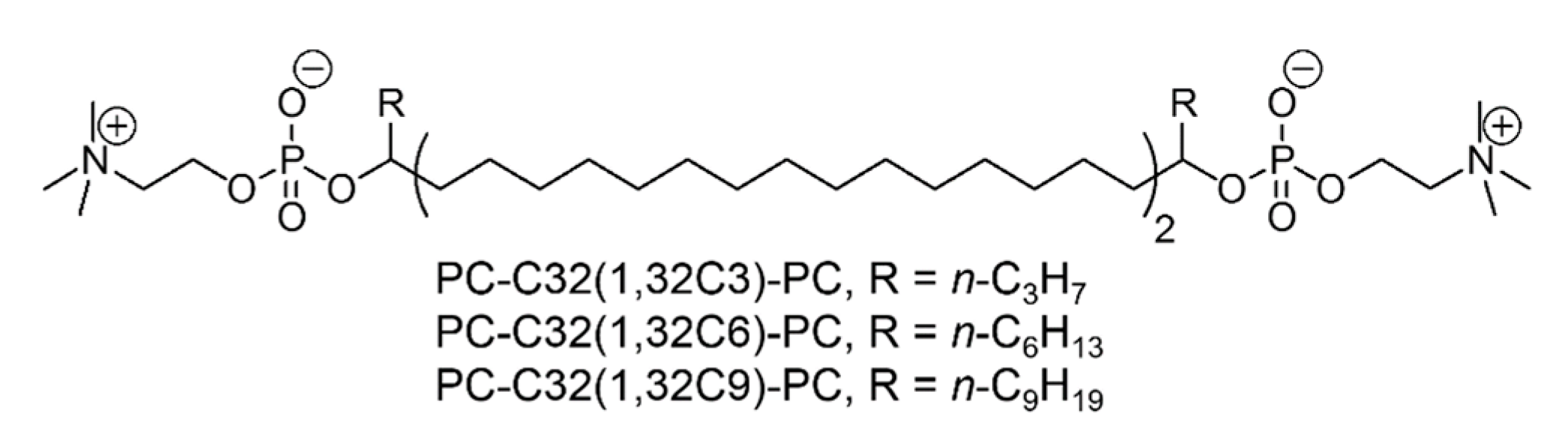
Bolalipid-Doped Liposomes: Can Bolalipids Increase the Integrity of Liposomes Exposed to Gastrointestinal Fluids?


Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Sample Preparation
2.2.2. Differential Scanning Calorimetry (DSC)
2.2.3. Transmission Electron Microscopy of Vitrified Specimens (Cryo-TEM)
2.2.4. Transmission Electron Microscopy of Replica of Freeze-Fractured Samples (FFEM)
2.2.5. Dynamic Light Scattering (DLS)
2.2.6. Dithionite Fluorescence Assay
2.2.7. Calcein Release Assay
2.2.8. Freeze-Drying and Calcein Retention
3. Results and Discussion
3.1. Miscibility of PC-C32(1,32C9)-PC with Phosphatidylcholines Derived from Soy Beans (soyPC)
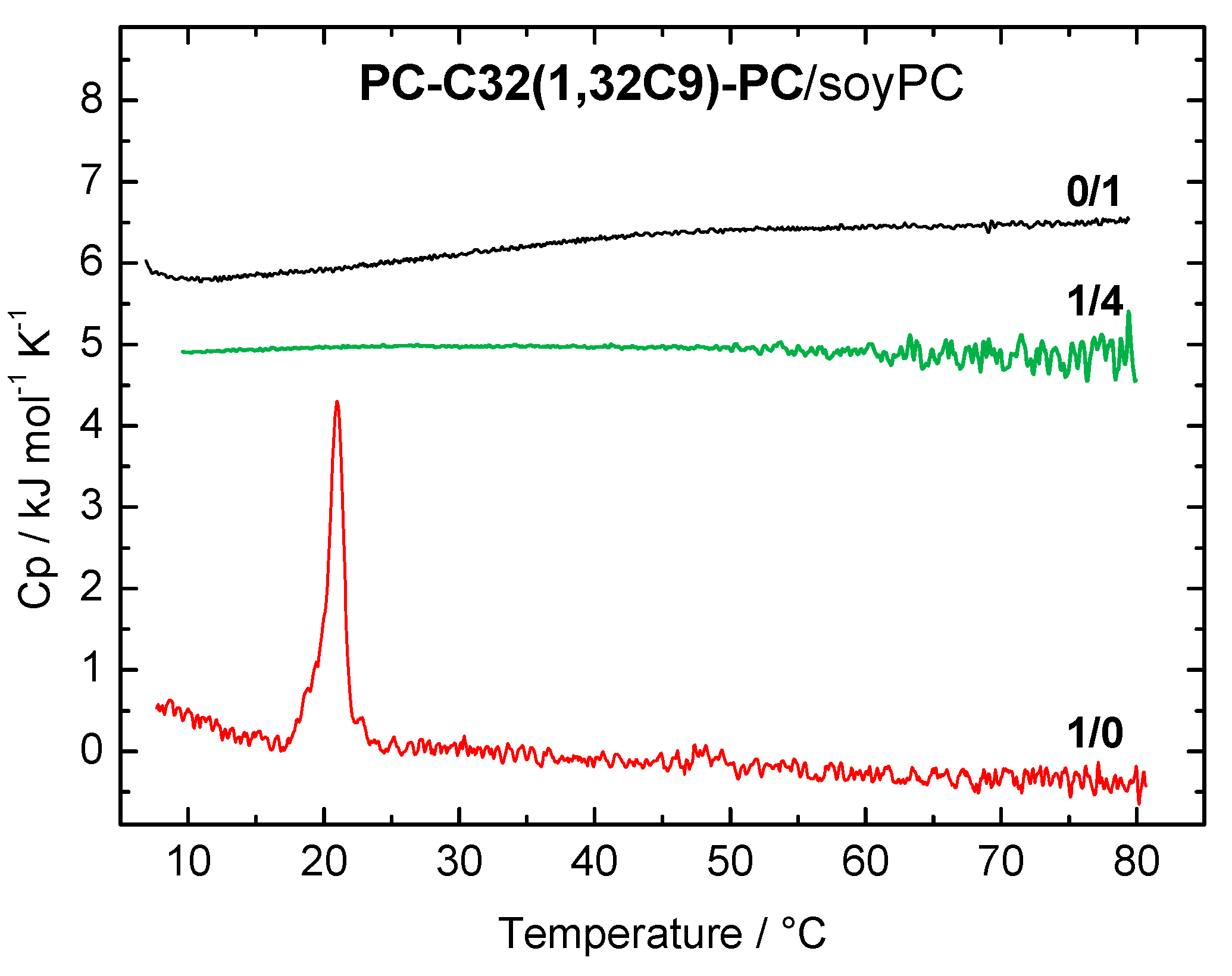
3.1.1. DSC Measurements
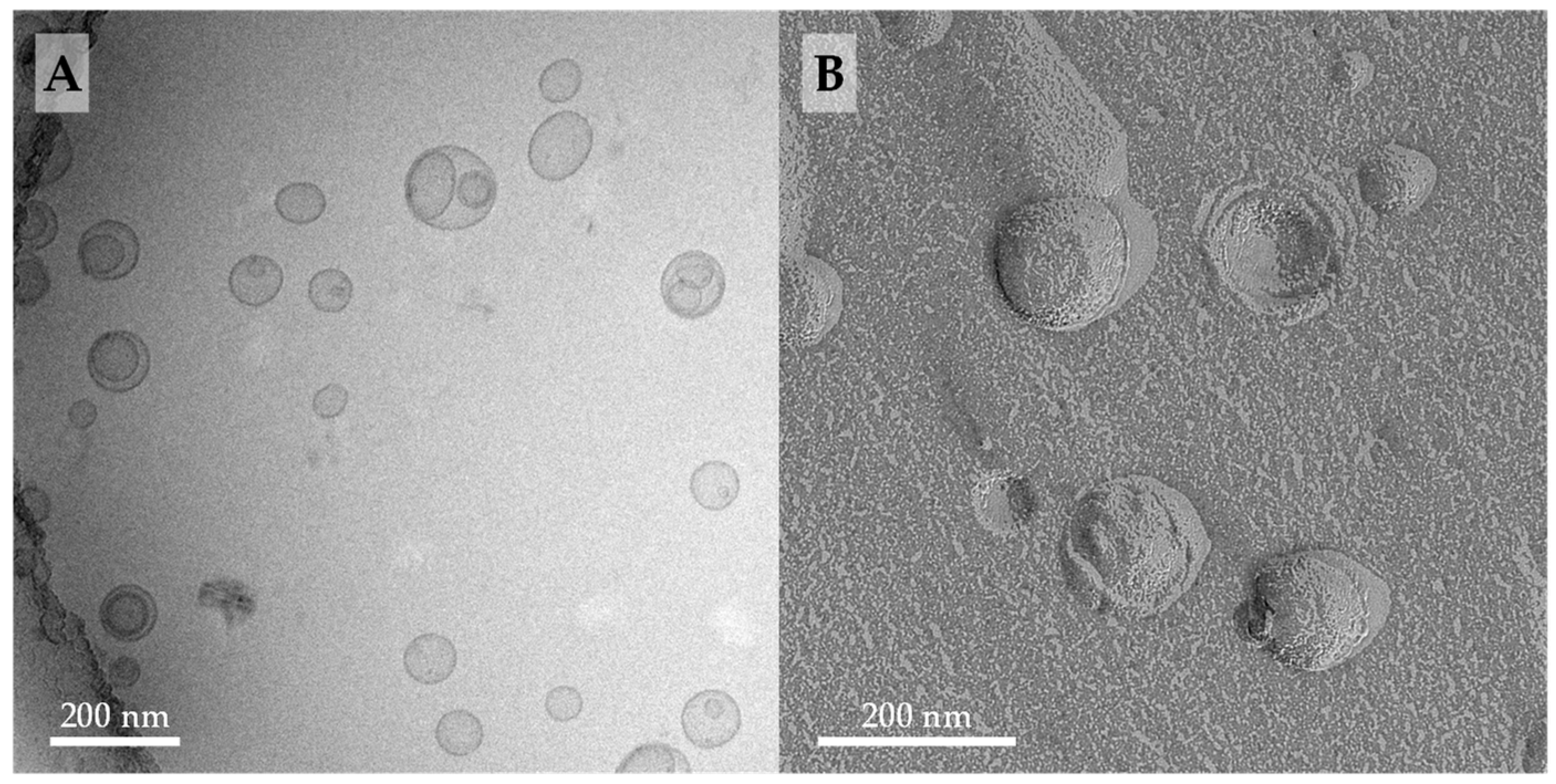
3.1.2. Cryo-TEM and FFEM Investigations
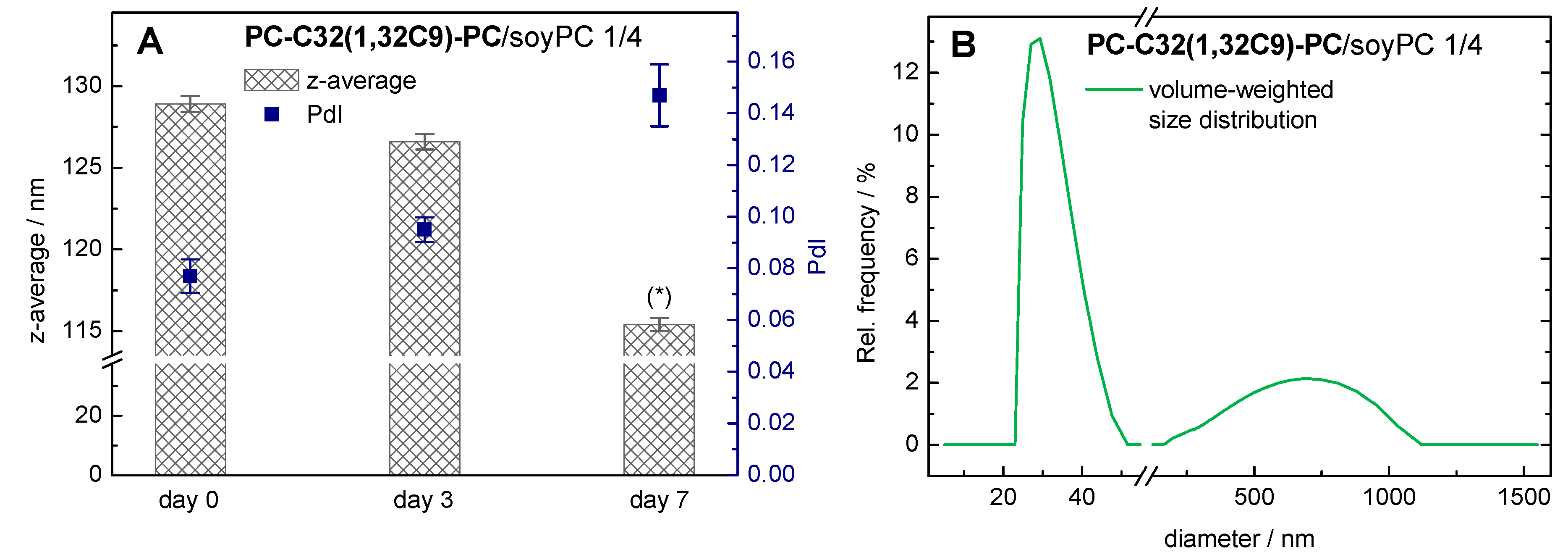
3.1.3. DLS Measurements
3.2. Integrity of Bolasomes
3.2.1. Permeability of Bolasomes Against Small Ions: The Dithionite Assay
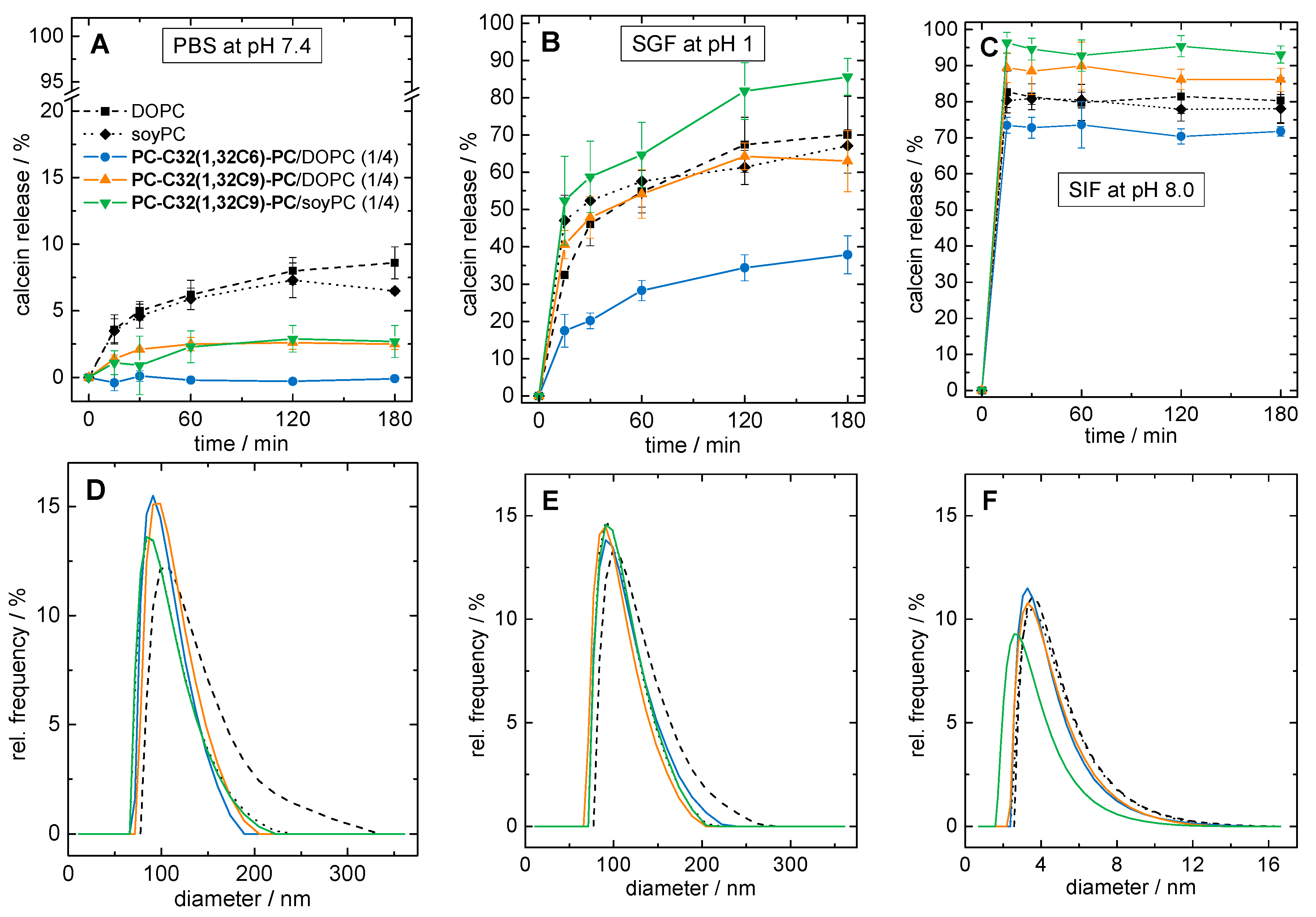
3.2.2. Stability of Bolasomes in Different Digestive Media
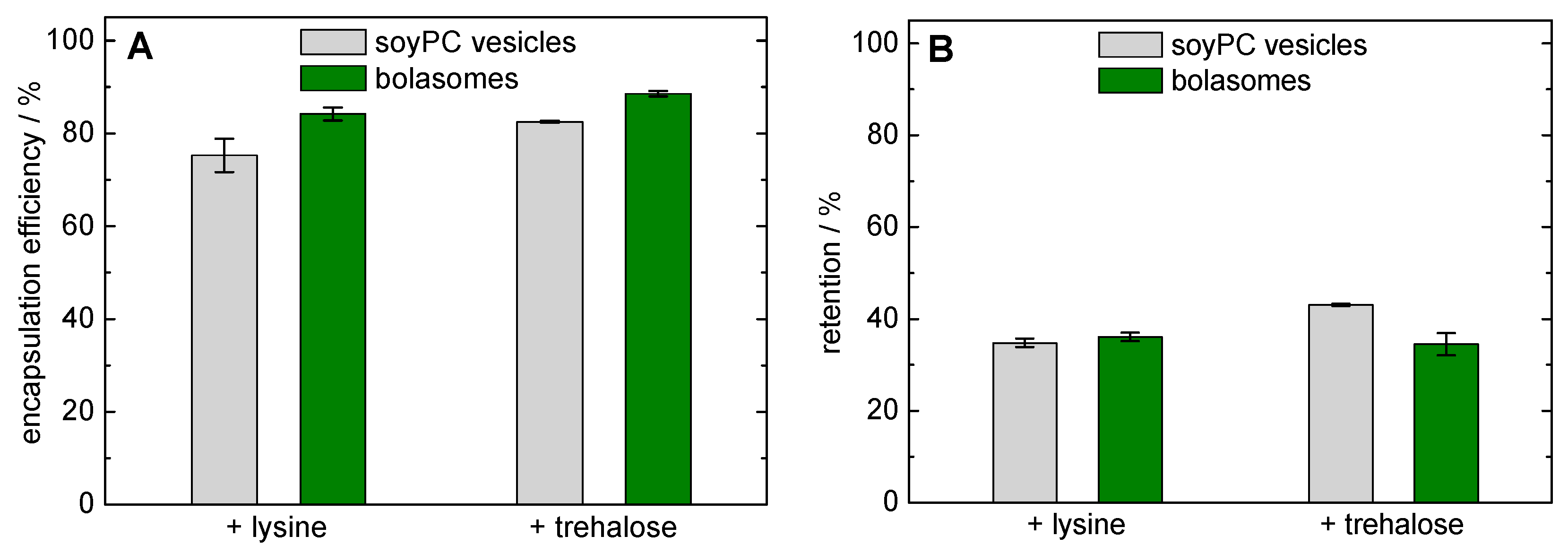
3.2.3. Freeze-Drying and Re-Hydration of Bolasomes
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Felice, B.; Prabhakaran, M.P.; Rodriguez, A.P.; Ramakrishna, S. Drug delivery vehicles on a nano-engineering perspective. Mater. Sci. Eng. C Mater. Biol. Appl. 2014, 41, 178–195. [Google Scholar] [CrossRef] [PubMed]
- Bozzuto, G.; Molinari, A. Liposomes as nanomedical devices. Int. J. Nanomed. 2015, 10, 975–999. [Google Scholar] [CrossRef] [PubMed]
- Sessa, G.; Weissmann, G. Incorporation of lysozyme into liposomes. A model for structure-linked latency. J. Biol. Chem. 1970, 245, 3295–3301. [Google Scholar] [PubMed]
- Li, H.; Song, J.-H.; Park, J.-S.; Han, K. Polyethylene glycol-coated liposomes for oral delivery of recombinant human epidermal growth factor. Int. J. Pharm. 2003, 258, 11–19. [Google Scholar] [CrossRef]
- Chen, Y.; Lu, Y.; Chen, J.; Lai, J.; Sun, J.; Hu, F.; Wu, W. Enhanced bioavailability of the poorly water-soluble drug fenofibrate by using liposomes containing a bile salt. Int. J. Pharm. 2009, 376, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Khandelwal, K.; Pachauri, S.D.; Arya, A.; Pawar, V.K.; Joshi, T.; Dwivedi, P.; Ahmad, H.; Singh, B.; Sharma, K.; Kanojiya, S.; et al. Improved oral bioavailability of novel antithrombotic s002-333 via chitosan coated liposomes: A pharmacokinetic assessment. RSC Adv. 2015, 5, 39168–39176. [Google Scholar] [CrossRef]
- Al-Remawi, M.; Elsayed, A.; Maghrabi, I.; Hamaidi, M.; Jaber, N. Chitosan/lecithin liposomal nanovesicles as an oral insulin delivery system. Pharm. Dev. Technol. 2017, 22, 390–398. [Google Scholar] [CrossRef]
- Daeihamed, M.; Dadashzadeh, S.; Haeri, A.; Akhlaghi, M.F. Potential of liposomes for enhancement of oral drug absorption. Curr. Drug Deliv. 2017, 14, 289–303. [Google Scholar] [CrossRef]
- Abu, L.A.S.; Ishida, T. Liposomal delivery systems: Design optimization and current applications. Biol. Pharm. Bull. 2017, 40, 1–10. [Google Scholar]
- Moen, M.D.; Lyseng-Williamson, K.A.; Scott, L.J. Liposomal amphotericin b: A review of its use as empirical therapy in febrile neutropenia and in the treatment of invasive fungal infections. Drugs 2009, 69, 361–392. [Google Scholar] [CrossRef] [PubMed]
- Rafiyath, S.M.; Rasul, M.; Lee, B.; Wei, G.; Lamba, G.; Liu, D. Comparison of safety and toxicity of liposomal doxorubicin vs. Conventional anthracyclines: A meta-analysis. Exp. Hematol. Oncol. 2012, 1, 10. [Google Scholar] [CrossRef] [PubMed]
- Silverman, J.A.; Reynolds, L.; Deitcher, S.R. Pharmacokinetics and pharmacodynamics of vincristine sulfate liposome injection (vsli) in adults with acute lymphoblastic leukemia. J. Clin. Pharmacol. 2013, 53, 1139–1145. [Google Scholar] [CrossRef] [PubMed]
- Rowland, R.N.; Woodley, J.F. The stability of liposomes in vitro to ph, bile salts and pancreatic lipase. Biochim. Biophys. Acta 1980, 620, 400–409. [Google Scholar] [CrossRef]
- Walde, P.; Sunamoto, J.; O’Connor, C.J. The mechanism of liposomal damage by taurocholate. Biochim. Biophys. Acta Biomembr. 1987, 905, 30–38. [Google Scholar] [CrossRef]
- Fricker, G.; Kromp, T.; Wendel, A.; Blume, A.; Zirkel, J.; Rebmann, H.; Setzer, C.; Quinkert, R.-O.; Martin, F.; Müller-Goymann, C. Phospholipids and lipid-based formulations in oral drug delivery. Pharm. Res. 2010, 27, 1469–1486. [Google Scholar] [CrossRef] [PubMed]
- Parmentier, J.; Thomas, N.; Muellertz, A.; Fricker, G.; Rades, T. Exploring the fate of liposomes in the intestine by dynamic in vitro lipolysis. Int. J. Pharm. 2012, 437, 253–263. [Google Scholar] [CrossRef]
- Liu, W.; Ye, A.; Liu, W.; Liu, C.; Han, J.; Singh, H. Behavior of liposomes loaded with bovine serum albumin during in vitro digestion. Food Chem. 2015, 175, 16–24. [Google Scholar] [CrossRef]
- van Hoogevest, P. Review—An update on the use of oral phospholipid excipients. Eur. J. Pharm. Sci. 2017, 108, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sanitt, P.; Apiratikul, N.; Niyomtham, N.; Yingyongnarongkul, B.E.; Assavalapsakul, W.; Panyim, S.; Udomkit, A. Cholesterol-based cationic liposome increases dsrna protection of yellow head virus infection in penaeus vannamei. J. Biotechnol. 2016, 228, 95–102. [Google Scholar] [CrossRef]
- He, H.; Lu, Y.; Qi, J.; Zhu, Q.; Chen, Z.; Wu, W. Adapting liposomes for oral drug delivery. Acta Pharm. Sin. B 2019, 9, 36–48. [Google Scholar] [CrossRef]
- Thongborisute, J.; Tsuruta, A.; Kawabata, Y.; Takeuchi, H. The effect of particle structure of chitosan-coated liposomes and type of chitosan on oral delivery of calcitonin. J. Drug Target. 2006, 14, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wu, J.; Sun, M.; Guo, C.; Yu, A.; Cao, F.; Zhao, L.; Tan, Q.; Zhai, G. N-trimethyl chitosan chloride-coated liposomes for the oral delivery of curcumin. J. Liposome Res. 2012, 22, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.K.; Harde, H.; Thanki, K.; Jain, S. Improved stability and antidiabetic potential of insulin containing folic acid functionalized polymer stabilized multilayered liposomes following oral administration. Biomacromolecules 2014, 15, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.; Su, Z.; Li, S.; Sun, M.; Xiao, Y.; Ping, Q.; Deng, Y. Oral absorption enhancement of salmon calcitonin by using both n-trimethyl chitosan chloride and oligoarginines-modified liposomes as the carriers. Drug Deliv. 2014, 21, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Han, Y.; Chen, C.; Cao, Y.; Wang, S.; Shen, W.; Zhang, H.; Li, Y.; van Dongen, M.A.; He, B.; et al. Oral absorption enhancement of probucol by pegylated g5 pamam dendrimer modified nanoliposomes. Mol. Pharm. 2015, 12, 665–674. [Google Scholar] [CrossRef]
- Song, K.-H.; Chung, S.-J.; Shim, C.-K. Enhanced intestinal absorption of salmon calcitonin (sct) from proliposomes containing bile salts. J. Controll. Release 2005, 106, 298–308. [Google Scholar] [CrossRef]
- Ayogu, I.J.; Ogbonna, O.; Ayolugbe, C.I.; Attama, A.A. Evaluation of the pharmacodynamic activity of insulin from bilosomal formulation. Curr. Drug Deliv. 2009, 6, 415–418. [Google Scholar] [CrossRef]
- Guan, P.; Lu, Y.; Qi, J.; Niu, M.; Lian, R.; Hu, F.; Wu, W. Enhanced oral bioavailability of cyclosporine a by liposomes containing a bile salt. Int. J. Nanomed. 2011, 6, 965–974. [Google Scholar]
- Aburahma, M.H. Bile salts-containing vesicles: Promising pharmaceutical carriers for oral delivery of poorly water-soluble drugs and peptide/protein-based therapeutics or vaccines. Drug Deliv. 2016, 23, 1847–1867. [Google Scholar] [CrossRef]
- Shukla, A.; Mishra, V.; Kesharwani, P. Bilosomes in the context of oral immunization: Development challenges and opportunities. Drug Discov. Today 2016, 21, 888–899. [Google Scholar] [CrossRef]
- Fuhrhop, J.-H.; Wang, T. Bolaamphiphiles. Chem. Rev. 2004, 104, 2901–2937. [Google Scholar] [CrossRef] [PubMed]
- Elferink, M.G.L.; Dewit, J.G.; Driessen, A.J.M.; Konings, W.N. Stability and proton-permeability of liposomes composed of archaeal tetraether lipids. Biochim. Biophys. Acta Biomembr. 1994, 1193, 247–254. [Google Scholar] [CrossRef]
- Koyanagi, T.; Leriche, G.; Yep, A.; Onofrei, D.; Holland, G.P.; Mayer, M.; Yang, J. Effect of headgroups on small-ion permeability across archaea-inspired tetraether lipid membranes. Chem. A Eur. J. 2016, 22, 8074–8077. [Google Scholar] [CrossRef] [PubMed]
- Koyanagi, T.; Leriche, G.; Onofrei, D.; Holland, G.P.; Mayer, M.; Yang, J. Cyclohexane rings reduce membrane permeability to small ions in archaea-inspired tetraether lipids. Angew. Chem. Int. Ed. 2016, 55, 1890–1893. [Google Scholar] [CrossRef]
- Parmentier, J.; Thewes, B.; Gropp, F.; Fricker, G. Oral peptide delivery by tetraether lipid liposomes. Int. J. Pharm. 2011, 415, 150–157. [Google Scholar] [CrossRef]
- Parmentier, J.; Hofhaus, G.; Thomas, S.; Cuesta, L.C.; Gropp, F.; Schroeder, R.; Hartmann, K.; Fricker, G. Improved oral bioavailability of human growth hormone by a combination of liposomes containing bio-enhancers and tetraether lipids and omeprazole. J. Pharm. Sci. 2014, 103, 3985–3993. [Google Scholar] [CrossRef]
- Uhl, P.; Helm, F.; Hofhaus, G.; Brings, S.; Kaufman, C.; Leotta, K.; Urban, S.; Haberkorn, U.; Mier, W.; Fricker, G. A liposomal formulation for the oral application of the investigational hepatitis b drug myrcludex b. Eur. J. Pharm. Biopharm. 2016, 103, 159–166. [Google Scholar] [CrossRef]
- Patel, G.B.; Agnew, B.J.; Deschatelets, L.; Fleming, L.P.; Sprott, G.D. In vitro assessment of archaeosome stability for developing oral delivery systems. Int. J. Pharm. 2000, 194, 39–49. [Google Scholar] [CrossRef]
- Kaur, G.; Garg, T.; Rath, G.; Goyal, A.K. Archaeosomes: An excellent carrier for drug and cell delivery. Drug Deliv. 2016, 23, 2497–2512. [Google Scholar] [CrossRef]
- Brown, D.A.; Venegas, B.; Cooke, P.H.; English, V.; Chong, P.L.-G. Bipolar tetraether archaeosomes exhibit unusual stability against autoclaving as studied by dynamic light scattering and electron microscopy. Chem. Phys. Lipids 2009, 159, 95–103. [Google Scholar] [CrossRef]
- Omri, A.; Agnew, B.J.; Patel, G.B. Short-term repeated-dose toxicity profile of archaeosomes administered to mice via intravenous and oral routes. Int. J. Toxicol. 2003, 22, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, T.; Ibaragi, K.; Kakinuma, K. Total synthesis of archaeal 72-membered macrocyclic tetraether lipids. J. Org. Chem. 1998, 63, 2689–2698. [Google Scholar] [CrossRef] [PubMed]
- Patel, G.B.; Sprott, G.D. Archaeobacterial ether lipid liposomes (archaeosomes) as novel vaccine and drug delivery systems. Crit. Rev. Biotechnol. 1999, 19, 317–357. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, A.C.; Jensen, S.M.; Fricker, G.; Brandl, M.; Treusch, A.H. Archaeal lipids in oral delivery of therapeutic peptides. Eur. J. Pharm. Sci. 2017, 108, 101–110. [Google Scholar] [CrossRef]
- Köhler, K.; Förster, G.; Hauser, A.; Dobner, B.; Heiser, U.F.; Ziethe, F.; Richter, W.; Steiniger, F.; Drechsler, M.; Stettin, H.; et al. Self-assembly in a bipolar phosphocholine-water system: The formation of nanofibers and hydrogels. Angew. Chem. Int. Ed. 2004, 43, 245–247. [Google Scholar] [CrossRef]
- Meister, A.; Köhler, K.; Drescher, S.; Dobner, B.; Karlsson, G.; Edwards, K.; Hause, G.; Blume, A. Mixing behaviour of a symmetrical single-chain bolaamphiphile with phospholipids. Soft Matter 2007, 3, 1025–1031. [Google Scholar] [CrossRef]
- Drescher, S.; Sonnenberger, S.; Meister, A.; Blume, A.; Dobner, B. Synthesis and aggregation behaviour of symmetrical phenylene/biphenylene-modified bolaamphiphiles. Mon. Chem. 2012, 143, 1533–1543. [Google Scholar] [CrossRef]
- Drescher, S.; Becker, S.; Dobner, B.; Blume, A. Bis-sonogashira cross-coupling: An expeditious approach towards long-chain, phenylene-modified 1, ω-diols. RSC Adv. 2012, 2, 4052–4054. [Google Scholar] [CrossRef]
- Drescher, S.; Meister, A.; Garamus, V.M.; Hause, G.; Garvey, C.J.; Dobner, B.; Blume, A. Influence of the substitution pattern of phenylene-modified bolaamphiphiles on the aggregations behavior and the miscibility with conventional phospholipids. Eur. J. Lipid Sci. Technol. 2014, 116, 1205–1216. [Google Scholar] [CrossRef]
- Drescher, S.; Lechner, B.-D.; Garamus, V.M.; Almásy, L.; Meister, A.; Blume, A. The headgroup (a) symmetry strongly determines the aggregation behavior of single-chain phenylene-modified bolalipids and their miscibility with classical phospholipids. Langmuir 2014, 30, 9273–9284. [Google Scholar] [CrossRef]
- Drescher, S.; Garamus, V.M.; Garvey, C.J.; Meister, A.; Blume, A. Aggregation behaviour of a single-chain, phenylene-modified bolalipid and its miscibility with classical phospholipids. Beilstein J. Org. Chem. 2017, 13, 995–1007. [Google Scholar] [CrossRef] [PubMed]
- Gruhle, K.; Müller, S.; Meister, A.; Drescher, S. Synthesis and aggregation behaviour of single-chain, 1, 32-alkyl branched bis(phosphocholines): Effect of lateral chain length. Org. Biomol. Chem. 2018, 16, 2711–2724. [Google Scholar] [CrossRef] [PubMed]
- Müller, S.; Kind, M.; Gruhle, K.; Hause, G.; Meister, A.; Drescher, S. Mixing behaviour of bilayer-forming phosphatidylcholines with single-chain alkyl-branched bolalipids: Effect of lateral chain length. Biophys. Chem. 2019, 244, 1–10. [Google Scholar] [CrossRef] [PubMed]
- van Hoogevest, P.; Wendel, A. The use of natural and synthetic phospholipids as pharmaceutical excipients. Eur. J. Lipid Sci. Technol. 2014, 116, 1088–1107. [Google Scholar] [CrossRef]
- Bangham, A.D.; Standish, M.M.; Watkins, J.C. Diffusion of univalent ions across the lamellae of swollen phospholipids. J. Mol. Biol. 1965, 13, 238–252. [Google Scholar] [CrossRef]
- Müller, S.; Meister, A.; Otto, C.; Hause, G.; Drescher, S. Mixing behaviour of asymmetrical glycerol diether bolalipids with saturated and unsaturated phosphatidylcholines. Biophys. Chem. 2018, 238, 39–48. [Google Scholar] [CrossRef]
- McIntyre, J.C.; Sleight, R.G. Fluorescence assay for phospholipid membrane asymmetry. Biochemistry 1991, 30, 11819–11827. [Google Scholar] [CrossRef]
- Maherani, B.; Arab-Tehrany, E.; Kheirolomoom, A.; Geny, D.; Linder, M. Calcein release behavior from liposomal bilayer; influence of physicochemical/mechanical/structural properties of lipids. Biochimie 2013, 95, 2018–2033. [Google Scholar] [CrossRef]
- Crowe, L.M.; Crowe, J.H.; Rudolph, A.; Womersley, C.; Appel, L. Preservation of freeze-dried liposomes by trehalose. Arch. Biochem. Biophys. 1985, 242, 240–247. [Google Scholar] [CrossRef]
- Wang, T.; Hammond, E.G.; Fehr, W.R. Neutral and polar lipid phase transition of soybeans with various saturated fatty acid contents. J. Am. Oil Chem. Soc. 2001, 78, 1139–1144. [Google Scholar] [CrossRef]
- Baxa, U. Imaging of liposomes by transmission electron microscopy. In Characterization of Nanoparticles Intended for Drug Delivery; McNeil, S.E., Ed.; Springer: New York, NY, USA, 2018; pp. 73–88. [Google Scholar]
- Meister, A.; Blume, A. (Cryo) transmission electron microscopy of phospholipid model membranes interacting with amphiphilic and polyphilic molecules. Polymers 2017, 9, 521. [Google Scholar] [CrossRef] [PubMed]
- Rawicz, W.; Olbrich, K.C.; McIntosh, T.; Needham, D.; Evans, E. Effect of chain length and unsaturation on elasticity of lipid bilayers. Biophys. J. 2000, 79, 328–339. [Google Scholar] [CrossRef]
- Beveridge, T.J.; Choquet, C.G.; Patel, G.B.; Sprott, G.D. Freeze-fracture planes of methanogen membranes correlate with the content of tetraether lipids. J. Bacteriol. 1993, 175, 1191–1197. [Google Scholar] [CrossRef] [PubMed]
- Hassan, P.A.; Rana, S.; Verma, G. Making sense of brownian motion: Colloid characterization by dynamic light scattering. Langmuir 2015, 31, 3–12. [Google Scholar] [CrossRef]
- Froekjaer, S.; Hjorth, E.L.; Woerts, O. Stability and storage of liposomes. Alfred Benzon Symp. 1982, 17, 384–401. [Google Scholar]
- Hager, A.A.; De Paoli, T.; Ihlo, J.E.; Farach, H.A.; Poole, C.P. Stability study of lecithin liposomes during storage using esr. Spectrochim. Acta Part A Mol. Spectrosc. 1993, 49, 1999–2005. [Google Scholar] [CrossRef]
- van de Vossenberg, J.L.C.M.; Ubbink-Kok, T.; Elferink, M.G.L.; Driessen, A.J.M.; Konings, W.N. Ion permeability of the cytoplasmic membrane limits the maximum growth temperature of bacteria and archaea. Mol. Microbiol. 1995, 18, 925–932. [Google Scholar] [CrossRef]
- Langner, M.; Hui, S.W. Dithionite penetration through phospholipid bilayers as a measure of defects in lipid molecular packing. Chem. Phys. Lipids 1993, 65, 23–30. [Google Scholar] [CrossRef]
- Balch, C.; Morris, R.; Brooks, E.; Sleight, R.G. The use of n-(7-nitrobenz-2-oxa-1,3-diazole-4-yl)-labeled lipids in determining transmembrane lipid distribution. Chem. Phys. Lipids 1994, 70, 205–212. [Google Scholar] [CrossRef]
- Angeletti, C.; Nichols, J.W. Dithionite quenching rate measurement of the inside-outside membrane bilayer distribution of 7-nitrobenz-2-oxa-1,3-diazol-4-yl-labeled phospholipids. Biochemistry 1998, 37, 15114–15119. [Google Scholar] [CrossRef]
- Moreno, M.J.; Estronca, L.M.; Vaz, W.L. Translocation of phospholipids and dithionite permeability in liquid-ordered and liquid-disordered membranes. Biophys. J. 2006, 91, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Zhou, H.; Li, P.Y.; Speer, J.E.; Cheng, H. Structural elucidation of cell membrane-derived nanoparticles using molecular probes. J. Mater. Chem. B 2014, 2, 8231–8238. [Google Scholar] [CrossRef]
- Shinoda, W. Permeability across lipid membranes. Biochim. Biophys. Acta Biomembr. 2016, 1858, 2254–2265. [Google Scholar] [CrossRef] [PubMed]
- John, K.; Schreiber, S.; Kubelt, J.; Herrmann, A.; Muller, P. Transbilayer movement of phospholipids at the main phase transition of lipid membranes: Implications for rapid flip-flop in biological membranes. Biophys. J. 2002, 83, 3315–3323. [Google Scholar] [CrossRef]
- Liu, W.; Ye, A.; Liu, C.; Liu, W.; Singh, H. Structure and integrity of liposomes prepared from milk- or soybean-derived phospholipids during in vitro digestion. Food Res. Int. 2012, 48, 499–506. [Google Scholar] [CrossRef]
- Andrieux, K.; Forte, L.; Lesieur, S.; Paternostre, M.; Ollivon, M.; Grabielle-Madelmont, C. Solubilisation of dipalmitoylphosphatidylcholine bilayers by sodium taurocholate: A model to study the stability of liposomes in the gastrointestinal tract and their mechanism of interaction with a model bile salt. Eur. J. Pharm. Biopharm. 2009, 71, 346–355. [Google Scholar] [CrossRef]
- Birru, W.A.; Warren, D.B.; Ibrahim, A.; Williams, H.D.; Benameur, H.; Porter, C.J.H.; Chalmers, D.K.; Pouton, C.W. Digestion of phospholipids after secretion of bile into the duodenum changes the phase behavior of bile components. Mol. Pharm. 2014, 11, 2825–2834. [Google Scholar] [CrossRef]
- Mueller, K.; Schuster, A. Solubilization of multilamellar liposomes of egg yolk lecithin by the bile salt sodium taurodeoxycholate and the effect of cholesterol—A rapid-ultrafiltration study. Chem. Phys. Lipids 1990, 52, 111–127. [Google Scholar] [CrossRef]
- Parmentier, J.; Becker, M.M.M.; Heintz, U.; Fricker, G. Stability of liposomes containing bio-enhancers and tetraether lipids in simulated gastro-intestinal fluids. Int. J. Pharm. 2011, 405, 210–217. [Google Scholar] [CrossRef]
- Immordino, M.L.; Dosio, F.; Cattel, L. Stealth liposomes: Review of the basic science, rationale, and clinical applications, existing and potential. Int. J. Nanomed. 2006, 1, 297–315. [Google Scholar]
- Barenholz, Y. Doxil—The first fda-approved nano-drug: Lessons learned. J. Control. Release 2012, 160, 117–134. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, C.; Sivna, P.L.; Rane, S.; Yadav, R.; Rao, S.P.; Kumar, B.; Dewangan, M.K.; Sinha, D. Liposome: A powerful approach for vesicular drug delivery system. World J. Pharm. Res. 2015, 4, 1110–1141. [Google Scholar]
- van Winden, E.C.A. Freeze-drying of liposomes: Theory and practice. In Methods in Enzymology; Academic Press: New York, NY, USA, 2003; Volume 367, pp. 99–110. [Google Scholar]
- van den Hoven, J.M.; Metselaar, J.M.; Storm, G.; Beijnen, J.H.; Nuijen, B. Cyclodextrin as membrane protectant in spray-drying and freeze-drying of pegylated liposomes. Int. J. Pharm. 2012, 438, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Hu, X.; Shen, B.; Xie, Y.; Shen, C.; Lu, Y.; Qi, J.; Yuan, H.; Wu, W. Enhanced stability of liposomes against solidification stress during freeze-drying and spray-drying by coating with calcium alginate. J. Drug Deliv. Sci. Technol. 2015, 30, 163–170. [Google Scholar] [CrossRef]
- Guan, P.; Lu, Y.; Qi, J.; Niu, M.; Lian, R.; Wu, W. Solidification of liposomes by freeze-drying: The importance of incorporating gelatin as interior support on enhanced physical stability. Int. J. Pharm. 2015, 478, 655–664. [Google Scholar] [CrossRef]
- Franzé, S.; Selmin, F.; Samaritani, E.; Minghetti, P.; Cilurzo, F. Lyophilization of liposomal formulations: Still necessary, still challenging. Pharmaceutics 2018, 10, 139. [Google Scholar] [CrossRef]
- Chen, C.; Han, D.; Cai, C.; Tang, X. An overview of liposome lyophilization and its future potential. J. Control. Release 2010, 142, 299–311. [Google Scholar] [CrossRef]
- Crowe, J.H.; Hoekstra, F.A.; Nguyen, K.H.N.; Crowe, L.M. Is vitrification involved in depression of the phase transition temperature in dry phospholipids? Biochim. Biophys. Acta Biomembr. 1996, 1280, 187–196. [Google Scholar] [CrossRef]
- Mohammed, A.R.; Coombes, A.G.A.; Perrie, Y. Amino acids as cryoprotectants for liposomal delivery systems. Eur. J. Pharm. Sci. 2007, 30, 406–413. [Google Scholar] [CrossRef]
- Tanaka, K.; Takeda, T.; Fujii, K.; Miyajima, K. Cryoprotective mechanism of saccharides on freeze-drying of liposome. Chem. Pharm. Bullet. 1992, 40, 1–5. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vesicle Composition | Exponential Fit 1 | Inner NBD Lipids (%) 2 | |||
|---|---|---|---|---|---|
| a | k | c | R2 | ||
| DOPC | 0.310 | 3.0 × 10−3 | 0.224 | 0.996 | 55.4 |
| PC-C32(1,32C6)-PC/DOPC (1/4) | 0.396 | 5.8 × 10−3 | 0.115 | 0.999 | 51.1 |
| PC-C32(1,32C9)-PC/DOPC (1/4) | 0.318 | 7.1 × 10−3 | 0.191 | 0.999 | 57.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Müller, S.; Gruhle, K.; Meister, A.; Hause, G.; Drescher, S. Bolalipid-Doped Liposomes: Can Bolalipids Increase the Integrity of Liposomes Exposed to Gastrointestinal Fluids? Pharmaceutics 2019, 11, 646. https://doi.org/10.3390/pharmaceutics11120646
Müller S, Gruhle K, Meister A, Hause G, Drescher S. Bolalipid-Doped Liposomes: Can Bolalipids Increase the Integrity of Liposomes Exposed to Gastrointestinal Fluids? Pharmaceutics. 2019; 11(12):646. https://doi.org/10.3390/pharmaceutics11120646
Chicago/Turabian StyleMüller, Sindy, Kai Gruhle, Annette Meister, Gerd Hause, and Simon Drescher. 2019. "Bolalipid-Doped Liposomes: Can Bolalipids Increase the Integrity of Liposomes Exposed to Gastrointestinal Fluids?" Pharmaceutics 11, no. 12: 646. https://doi.org/10.3390/pharmaceutics11120646
APA StyleMüller, S., Gruhle, K., Meister, A., Hause, G., & Drescher, S. (2019). Bolalipid-Doped Liposomes: Can Bolalipids Increase the Integrity of Liposomes Exposed to Gastrointestinal Fluids? Pharmaceutics, 11(12), 646. https://doi.org/10.3390/pharmaceutics11120646