Increased Efficacy of Oral Fixed-Dose Combination of Amphotericin B and AHCC® Natural Adjuvant against Aspergillosis

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
2.3. Inoculum
2.4. Murine Model of Pulmonary Aspergillosis
2.5. Treatment
2.6. Fungal Burden Studies
2.7. Sera Determinations
2.8. Statistical Analysis
3. Results
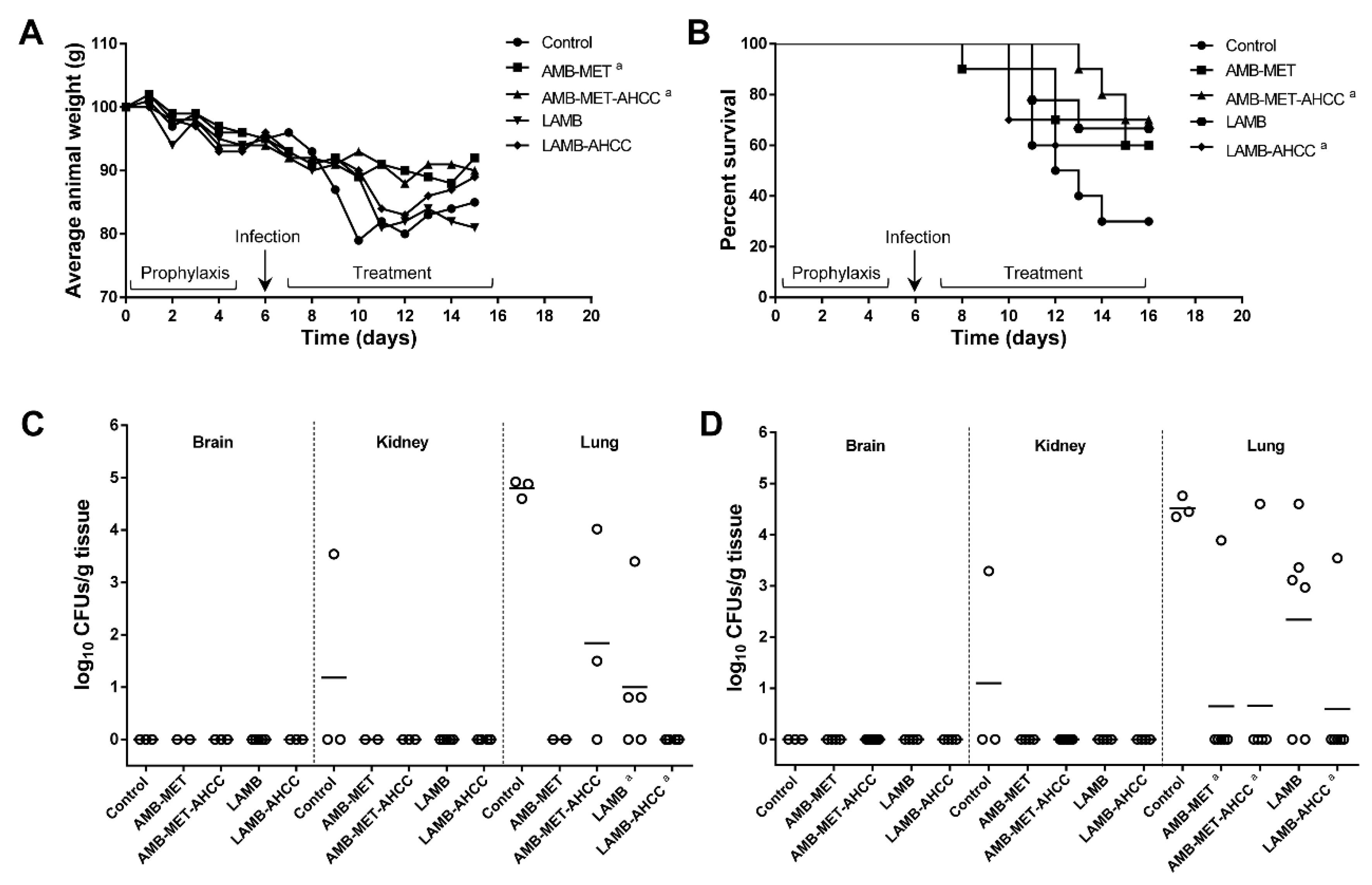
3.1. Survival and Body Weight Progression
3.2. Fungal Burden
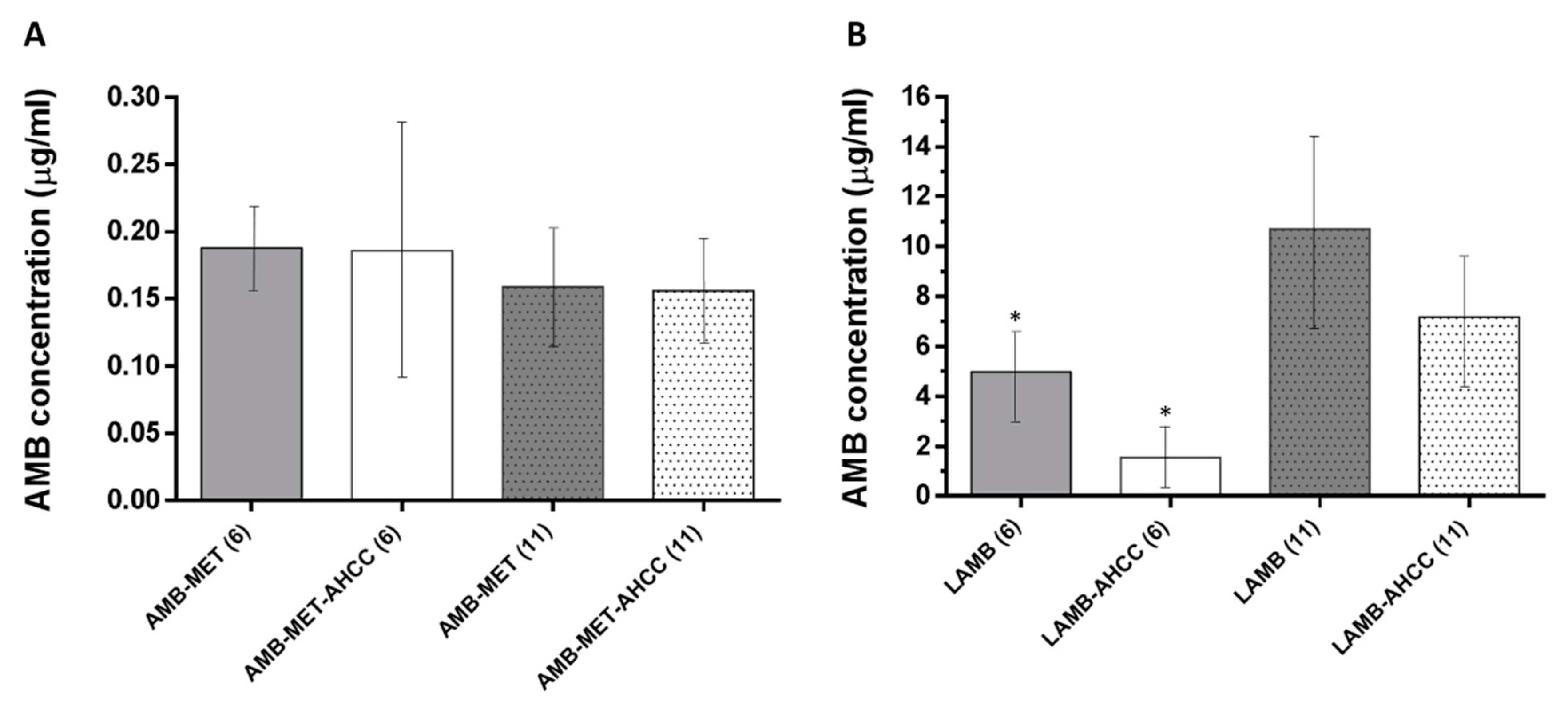
3.3. AMB Concentration in Plasma
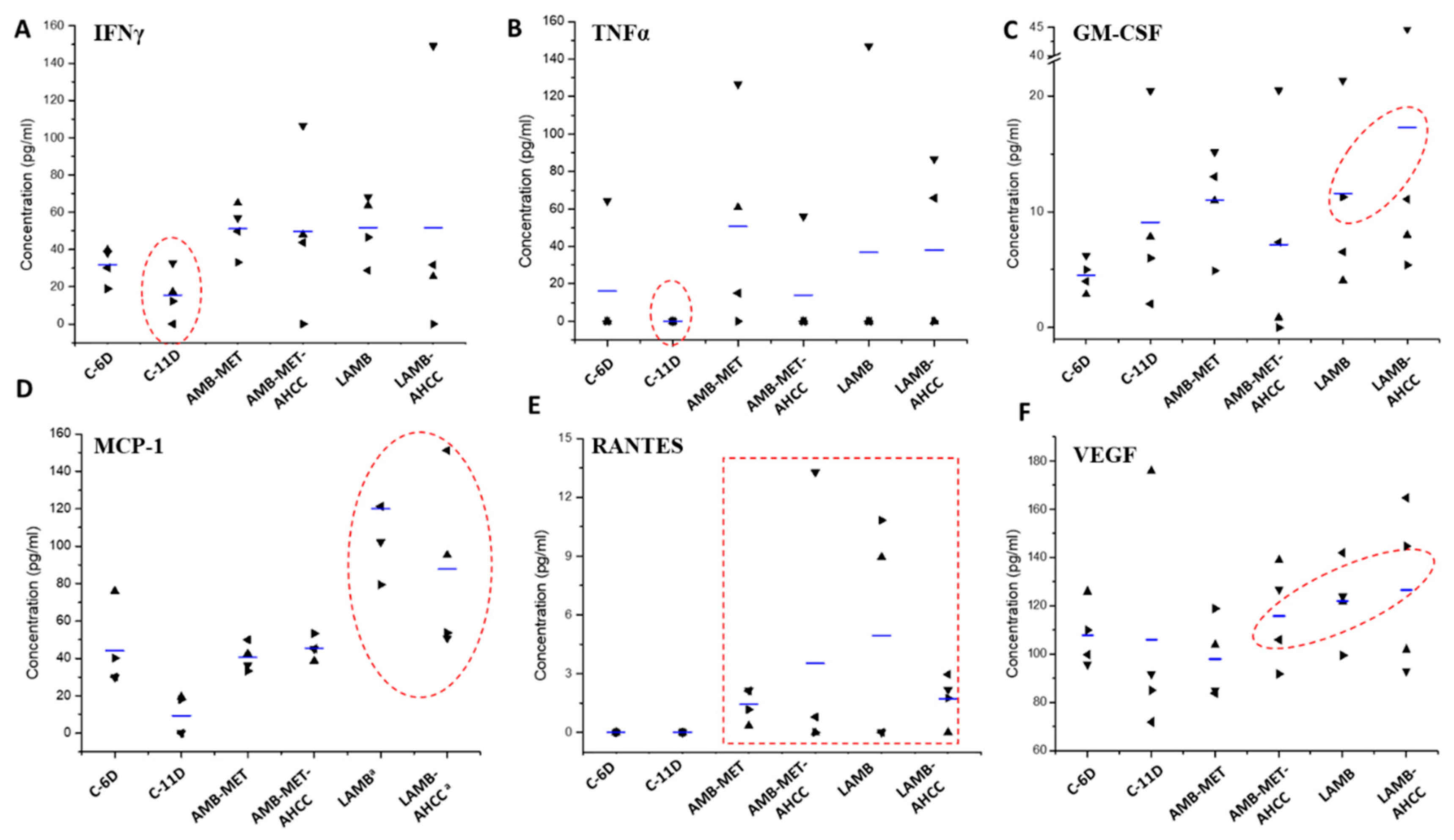
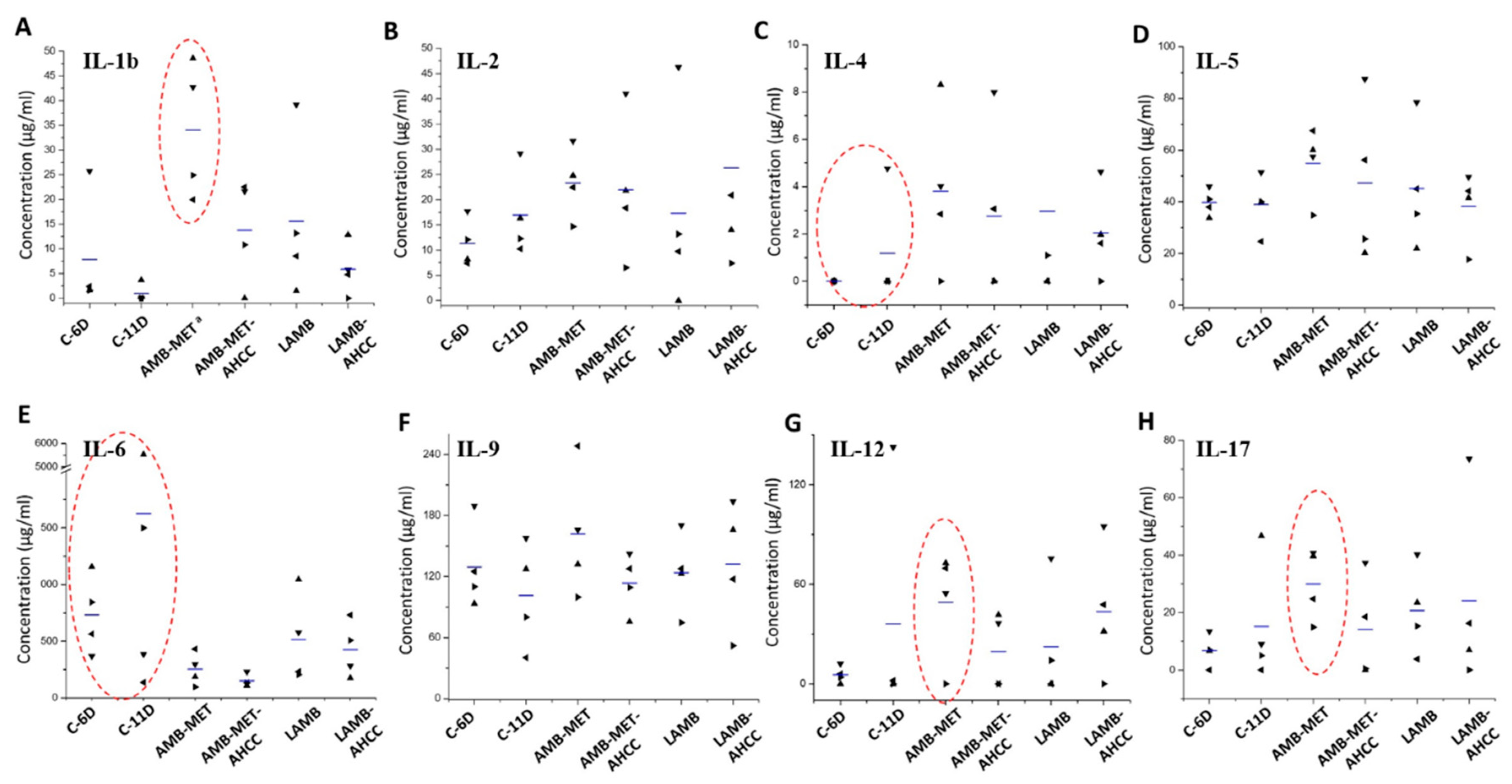
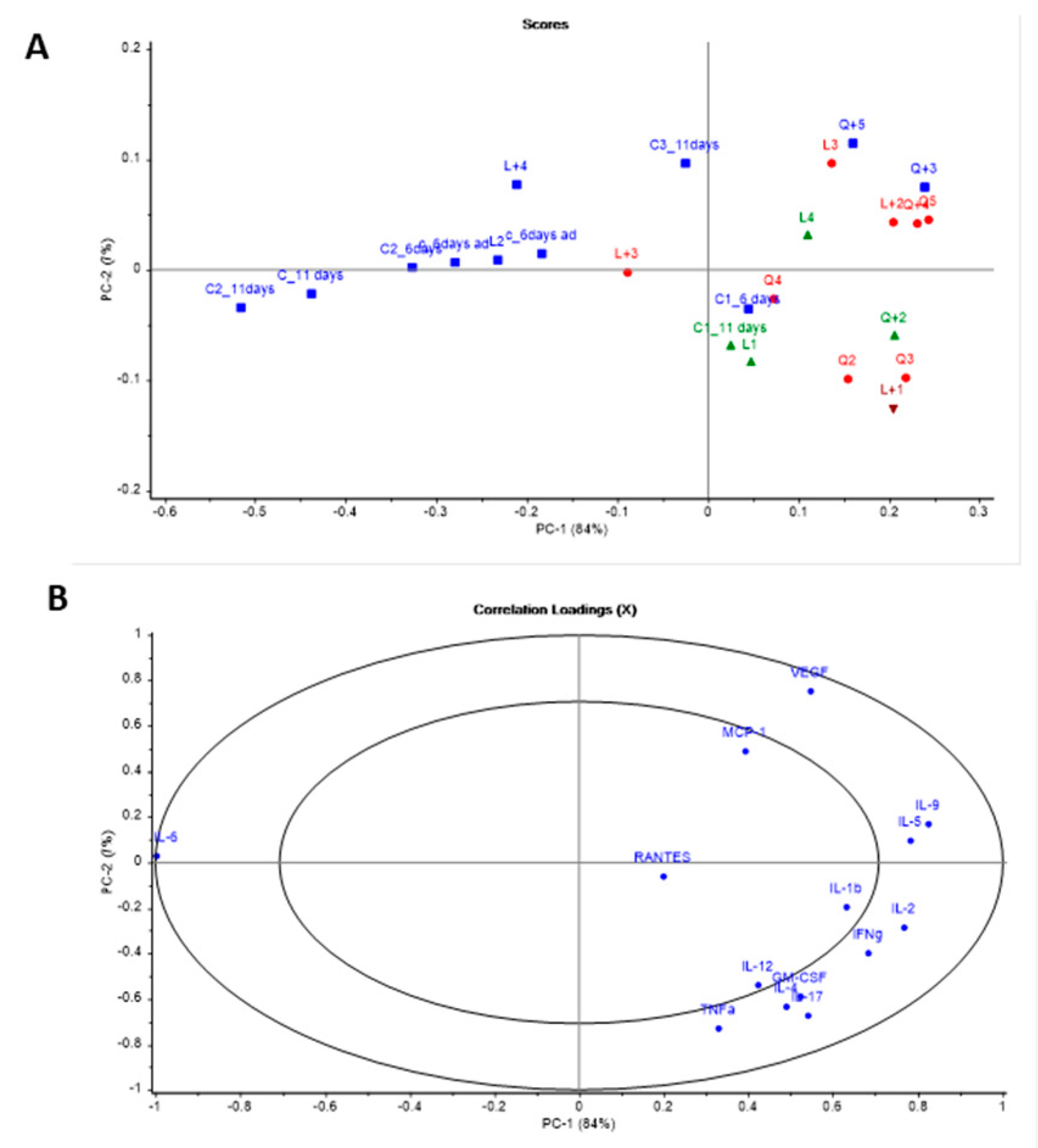
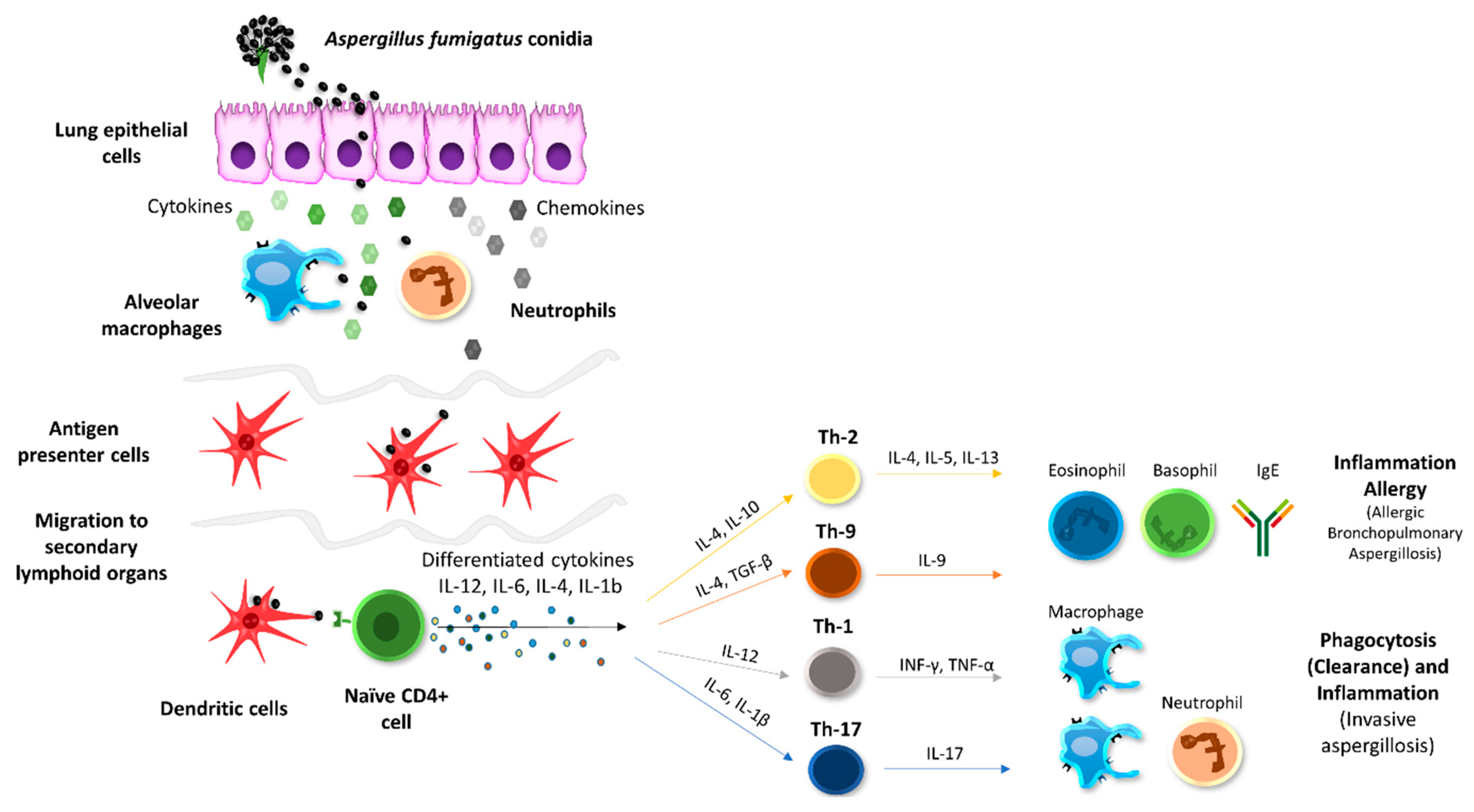
3.4. Cytokine Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chowdhary, A.; Sharma, C.; Meis, J.F. Azole-Resistant Aspergillosis: Epidemiology, Molecular Mechanisms, and Treatment. J. Infect. Dis. 2017, 216, S436–S444. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, S.S. Global Aspects of Triazole Resistance in Aspergillus fumigatus with Focus on Latin American Countries. J. Fungi 2017, 3, 5. [Google Scholar] [CrossRef]
- Brown, G.D.; Denning, D.W.; Gow, N.A.R.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden killers: Human fungal infections. Sci. Transl. Med. 2012, 4, 165rv13. [Google Scholar] [CrossRef] [PubMed]
- Lamoth, F. Aspergillus fumigatus-Related Species in Clinical Practice. Front. Microbiol. 2016, 7, 683. [Google Scholar] [CrossRef]
- Patterson, T.F.; Thompson, G.R.; Denning, D.W.; Fishman, J.A.; Hadley, S.; Herbrecht, R.; Kontoyiannis, D.P.; Marr, K.A.; Morrison, V.A.; Nguyen, M.H.; et al. Practice guidelines for the diagnosis and management of aspergillosis: 2016 update by the infectious diseases society of America. Clin. Infect. Dis. 2016, 63, e1–e60. [Google Scholar] [CrossRef] [PubMed]
- Wiederhold, N.; Patterson, T. Emergence of Azole Resistance in Aspergillus. Semin. Respir. Crit. Care Med. 2015, 36, 673–680. [Google Scholar] [CrossRef]
- Rivero-Menendez, O.; Alastruey-Izquierdo, A.; Mellado, E.; Cuenca-Estrella, M. Triazole Resistance in Aspergillus spp.: A Worldwide Problem? J. Fungi 2016, 2, 21. [Google Scholar] [CrossRef]
- Aruanno, M.; Glampedakis, E.; Lamoth, F. Echinocandins for the treatment of invasive aspergillosis: From laboratory to bedside. Antimicrob. Agents Chemother. 2019, 63, e00399-19. [Google Scholar] [CrossRef]
- Martin-Vicente, A.; Capilla, J.; Guarro, J. In Vivo Synergy of Amphotericin B plus Posaconazole in Murine Aspergillosis. Antimicrob. Agents Chemother. 2016, 60, 296–300. [Google Scholar] [CrossRef]
- Bedin, D.L.; de Jesus, F.P.K.; Keller, J.T.; Weiblen, C.; de Azevedo, M.I.; Oliveira, V.; Morais, S.J.; Hartz, A.S. Evaluation of the efficacy of a posaconazole and anidulafungin combination in a murine model of pulmonary aspergillosis due to infection with Aspergillus fumigatus. Diagn. Microbiol. Infect. Dis. 2018, 90, 40–43. [Google Scholar] [CrossRef]
- Scorzoni, L.; de Paula e Silva, A.C.A.; Marcos, C.M.; Assato, P.A.; de Melo, W.C.M.A.; de Oliveira, H.C.; Costa-Orlandi, C.B.; Mendes-Giannini, M.J.; Fusco-Almeida, A.M. Antifungal Therapy: New Advances in the Understanding and Treatment of Mycosis. Front. Microbiol. 2017, 8, 36. [Google Scholar] [CrossRef] [PubMed]
- Steimbach, L.M.; Tonin, F.S.; Virtuoso, S.; Borba, H.H.L.; Sanches, A.C.C.; Wiens, A.; Fernandez-Llimós, F.; Pontarolo, R. Efficacy and safety of amphotericin B lipid-based formulations—A systematic review and meta-analysis. Mycoses 2017, 60, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Falagas, M.E.; Karageorgopoulos, D.E.; Tansarli, G.S. Continuous versus Conventional Infusion of Amphotericin B Deoxycholate: A Meta-Analysis. PLoS ONE 2013, 8, e77075. [Google Scholar] [CrossRef] [PubMed]
- Falci, D.R.; da Rosa, F.B.; Pasqualotto, A.C. Comparison of nephrotoxicity associated to different lipid formulations of amphotericin B: A real-life study. Mycoses 2015, 58, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Serrano, D.R.; Lalatsa, A.; Dea-Ayuela, M.A.; Bilbao-Ramos, P.E.; Garrett, N.L.; Moger, J.; Guarro, J.; Capilla, J.; Ballesteros, M.P.; Schätzlein, A.G.; et al. Oral particle uptake and organ targeting drives the activity of amphotericin B nanoparticles. Mol. Pharm. 2015, 12, 420–431. [Google Scholar] [CrossRef] [PubMed]
- de Castro Spadari, C.; Lopes, L.B.; Ishida, K. Potential Use of Alginate-Based Carriers As Antifungal Delivery System. Front. Microbiol. 2017, 8, 97. [Google Scholar] [CrossRef]
- Singh, P.K.; Jaiswal, A.K.; Pawar, V.K.; Raval, K.; Kumar, A.; Bora, H.K.; Dube, A.; Chourasia, M.K. Fabrication of 3-O-sn-Phosphatidyl-l-serine Anchored PLGA Nanoparticle Bearing Amphotericin B for Macrophage Targeting. Pharm. Res. 2018, 35, 60. [Google Scholar] [CrossRef]
- Chaudhari, M.B.; Desai, P.P.; Patel, P.A.; Patravale, V.B. Solid lipid nanoparticles of amphotericin B (AmbiOnp): In vitro and in vivo assessment towards safe and effective oral treatment module. Drug Deliv. Transl. Res. 2015, 6, 354–364. [Google Scholar] [CrossRef]
- Jain, S.; Valvi, P.U.; Swarnakar, N.K.; Thanki, K. Gelatin Coated Hybrid Lipid Nanoparticles for Oral Delivery of Amphotericin B. Mol. Pharm. 2012, 9, 2542–2553. [Google Scholar] [CrossRef]
- Money, N.P. Are mushrooms medicinal? Fungal Biol. 2016, 120, 449–453. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, S.; Wang, X.; Zhang, L.; Cheung, P.C.K. Advances in lentinan: Isolation, structure, chain conformation and bioactivities. Food Hydrocoll. 2011, 25, 196–206. [Google Scholar] [CrossRef]
- Ina, K.; Kataoka, T.; Ando, T. The use of lentinan for treating gastric cancer. Anticancer Agents Med. Chem. 2013, 13, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Yan, H.; Tang, J.; Chen, J.; Zhang, X. Polysaccharides in Lentinus edodes: Isolation, Structure, Immunomodulating Activity and Future Prospective. Crit. Rev. Food Sci. Nutr. 2014, 54, 474–487. [Google Scholar] [CrossRef] [PubMed]
- Sari, M.; Prange, A.; Lelley, J.I.; Hambitzer, R. Screening of beta-glucan contents in commercially cultivated and wild growing mushrooms. Food Chem. 2017, 216, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Hirose, A.; Sato, E.; Fujii, H.; Sun, B.; Nishioka, H.; Aruoma, O.I. The influence of active hexose correlated compound (AHCC) on cisplatin-evoked chemotherapeutic and side effects in tumor-bearing mice. Toxicol. Appl. Pharmacol. 2007, 222, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Mallet, J.F.; Graham, É.; Ritz, B.W.; Homma, K.; Matar, C. Active Hexose Correlated Compound (AHCC) promotes an intestinal immune response in BALB/c mice and in primary intestinal epithelial cell culture involving toll-like receptors TLR-2 and TLR-4. Eur. J. Nutr. 2016, 55, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Lepak, A.J.; Marchillo, K.; Vanhecker, J.; Andes, D.R. Posaconazole pharmacodynamic target determination against wild-type and Cyp51 mutant isolates of Aspergillus fumigatus in an in vivo model of invasive pulmonary aspergillosis. Antimicrob. Agents Chemother. 2013, 57, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Dixon, D.M.; Polak, A.; Walsh, T.J. Fungus dose-dependent primary pulmonary aspergillosis in immunosuppressed mice. Infect. Immun. 1989, 57, 1452–1456. [Google Scholar] [PubMed]
- Smith, P.J.; Olson, J.A.; Constable, D.; Schwartz, J.; Proffitt, R.T.; Adler-Moore, J.P. Effects of dosing regimen on accumulation, retention and prophylactic efficacy of liposomal amphotericin B. J. Antimicrob. Chemother. 2007, 59, 941–951. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Serrano, D.R.; Hernández, L.; Fleire, L.; González-Alvarez, I.; Montoya, A.; Ballesteros, M.P.; Dea-Ayuela, M.A.; Miró, G.; Bolás-Fernández, F.; Torrado, J.J. Hemolytic and pharmacokinetic studies of liposomal and particulate amphotericin B formulations. Int. J. Pharm. 2013, 447, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Genser, B.; Cooper, P.J.; Yazdanbakhsh, M.; Barreto, M.L.; Rodrigues, L.C. A guide to modern statistical analysis of immunological data. BMC Immunol. 2007, 8, 27. [Google Scholar] [CrossRef] [PubMed]
- Perlin, D.S.; Rautemaa-Richardson, R.; Alastruey-Izquierdo, A. The global problem of antifungal resistance: Prevalence, mechanisms, and management. Lancet Infect. Dis. 2017, 17, e383–e392. [Google Scholar] [CrossRef]
- Chowdhary, A.; Kathuria, S.; Xu, J.; Meis, J.F. Emergence of azole-resistant aspergillus fumigatus strains due to agricultural azole use creates an increasing threat to human health. PLoS Pathog. 2013, 9, e1003633. [Google Scholar] [CrossRef]
- Ignacio, R.M.; Kim, C.-S.; Kim, Y.-D.; Lee, H.-M.; Qi, X.-F.; Kim, S.-K. Therapeutic effect of Active Hexose-Correlated Compound (AHCC) combined with CpG-ODN (oligodeoxynucleotide) in B16 melanoma murine model. Cytokine 2015, 76, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Segarra, S.; Miró, G.; Montoya, A.; Pardo-Marín, L.; Teichenné, J.; Ferrer, L.; Cerón, J.J. Prevention of disease progression in Leishmania infantum-infected dogs with dietary nucleotides and active hexose correlated compound. Parasites Vectors 2018, 11, 103. [Google Scholar] [CrossRef] [PubMed]
- Nogusa, S.; Gerbino, J.; Ritz, B.W. Low-dose supplementation with active hexose correlated compound improves the immune response to acute influenza infection in C57BL/6 mice. Nutr. Res. 2009, 29, 139–143. [Google Scholar] [CrossRef]
- Belay, T.; Fu, C.-L.; Woart, A. Active Hexose Correlated Compound Activates Immune Function to Decrease Chlamydia trachomatis Shedding in a Murine Stress Model. J. Nutr. Med. Diet Care 2015, 1, JNMDC-1-006. [Google Scholar] [PubMed]
- Wang, S.; Welte, T.; Fang, H.; Chang, G.-J.J.; Born, W.K.; O’Brien, R.L.; Sun, B.; Fujii, H.; Kosuna, K.; Wang, T. Oral Administration of Active Hexose Correlated Compound Enhances Host Resistance to West Nile Encephalitis in Mice. J. Nutr. 2009, 139, 598–602. [Google Scholar] [CrossRef]
- Ritz, B.W. Supplementation with active hexose correlated compound increases survival following infectious challenge in mice. Nutr. Rev. 2008, 66, 526–531. [Google Scholar] [CrossRef]
- Olson, J.A.; George, A.; Constable, D.; Smith, P.; Proffitt, R.T.; Adler-Moore, J.P. Liposomal amphotericin B and echinocandins as monotherapy or sequential or concomitant therapy in murine disseminated and pulmonary Aspergillus fumigatus infections. Antimicrob. Agents Chemother. 2010, 54, 3884–3894. [Google Scholar] [CrossRef]
- Lewis, R.E.; Albert, N.D.; Kontoyiannis, D.P. Efficacy of Single-Dose Liposomal Amphotericin B or Micafungin Prophylaxis in a Neutropenic Murine Model of Invasive Pulmonary Aspergillosis. Antimicrob. Agents Chemother. 2008, 52, 4178–4180. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Vetvicka, V.; Vetvickova, J. Immune-enhancing effects of Maitake (Grifola frondosa) and Shiitake (Lentinula edodes) extracts. Ann. Transl. Med. 2014, 2, 14. [Google Scholar] [CrossRef] [PubMed]
- Daddaoua, A.; Martínez-Plata, E.; Ortega-González, M.; Ocón, B.; Aranda, C.J.; Zarzuelo, A.; Suárez, M.D.; de Medina, F.S.; Martínez-Augustin, O. The nutritional supplement Active Hexose Correlated Compound (AHCC) has direct immunomodulatory actions on intestinal epithelial cells and macrophages involving TLR/MyD88 and NF-κB/MAPK activation. Food Chem. 2013, 136, 1288–1295. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Fujii, H.; Walshe, T. Effects of active hexose correlated compound on frequency of CD4+ and CD8+ T cells producing interferon-γ and/or tumor necrosis factor–α in healthy adults. Hum. Immunol. 2010, 71, 1187–1190. [Google Scholar] [CrossRef] [PubMed]
- Thakur, R.; Anand, R.; Tiwari, S.; Singh, A.P.; Tiwary, B.N.; Shankar, J. Cytokines induce effector T-helper cells during invasive aspergillosis; what we have learned about T-helper cells? Front. Microbiol. 2015, 6, 429. [Google Scholar] [CrossRef] [PubMed]
- Dewi, I.M.W.; van de Veerdonk, F.L.; Gresnigt, M.S. The Multifaceted Role of T-Helper Responses in Host Defense against Aspergillus fumigatus. J. Fungi 2017, 3, 55. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.-W.; Lee, N.; Fujii, H.; Kang, I. Active Hexose Correlated Compound promotes T helper (Th) 17 and 1 cell responses via inducing IL-1β production from monocytes in humans. Cell. Immunol. 2012, 275, 19–23. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Heinekamp, T.; Schmidt, H.; Lapp, K.; Pähtz, V.; Shopova, I.; Köster-Eiserfunke, N.; Krüger, T.; Kniemeyer, O.; Brakhage, A.A. Interference of Aspergillus fumigatus with the immune response. Semin. Immunopathol. 2015, 37, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Goodyear, A.; Jones, A.; Troyer, R.; Bielefeldt-Ohmann, H.; Dow, S. Critical Protective Role for MCP-1 in Pneumonic Burkholderia mallei Infection. J. Immunol. 2010, 184, 1445–1454. [Google Scholar] [CrossRef] [PubMed]
- Ellis, M.; al-Ramadi, B.; Hedström, U.; Alizadeh, H.; Shammas, V.; Kristensen, J. Invasive fungal infections are associated with severe depletion of circulating RANTES. J. Med. Microbiol. 2005, 54, 1017–1022. [Google Scholar] [CrossRef] [PubMed]
- Osherov, N.; Ben-Ami, R. Modulation of Host Angiogenesis as a Microbial Survival Strategy and Therapeutic Target. PLoS Pathog. 2016, 12, e1005479. [Google Scholar] [CrossRef]
- Antachopoulos, C.; Roilides, E. Cytokines and fungal infections. Br. J. Haematol. 2005, 129, 583–596. [Google Scholar] [CrossRef] [PubMed]
- Deepe, G.S. Tumor necrosis factor-alpha and host resistance to the pathogenic fungus, Histoplasma capsulatum. J. Investig. Dermatol. Symp. Proc. 2007, 12, 34–37. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Cantero, A.; Serrano, D.R.; Navarro-Rodríguez, P.; Schätzlein, A.G.; Uchegbu, I.F.; Torrado, J.J.; Capilla, J. Increased Efficacy of Oral Fixed-Dose Combination of Amphotericin B and AHCC® Natural Adjuvant against Aspergillosis. Pharmaceutics 2019, 11, 456. https://doi.org/10.3390/pharmaceutics11090456
Pérez-Cantero A, Serrano DR, Navarro-Rodríguez P, Schätzlein AG, Uchegbu IF, Torrado JJ, Capilla J. Increased Efficacy of Oral Fixed-Dose Combination of Amphotericin B and AHCC® Natural Adjuvant against Aspergillosis. Pharmaceutics. 2019; 11(9):456. https://doi.org/10.3390/pharmaceutics11090456
Chicago/Turabian StylePérez-Cantero, Alba, Dolores R. Serrano, Patricia Navarro-Rodríguez, Andreas G. Schätzlein, Ijeoma F. Uchegbu, Juan J. Torrado, and Javier Capilla. 2019. "Increased Efficacy of Oral Fixed-Dose Combination of Amphotericin B and AHCC® Natural Adjuvant against Aspergillosis" Pharmaceutics 11, no. 9: 456. https://doi.org/10.3390/pharmaceutics11090456
APA StylePérez-Cantero, A., Serrano, D. R., Navarro-Rodríguez, P., Schätzlein, A. G., Uchegbu, I. F., Torrado, J. J., & Capilla, J. (2019). Increased Efficacy of Oral Fixed-Dose Combination of Amphotericin B and AHCC® Natural Adjuvant against Aspergillosis. Pharmaceutics, 11(9), 456. https://doi.org/10.3390/pharmaceutics11090456