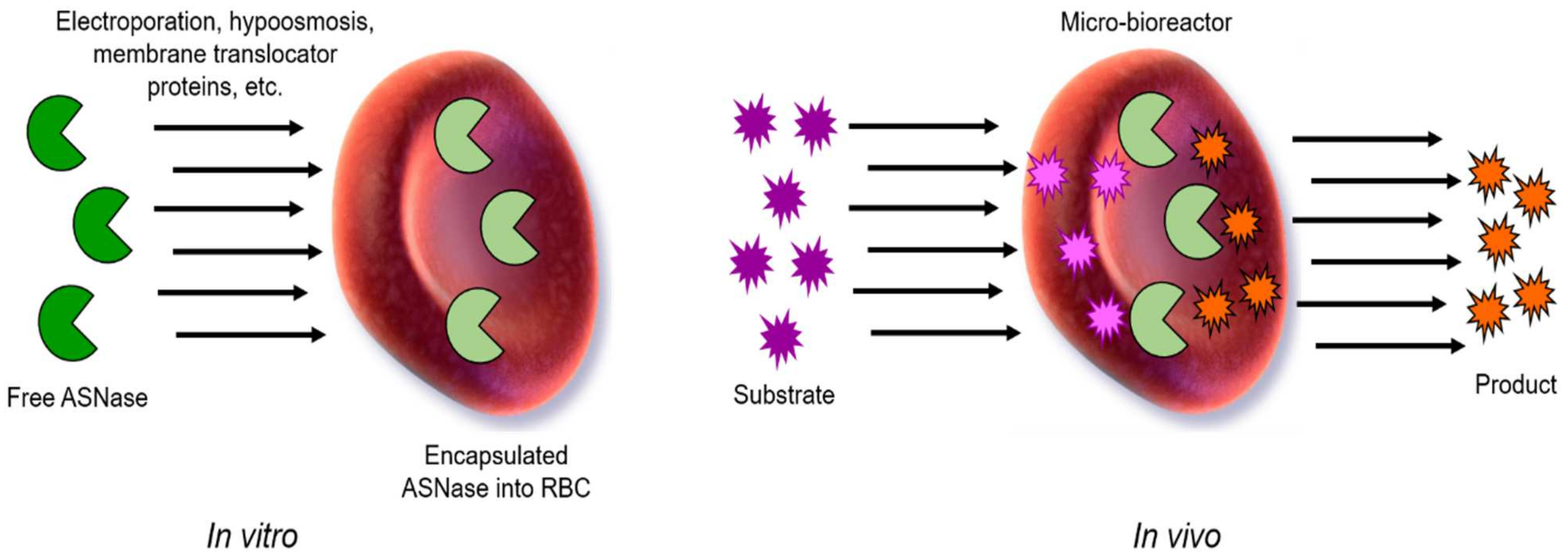
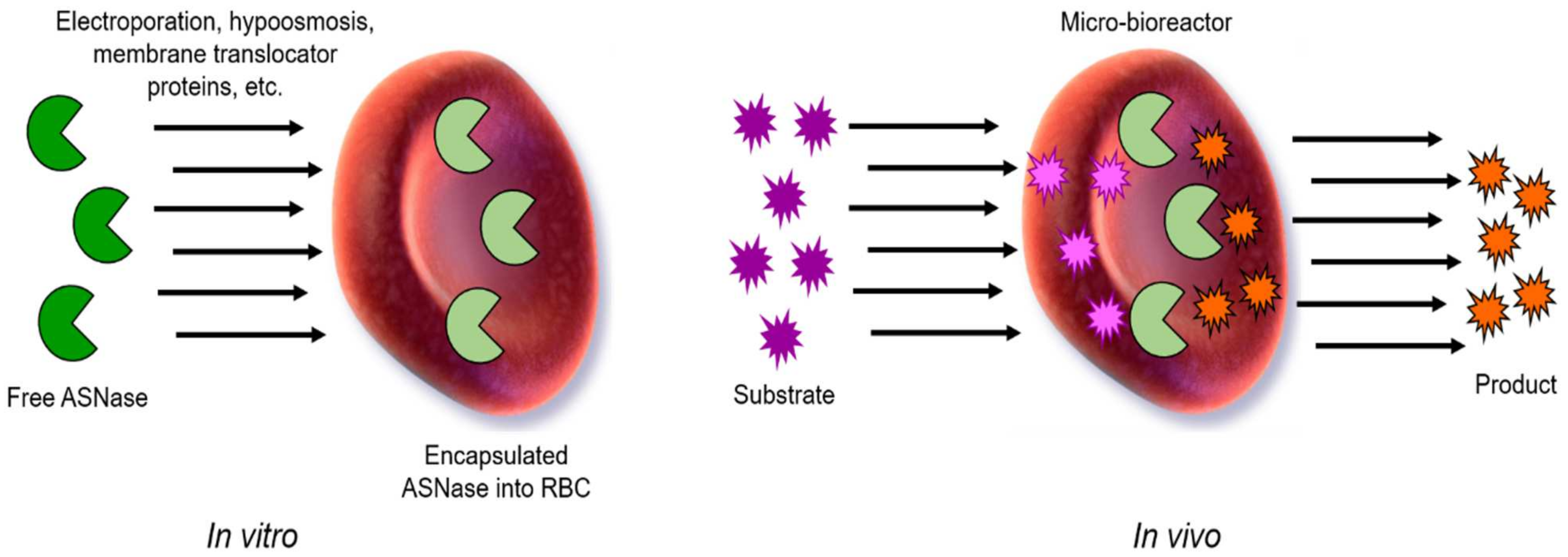
13. Red Blood Cell-ASNase
14. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Malard, F.; Mohty, M. Acute lymphoblastic leukaemia. Lancet 2020, 395, 1146–1162. [Google Scholar] [CrossRef]
- Kato, M.; Manabe, A. Treatment and biology of pediatric acute lymphoblastic leukemia. Pediatr. Int. 2018, 60, 4–12. [Google Scholar] [CrossRef]
- Broome, J. Evidence that the L-Asparaginase Activity of Guinea Pig Serum is responsible for its Antilymphoma Effects. Nature 1961, 191, 1114–1115. [Google Scholar] [CrossRef]
- Schrek, R.; Dolowy, W.C.; Ammeraal, R.N. L-asparaginase: Toxicity to normal and leukemic human lymphocytes. Science 1967, 155, 329–330. [Google Scholar] [CrossRef] [PubMed]
- Bussolati, O.; Belletti, S.; Uggeri, J.; Gatti, R.; Orlandini, G.; Dall’Asta, V.; Gazzola, G.C. Characterization of apoptotic phenomena induced by treatment with L-asparaginase in NIH3T3 cells. Exp. Cell Res. 1995, 220, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Ueno, T.; Ohtawa, K.; Mitsui, K.; Kodera, Y.; Hiroto, M.; Matsushima, A.; Inada, Y.; Nishimura, H. Cell cycle arrest and apoptosis of leukemia cells induced by L-asparaginase. Leukemia 1997, 11, 1858–1861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terwilliger, T.; Abdul-Hay, M. Acute lymphoblastic leukemia: A comprehensive review and 2017 update. Blood Cancer J. 2017, 7, e577. [Google Scholar] [CrossRef] [Green Version]
- Boissel, N.; Auclerc, M.F.; Lhéritier, V.; Perel, Y.; Thomas, X.; Leblanc, T.; Rousselot, P.; Cayuela, J.M.; Gabert, J.; Fegueux, N.; et al. Should adolescents with acute lymphoblastic leukemia be treated as old children or young adults? Comparison of the French FRALLE-93 and LALA-94 trials. J. Clin. Oncol. 2003, 21, 774–780. [Google Scholar] [CrossRef] [PubMed]
- De Bont, J.M.; Holt, B.; Dekker, A.W.; van der Does-van den Berg, A.; Sonneveld, P.; Pieters, R. Significant difference in outcome for adolescents with acute lymphoblastic leukemia treated on pediatric vs adult protocols in the Netherlands. Leukemia 2004, 18, 2032–2035. [Google Scholar] [CrossRef]
- Ramanujachar, R.; Richards, S.; Hann, I.; Goldstone, A.; Mitchell, C.; Vora, A.; Rowe, J.; Webb, D. Adolescents with acute lymphoblastic leukaemia: Outcome on UK national paediatric (ALL97) and adult (UKALLXII/E2993) trials. Pediatr. Blood Cancer 2017, 48, 254–261. [Google Scholar] [CrossRef]
- Boissel, N.; Sender, L.S. Best Practices in Adolescent and Young Adult Patients with Acute Lymphoblastic Leukemia: A Focus on Asparaginase. J. Adolesc. Young Adult Oncol. 2015, 4, 118–128. [Google Scholar] [CrossRef] [Green Version]
- Salzer, W.; Bostrom, B.; Messinger, Y.; Perissinotti, A.J.; Marini, B. Asparaginase activity levels and monitoring in patients with acute lymphoblastic leukemia. Leuk. Lymphoma 2018, 59, 1797–1806. [Google Scholar] [CrossRef]
- Avramis, V.I.; Tiwari, P.N. Asparaginase (native ASNase or pegylated ASNase) in the treatment of acute lymphoblastic leukemia. Int. J. Nanomed. 2006, 1, 241–254. [Google Scholar]
- Pieters, R.; Hunger, S.P.; Boos, J.; Rizzari, C.; Silverman, L.; Baruchel, A.; Goekbuget, N.; Schrappe, M.; Pui, C.H. L-asparaginase treatment in acute lymphoblastic leukemia: A focus on Erwinia asparaginase. Cancer 2011, 117, 238–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.Q.; Tao, M.L.; Shen, W.D.; Zhou, Y.Z.; Ding, Y.; Ma, Y.; Zhou, W.L. Immobilization of L-asparaginase on the microparticles of the natural silk sericin protein and its characters. Biomaterials 2004, 17, 3751–3759. [Google Scholar] [CrossRef]
- Brumano, L.P.; da Silva, F.V.S.; Costa-Silva, T.A.; Apolinário, A.C.; Santos, J.H.P.M.; Kleingesinds, E.K.; Monteiro, G.; Rangel-Yagui, C.O.; Benyahia, B.; Junior, A.P. Development of L-Asparaginase Biobetters: Current Research Status and Review of the Desirable Quality Profiles. Front. Bioeng. Biotechnol. 2019, 6, 212. [Google Scholar] [CrossRef] [Green Version]
- Sassi, A.B.; Nagarkar, R.; Hamblin, P. Chapter 9—Biobetter Biologics. In Novel Approaches and Strategies for Biologics, Vaccines and Cancer Therapies; Singh, M., Salnikova, M., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 199–217. ISBN 9780124166035. [Google Scholar] [CrossRef]
- Verma, S.; Jain, P.; Chauhan, P. Biobetters: The better biologics and their regulatory overview. Int. J. Drug Regul. Aff. 2018, 4, 13–20. [Google Scholar] [CrossRef]
- Juran, J.M. Juran on Quality by Design: The New Steps for Planning Quality into Goods and Services; The Free Press: New York, NY, USA, 1992. [Google Scholar]
- Grangeia, H.B.; Silva, C.; Simões, S.P.; Reis, M.S. Quality by design in pharmaceutical manufacturing: A systematic review of current status, challenges and future perspectives. Eur. J. Pharm. Biopharm. 2020, 147, 19–37. [Google Scholar] [CrossRef] [PubMed]
- Keating, G.M. Asparaginase Erwinia chrysanthemi (Erwinaze®): A guide to its use in acute lymphoblastic leukemia in the USA. BioDrugs 2013, 27, 413–418. [Google Scholar] [CrossRef]
- Heo, Y.A.; Syed, Y.Y.; Keam, S.J. Pegaspargase: A Review in Acute Lymphoblastic Leukaemia. Drugs 2019, 79, 767–777. [Google Scholar] [CrossRef] [Green Version]
- Lopes, A.M.; Oliveira-Nascimento, L.; Ribeiro, A.; Tairum, C.A., Jr.; Breyer, C.A.; Oliveira, M.A.; Monteiro, G.; Souza-Motta, C.M.; Magalhães, P.O.; Avendaño, J.G.; et al. Therapeutic l-asparaginase: Upstream, downstream and beyond. Crit. Rev. Biotechnol. 2017, 37, 82–99. [Google Scholar] [CrossRef] [Green Version]
- Ulu, A.; Ates, B. Immobilization of l-Asparaginase on Carrier Materials: A Comprehensive Review. Bioconjug. Chem. 2017, 28, 1598–1610. [Google Scholar] [CrossRef]
- Beckett, A.; Gervais, D. What makes a good new therapeutic L-asparaginase? World J. Microbiol. Biotechnol. 2019, 35, 152. [Google Scholar] [CrossRef]
- Huerta-Saquero, A.; Evangelista-Martínez, Z.; Moreno-Enriquez, A.; Perez-Rueda, E. Rhizobium etli asparaginase II: An alternative for acute lymphoblastic leukemia (ALL) treatment. Bioengineered 2013, 4, 30–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qeshmi, I.F.; Homaei, A.; Fernandes, P.; Javadpour, S. Marine microbial L-asparaginase: Biochemistry, molecular approaches and applications in tumor therapy and in food industry. Microbiol. Res. 2018, 208, 99–112. [Google Scholar] [CrossRef]
- Souza, P.M.; de Freitas, M.M.; Cardoso, S.L.; Pessoa, A.; Guerra, E.N.S.; Magalhães, P.O. Optimization and purification of l-asparaginase from fungi: A systematic review. Crit. Rev. Oncol. Hematol. 2017, 120, 194–202. [Google Scholar] [CrossRef]
- Muneer, F.; Hussnain, M.S.; Azeem, F.; Rasul, I.; Muzammil, S.; Zubair, M.; Afzal, M.; Nadeem, H. Microbial L-asparaginase: Purification, characterization and applications. Arch. Microbiol. 2020, 202, 967–981. [Google Scholar] [CrossRef] [PubMed]
- Rathod, S.; Ramsey, M.; Relling, M.V.; Finkelman, F.D.; Fernandez, C.A. Hypersensitivity reactions to asparaginase in mice are mediated by anti-asparaginase IgE and IgG and the immunoglobulin receptors FcεRI and FcγRIII. Haematologica 2019, 104, 319–329. [Google Scholar] [CrossRef]
- Nordengrün, M.; Michalik, S.; Völker, U.; Bröker, B.M.; Gómez-Gascón, L. The quest for bacterial allergens. Int. J. Med. Microbiol. 2018, 308, 738–750. [Google Scholar] [CrossRef] [PubMed]
- Cruz, M.E.M.; Gaspar, M.M.; Lopes, F.; Jorge, J.S.; Perez-Soler, R. Liposomal l-asparaginase: In vitro evaluation. Int. J. Pharm. 1993, 96, 67–77. [Google Scholar] [CrossRef]
- Jorge, J.C.S.; Perez-Soler, R.; Morais, J.G.; Cruz, M.E.M. Liposomal palmitoyl-L-asparaginase: Characterization and biological activity. Cancer Chemother. Pharmacol. 1994, 34, 230–234. [Google Scholar] [CrossRef]
- Gaspar, M.M.; Perez-Soler, R.; Cruz, M.E.M. Biological characterization of L-asparaginase liposomal formulations. Cancer Chemother. Pharmacol. 1996, 38, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, M.M.; Blanco, D.; Cruz, M.E.M.; Alonso, M.J. Formulation of L-asparaginase-loaded poly(lactide-co-glycolide) nanoparticles: Influence of polymer properties on enzyme loading, activity and in vitro release. J. Cntrl. Rel. 1998, 52, 53–62. [Google Scholar] [CrossRef]
- Baran, E.T.; Özer, N.; Hasirci, V. In vivo half-life of nanoencapsulated L-asparaginase. J. Mater. Sci.: Mater. Med. 2002, 13, 1113–1121. [Google Scholar] [CrossRef]
- Bahreini, E.; Aghaiypour, K.; Abbasalipourkabir, R.; Goodarzi, M.T.; Saidijam, M. Preparation and nanoencapsulation of l-asparaginase II in chitosan-tripolyphosphate nanoparticles and in vitro release study. Nanoscale Res. Lett. 2014, 9, 340. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.Q.; Wang, y.; Wang, H.Y.; Zhu, L.; Zhou, Z.Z. Highly efficient processing of silk fibroin nanoparticle-l-asparaginase bioconjugates and their characterization as a drug delivery system. Soft Matter. 2011, 7, 9728–9736. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Zhang, P.Y. Polymersomes in Nanomedicine—A Review. Curr. Med. Chem. 2017, 13, 124–129. [Google Scholar] [CrossRef]
- Tinoco, A.; Ribeiro, A.; Oliveira, C.; Parpot, P.; Gomes, A.; Cavaco-Paulo, A. Albumin/asparaginase capsules prepared by ultrasound to retain ammonia. Appl. Microbiol. Biotechnol. 2016, 100, 9499–9508. [Google Scholar] [CrossRef] [Green Version]
- Bueno, C.Z.; Apolinário, A.C.; Duro-Castano, A.; Poma, A.; Pessoa, A., Jr.; Rangel-Yagui, C.O.; Battaglia, G. L-Asparaginase Encapsulation into Asymmetric Permeable Polymersomes. ACS Macro. Lett. 2020, 9, 1471–1477. [Google Scholar] [CrossRef]
- Mu, X.; Qiao, J.; Qi, L.; Dong, P.; Ma, H. Poly(2-vinyl-4,4-dimethylazlactone)-functionalized magnetic nanoparticles as carriers for enzyme immobilization and its application. ACS Appl. Mater. Interfaces 2014, 6, 21346–21354. [Google Scholar] [CrossRef] [PubMed]
- Sueyoshi, D.; Anraku, Y.; Komatsu, T.; Urano, Y.; Kataoka, K. Enzyme-Loaded Polyion Complex Vesicles as in Vivo Nanoreactors Working Sustainably under the Blood Circulation: Characterization and Functional Evaluation. Biomacromolecules 2017, 18, 1189–1196. [Google Scholar] [CrossRef]
- Alpar, H.O.; Lewis, D.A. Therapeutic efficacy of asparaginase encapsulated in intact erythrocytes. Biochem. Pharmacol. 1985, 34, 257–261. [Google Scholar] [CrossRef]
- Ha, W.; Meng, X.W.; Li, Q.; Fan, M.M.; Peng, S.L.; Ding, L.S.; Tian, X.; Zhang, S.; Li, B.J. Self-assembly hollow nanosphere for enzyme encapsulation. Soft Mat. 2010, 6, 1405–1408. [Google Scholar] [CrossRef]
- Díaz-Barriga, C.; Villanueva-Flores, F.; Quester, K.; Zárate-Romero, A.; Cadena-Nava, R.D.; Huerta-Saquero, A. Asparaginase-Phage P22 Nanoreactors: Toward a Biobetter Development for Acute Lymphoblastic Leukemia Treatment. Pharmaceutics 2021, 13, 604. [Google Scholar] [CrossRef]
- Patil, Y.P.; Jadhav, S. Novel methods for liposome preparation. Chem. Phys. Lipids 2014, 177, 8–18. [Google Scholar] [CrossRef]
- Sessa, G.; Weissmann, G. Incorporation of lysozyme into liposomes. A model for structure-linked latency. J. Biol. Chem. 1970, 245, 3295–3301. [Google Scholar] [CrossRef]
- Has, C.; Sunthar, P. A comprehensive review on recent preparation techniques of liposomes. J. Liposome Res. 2020, 30, 336–365. [Google Scholar] [CrossRef]
- Chang, H.I.; Yeh, M.K. Clinical development of liposome-based drugs: Formulation, characterization, and therapeutic efficacy. Int. J. Nanomedicine 2012, 7, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Mozafari, M.R.; Mortazavi, S.M. Nanoliposomes: From Fundamentals to Recent Developments; Trafford Publishing Ltd: Oxford, UK, 2005; ISBN-10: 1412055458. [Google Scholar]
- Zhu, M.; Cheng, Y.; Luo, Q.; El-khateeb, M. A review of synthetic approaches to hollow nanostructures. Mater. Chem. Front. 2021, 5, 2552–2587. [Google Scholar] [CrossRef]
- Jahadi, M.; Khosravi-Darani, K. Liposomal Encapsulation Enzymes: From Medical Applications to Kinetic Characteristics. Mini-Rev. Med. Chem. 2017, 17, 366–370. [Google Scholar] [CrossRef]
- Do, T.T.; Do, T.P.; Nguyen, T.N.; Nguyen, T.C.; Vu, T.T.P.; Nguyen, T.G.A. Nanoliposomal L-Asparaginase and Its Antitumor Activities in Lewis Lung Carcinoma Tumor-Induced BALB/c Mice. Adv. Mater. Sci. Eng. 2019, 3534807. [Google Scholar] [CrossRef] [Green Version]
- Giorno, L.; Drioli, E. Biocatalytic membrane reactors: Applications and perspectives. Trends Biotechnol. 2000, 18, 339–349. [Google Scholar] [CrossRef]
- Castner, J.F.; Wingard, L.B., Jr. Mass transport and reaction kinetic parameters determined electrochemically for immobilized glucose oxidase. Biochemistry 1984, 23, 2203–2210. [Google Scholar] [CrossRef] [PubMed]
- Bartolini, M.; Andrisano, V.; Wainer, I.W. Development and characterization of an immobilized enzyme reactor based on glyceraldehyde-3-phosphate dehydrogenase for on-line enzymatic studies. J. Chromatogr. A 2003, 987, 331–340. [Google Scholar] [CrossRef]
- Sharma, A.; Sharma, U.S. Liposomes in drug delivery: Progress and limitations. Int. J. Pharm. 1997, 154, 123–140. [Google Scholar] [CrossRef]
- Allen, T.M. The use of glycolipids and hydrophilic polymers in avoiding rapid uptake of liposomes by the mononuclear phagocyte system. Adv. Drug Deliv. Rev. 1994, 13, 285–309. [Google Scholar] [CrossRef]
- Corrêa, A.C.N.T.F.; Vericimo, M.A.; Dashevskiy, A.; Pereira, P.R.; Paschoalin, V.M.F. Liposomal Taro Lectin Nanocapsules Control Human Glioblastoma and Mammary Adenocarcinoma Cell Proliferation. Molecules 2019, 24, 471. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Abetxuko, A.; Sánchez-deAlcázar, D.; Muñumer, P.; Beloqui, A. Tunable Polymeric Scaffolds for Enzyme Immobilization. Front. Bioeng. Biotechnol. 2020, 8, 830. [Google Scholar] [CrossRef]
- Rivera-Briso, A.L.; Serrano-Aroca, Á. Poly(3-Hydroxybutyrate-co-3-Hydroxyvalerate): Enhancement Strategies for Advanced Applications. Polymers 2018, 10, 732. [Google Scholar] [CrossRef] [Green Version]
- Kasaai, M.R. Molecular Weight Distribution for Biopolymers: A Review. J. Polym. Biopolym. Phys. Chem. 2018, 6, 39–44. [Google Scholar] [CrossRef]
- Einfalt, T.; Witzigmann, D.; Edlinger, C.; Sieber, S.; Goers, R.; Najer, A.; Spulber, M.; Onaca-Fischer, O.; Huwyler, J.; Palivan, C.G. Biomimetic artificial organelles with in vitro and in vivo activity triggered by reduction in microenvironment. Nat. Commun. 2018, 9, 1127. [Google Scholar] [CrossRef] [Green Version]
- Blackman, L.D.; Varlas, S.; Arno, M.C.; Houston, Z.H.; Fletcher, N.L.; Thurecht, K.J.; Hasan, M.; Gibson, M.I.; O’Reilly, R.K. Confinement of Therapeutic Enzymes in Selectively Permeable Polymer Vesicles by Polymerization-Induced Self-Assembly (PISA) Reduces Antibody Binding and Proteolytic Susceptibility. ACS Cent. Sci. 2018, 4, 718–723. [Google Scholar] [CrossRef] [PubMed]
- Rideau, E.; Dimova, R.; Schwille, P.; Wurm, F.R.; Landfester, K. Liposomes and polymersomes: A comparative review towards cell mimicking. Chem. Soc. Rev. 2018, 47, 8572–8610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apolinário, A.C.; Ferraro, R.B.; de Oliveira, C.A.; Pessoa, A., Jr.; de Oliveira Rangel-Yagui, C. Quality-by-Design Approach for Biological API Encapsulation into Polymersomes Using “Off-the-Shelf” Materials: A Study on L-Asparaginase. AAPS PharmSciTech 2019, 20, 251. [Google Scholar] [CrossRef]
- Wu, K.; Su, D.; Liu, J.; Saha, R.; Wang, J.P. Magnetic nanoparticles in nanomedicine: A review of recent advances. Nanotechnology 2019, 30, 502003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samrot, A.V.; Sahithya, C.S.; Selvarani, J.A.; Purayil, S.K.; Ponnaiah, P. A review on synthesis, characterization and potential biological applications of superparamagnetic iron oxide nanoparticles. Curr. Res. Green Sustain. Chem. 2021, 4, 100042. [Google Scholar] [CrossRef]
- Alam, S.; Ahmad, R.; Pranaw, K.; Mishra, P.; Khare, S.K. Asparaginase conjugated magnetic nanoparticles used for reducing acrylamide formation in food model system. Bioresour. Technol. 2018, 269, 121–126. [Google Scholar] [CrossRef]
- Teodor, E.; Litescu, S.C.; Lazar, V.; Somoghi, R. Hydrogel-magnetic nanoparticles with immobilized L-asparaginase for biomedical applications. J. Mater. Sci. Mater. Med. 2009, 20, 1307–1314. [Google Scholar] [CrossRef]
- Baskar, G.; George, G.B. Glutaraldehyde-Mediated Synthesis of Asparaginase-Bound Maghemite Nanocomposites: Cytotoxicity against Human Colon Adenocarcinoma Cells. Asian Pac. J. Cancer Prev. 2016, 17, 4237–4240. [Google Scholar] [PubMed]
- Orhan, H.; Uygun, D.A. Immobilization of L-Asparaginase on Magnetic Nanoparticles for Cancer Treatment. Appl. Biochem. Biotechnol. 2020, 191, 1432–1443. [Google Scholar] [CrossRef]
- Ulu, A.; Noma, S.A.A.; Koytepe, S.; Ates, B. Magnetic Fe3O4@MCM-41 core-shell nanoparticles functionalized with thiol silane for efficient l-asparaginase immobilization. Artif. Cells Nanomed. Biotechnol. 2018, 46 (Suppl. 2), 1035–1045. [Google Scholar] [CrossRef] [Green Version]
- Feng, Q.; Liu, Y.; Huang, J.; Chen, K.; Huang, J.; Xiao, K. Uptake, distribution, clearance, and toxicity of iron oxide nanoparticles with different sizes and coatings. Sci. Rep. 2018, 8, 2082. [Google Scholar] [CrossRef]
- Malhotra, N.; Lee, J.S.; Liman, R.A.D.; Ruallo, J.M.S.; Villaflores, O.B.; Ger, T.R.; Hsiao, C.D. Potential Toxicity of Iron Oxide Magnetic Nanoparticles: A Review. Molecules 2020, 25, 3159. [Google Scholar] [CrossRef]
- Fernández-Bertólez, N.; Costa, C.; Bessa, M.J.; Park, M.; Carriere, M.; Dussert, F.; Teixeira, J.P.; Pásaro, E.; Laffon, B.; Valdiglesias, V. Assessment of oxidative damage induced by iron oxide nanoparticles on different nervous system cells. Mutat. Res. 2019, 845, 402989. [Google Scholar] [CrossRef]
- Kwolek, U.; Nakai, K.; Pluta, A.; Zatorska, M.; Wnuk, D.; Lasota, S.; Bednar, J.; Michalik, M.; Yusa, S.I.; Kepczynski, M. Polyion complex vesicles (PICsomes) from strong copolyelectrolytes. Stability and in vitro studies. Colloids Surf B Biointerfaces 2017, 158, 658–666. [Google Scholar] [CrossRef]
- Kurinomaru, T.; Shiraki, K. Noncovalent PEGylation of L-asparaginase using PEGylated polyelectrolyte. J. Pharm. Sci. 2015, 104, 587–592. [Google Scholar] [CrossRef] [Green Version]
- Tong, W.H.; Pieters, R.; Tissing, W.J.; van der Sluis, I.M. Desensitization protocol should not be used in acute lymphoblastic leukemia patients with silent inactivation of PEGasparaginase. Haematologica 2014, 99, e102–e104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grollmisch, A.; Kragl, U.; Großeheilmann, J. Enzyme Immobilization in Polymerized Ionic Liquids-based Hydrogels for Active and Reusable Biocatalysts. Synopen 2018, 2, 192–199. [Google Scholar] [CrossRef] [Green Version]
- Insua, I.; Wilkinson, A.; Fernandez-Trillo, F. Polyion complex (PIC) particles: Preparation and biomedical applications. Eur. Polym. J. 2016, 81, 198–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; He, Z.; Wang, H.; Qi, L.U.; Liu, G.; Zhang, X. Applications of hollow nanomaterials in environmental remediation and monitoring: A review. Front. Environ. Sci. Eng. 2015, 9, 770–783. [Google Scholar] [CrossRef]
- Yan, Z.J.; Li, W.Y.; Hu, X.Y.; Deng, X.; Wang, S.L.; Zhang, J.Q. Pharmacokinetics and Bioequiavailability of Asparaginase in Asparaginase Nanospheres. J. Sichuan Univ. Med Sci. Ed. 2016, 47, 411–415. [Google Scholar]
- Chauhan, K.; Hernandez-Meza, J.M.; Rodríguez-Hernández, A.G.; Juarez-Moreno, K.; Sengar, P.; Vazquez-Duhalt, R. Multifunctionalized biocatalytic P22 nanoreactor for combinatory treatment of ER+ breast cancer. J. Nanobiotechnol. 2018, 16, 17. [Google Scholar] [CrossRef]
- Hill, B.D.; Zak, A.; Khera, E.; Wen, F. Engineering Virus-like Particles for Antigen and Drug Delivery. Curr. Protein Pept. Sci. 2018, 19, 112–127. [Google Scholar] [CrossRef]
- Nuñez-Rivera, A.; Fournier, P.G.J.; Arellano, D.L.; Rodriguez-Hernandez, A.G.; Vazquez-Duhalt, R.; Cadena-Nava, R.D. Brome mosaic virus-like particles as siRNA nanocarriers for biomedical purposes. Beilstein J. Nanotechnol. 2020, 11, 372–382. [Google Scholar] [CrossRef] [Green Version]
- Ihler, G.M.; Glew, R.H.; Schnure, F.W. Enzyme loading of erythrocytes. Proc. Natl. Acad. Sci. USA 1973, 70, 2663–2666. [Google Scholar] [CrossRef] [PubMed]
- Thomas, X.; Le Jeune, C. Erythrocyte encapsulated l-asparaginase (GRASPA) in acute leukemia. Int. J. Hematol. Oncol. 2016, 5, 11–25. [Google Scholar] [CrossRef]
- Bax, B.E. Erythrocytes as Carriers of Therapeutic Enzymes. Pharmaceutics 2020, 12, 435. [Google Scholar] [CrossRef] [PubMed]
- Domenech, C.; Thomas, X.; Chabaud, S.; Baruchel, A.; Gueyffier, F.; Mazingue, F.; Auvrignon, A.; Corm, S.; Dombret, H.; Chevallier, P.; et al. l-asparaginase loaded red blood cells in refractory or relapsing acute lymphoblastic leukaemia in children and adults: Results of the GRASPALL 2005-01 randomized trial. Br. J. Haematol. 2011, 153, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Hammel, P.; Fabienne, P.; Mineur, L.; Metges, J.P.; Andre, T.; De La Fouchardiere, C.; Louvet, C.; El Hajbi, F.; Faroux, R.; Guimbaud, R.; et al. Erythrocyte-encapsulated asparaginase (eryaspase) combined with chemotherapy in second-line treatment of advanced pancreatic cancer: An open-label, randomized Phase IIb trial. Eur. J. Cancer 2020, 124, 91–101. [Google Scholar] [CrossRef] [Green Version]
- Rossi, L.; Pierigè, F.; Aliano, M.P.; Magnani, M. Ongoing Developments and Clinical Progress in Drug-Loaded Red Blood Cell Technologies. BioDrugs 2020, 34, 265–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, Y.M.; Chung, H.S.; Moon, C.; Yockman, J.; Park, Y.J.; Gitlin, S.D.; David, A.E.; Yang, V.C. L-Asparaginase encapsulated intact erythrocytes for treatment of acute lymphoblastic leukemia (ALL). J. Control. Release 2009, 139, 182–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carriers for Asnase Encapsulation | Advantages | Disadvantages |
|---|---|---|
| LIPOSOME | Biodegradable. Approved for clinical trials. Easy synthesis. Low-cost production. Widely studied. | Low stability, leakiness, and low blood circulation times when are not functionalized. Fast elimination from the blood. |
| BIOPOLYMER | Biodegradable. Can be obtained at low cost from agricultural waste or through biotechnology, from bacterial cultures. Non-antigenic molecules. | Poor mechanical properties. Heterogeneous molecular weights could be a drawback that affects entrapment efficiency, stability, and enzyme kinetic parameters at the large-scale manufacturing. |
| SILK NANOPARTICLE | Biodegradable. Non-antigenic molecules. Low-cost production. | Heterogeneous molecular weights. |
| POLYMEROSOME | Can range from nm up to μm. Their membrane thickness is bigger than liposomes. More stable and permeable than unconjugated liposomes. They can be designed as stimuli-responsive carriers. Polymerosomes production is easily scaled up. | Non-biodegradable. Lack of enough information about in vivo cytotoxicity. |
| MAGNETIC NANOPARTICLE | Easy separation, and fate-direction under an external magnetic field. | Non-biodegradable. Some magnetic nanoparticles induced inflammation, ulceration, and a decrement of the cell growth rate, a decline in viability and triggering of neurobehavioral alterations in both cell lines and animal models. |
| POLYION VESICLE | Easy, non-expensive and scalable production. These formulations combined the advantages of ionic liquids (ionic conductivity, thermal, and chemical stability, tunable solution properties), and the properties of polymers. | Not always biodegradable. Poorly studied. |
| HOLLOW NANOSPHERE | Non immunogenic. Scalable process. | Poorly studied. |
| VIRUS-LIKE PARTICLE | Biodegradable. Scalable and low-cost production. High stability. | Immunogenic when are not functionalized. Poorly studied. |
| RED BLOOD CELL | Biodegradable, approved for clinical trials. Long blood circulating times. Non immunogenic. Widely studied. | Low rate of red blood cell survival after enzyme internalization. Its production requires permission for human samples managing. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villanueva-Flores, F.; Zárate-Romero, A.; Torres, A.G.; Huerta-Saquero, A. Encapsulation of Asparaginase as a Promising Strategy to Improve In Vivo Drug Performance. Pharmaceutics 2021, 13, 1965. https://doi.org/10.3390/pharmaceutics13111965
Villanueva-Flores F, Zárate-Romero A, Torres AG, Huerta-Saquero A. Encapsulation of Asparaginase as a Promising Strategy to Improve In Vivo Drug Performance. Pharmaceutics. 2021; 13(11):1965. https://doi.org/10.3390/pharmaceutics13111965
Chicago/Turabian StyleVillanueva-Flores, Francisca, Andrés Zárate-Romero, Alfredo G. Torres, and Alejandro Huerta-Saquero. 2021. "Encapsulation of Asparaginase as a Promising Strategy to Improve In Vivo Drug Performance" Pharmaceutics 13, no. 11: 1965. https://doi.org/10.3390/pharmaceutics13111965
APA StyleVillanueva-Flores, F., Zárate-Romero, A., Torres, A. G., & Huerta-Saquero, A. (2021). Encapsulation of Asparaginase as a Promising Strategy to Improve In Vivo Drug Performance. Pharmaceutics, 13(11), 1965. https://doi.org/10.3390/pharmaceutics13111965