
The Astrovirus Capsid: A Review

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
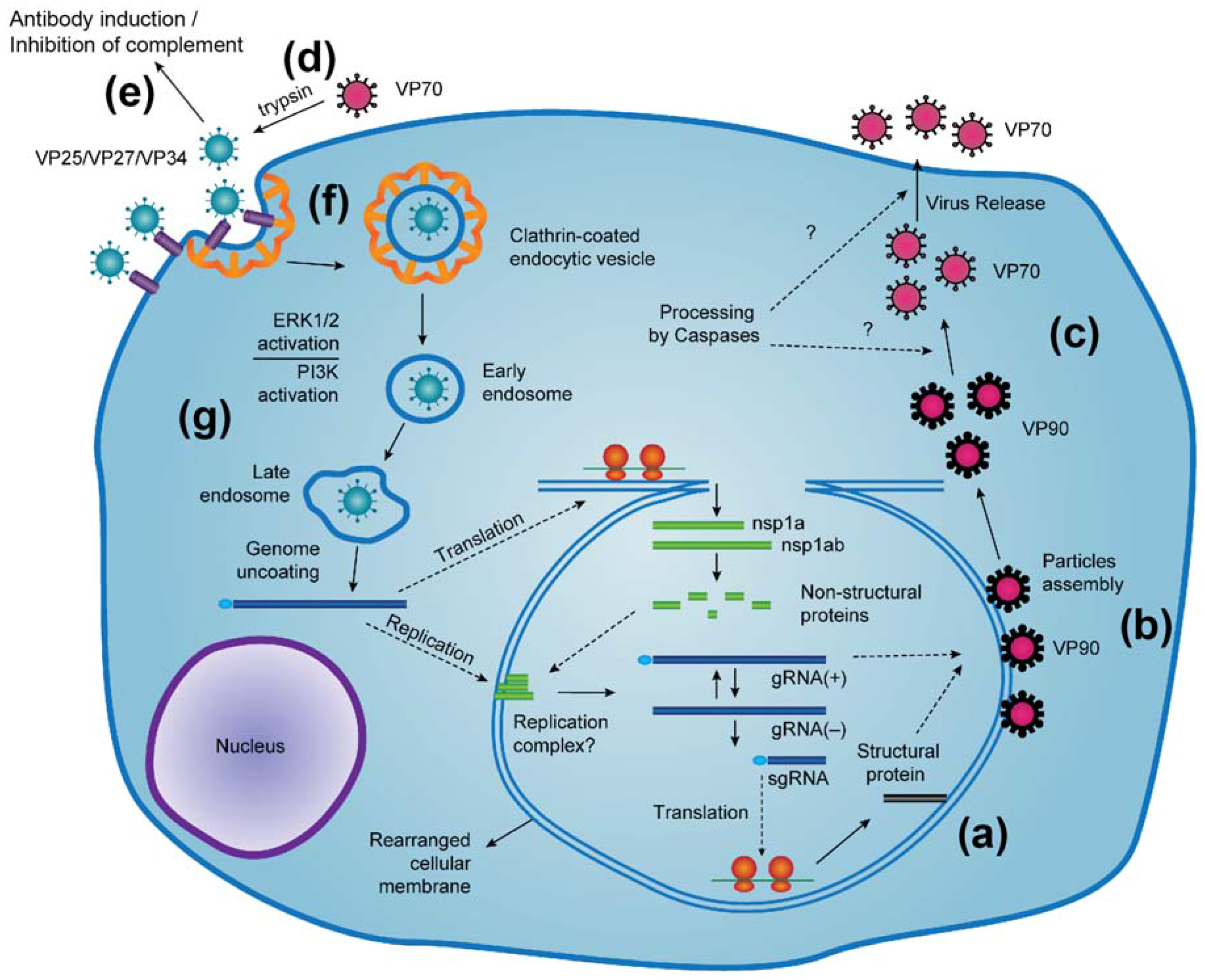
:1. Introduction
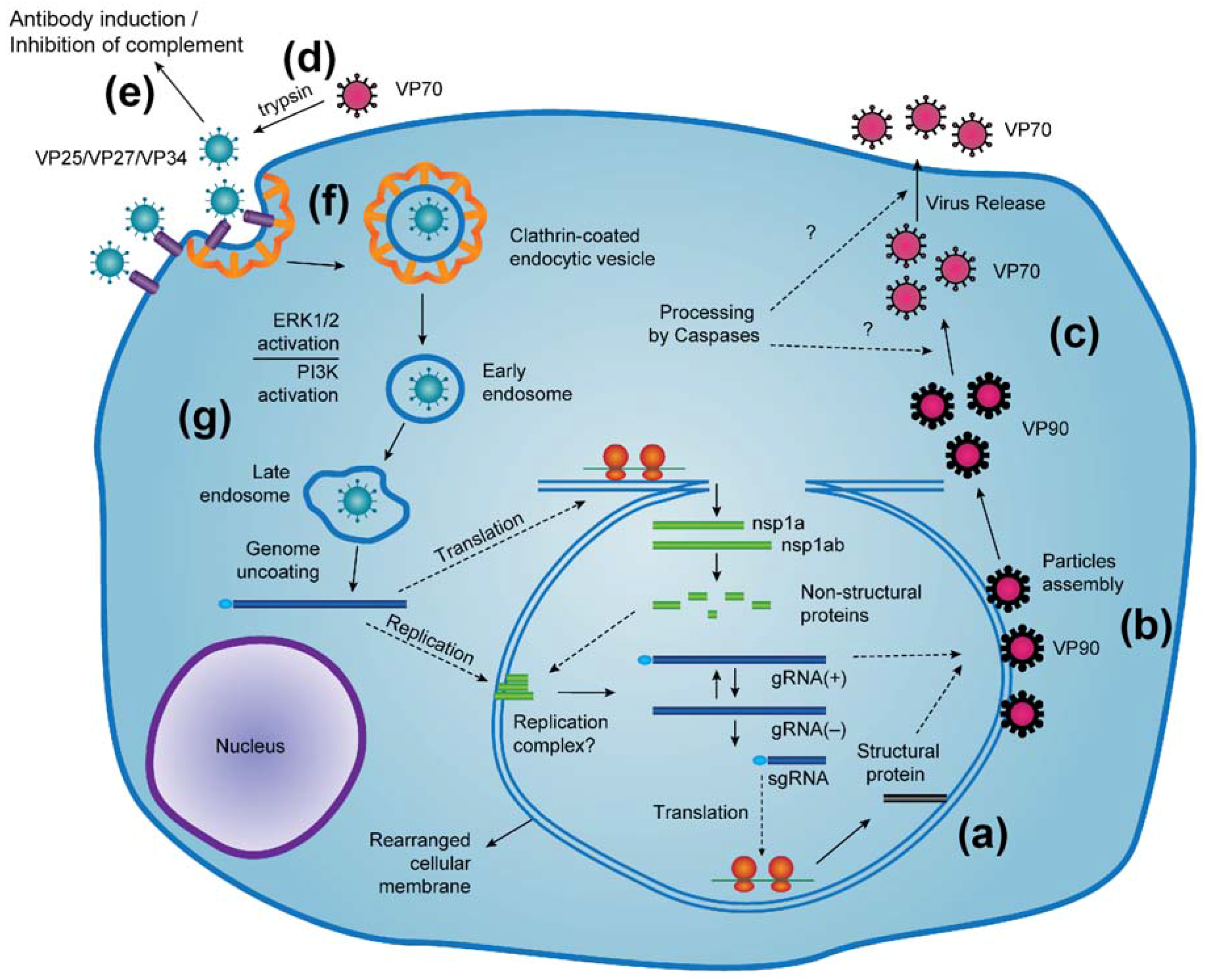
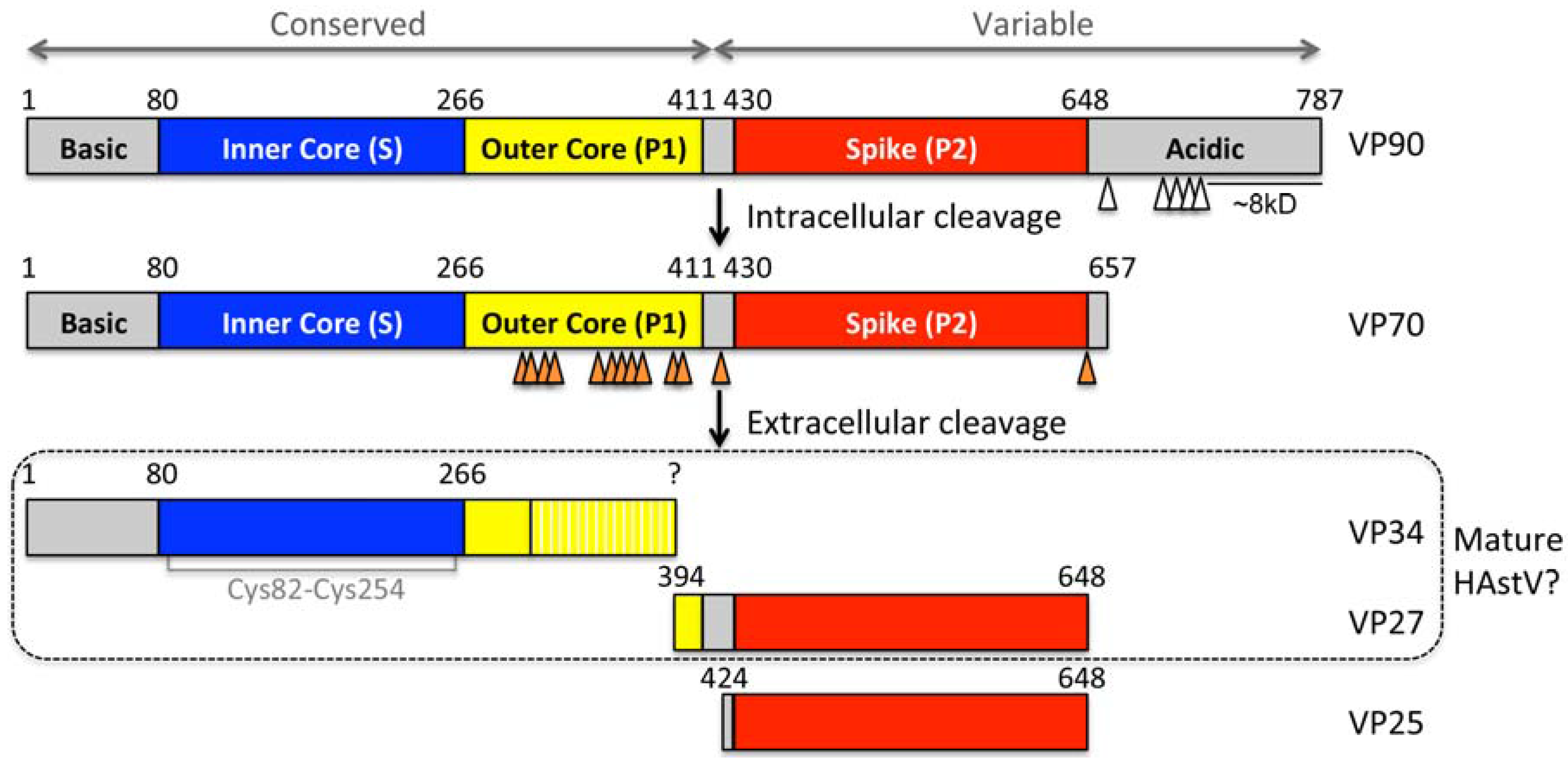
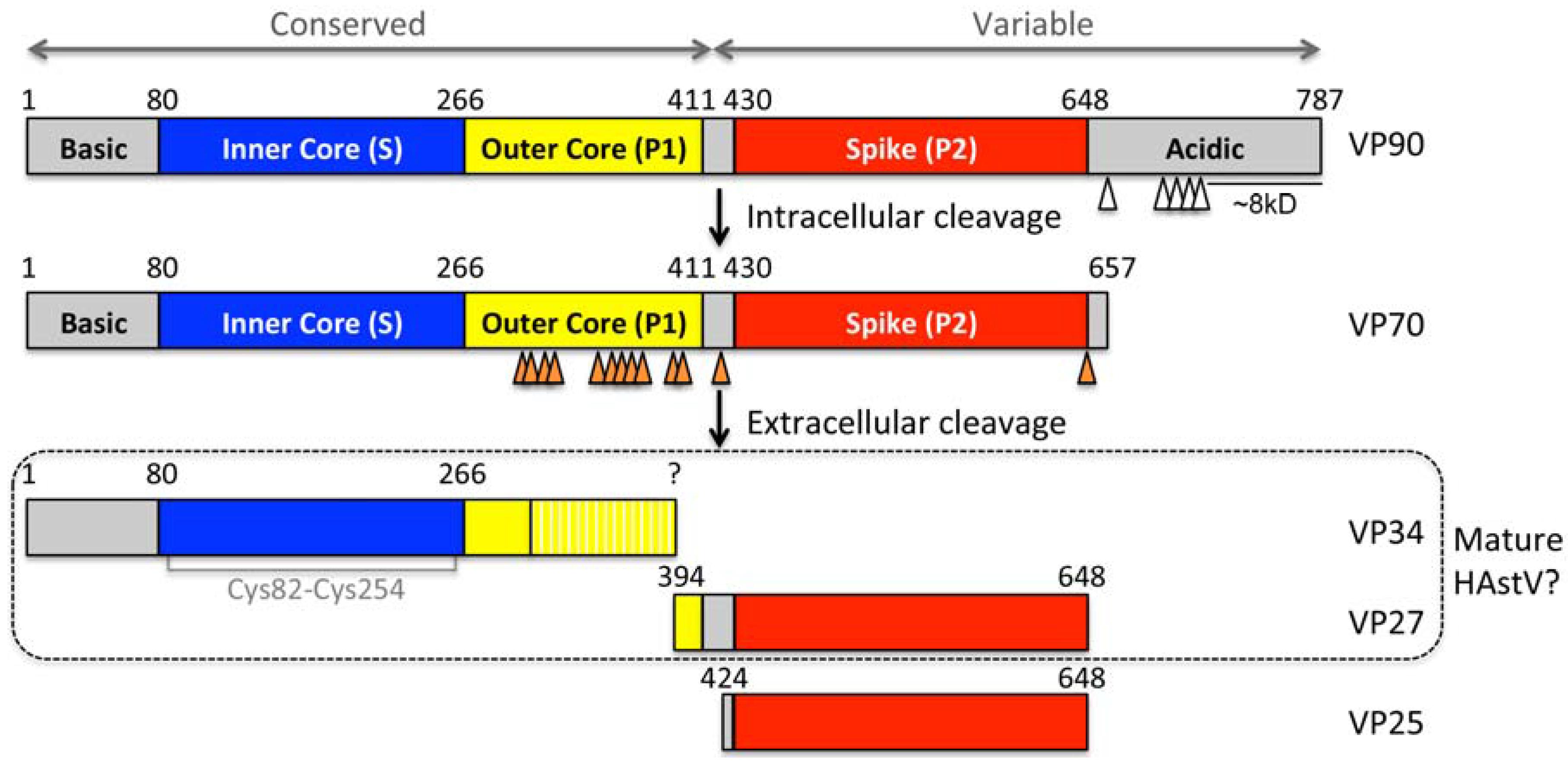
2. Capsid Assembly and Proteolytic Processing
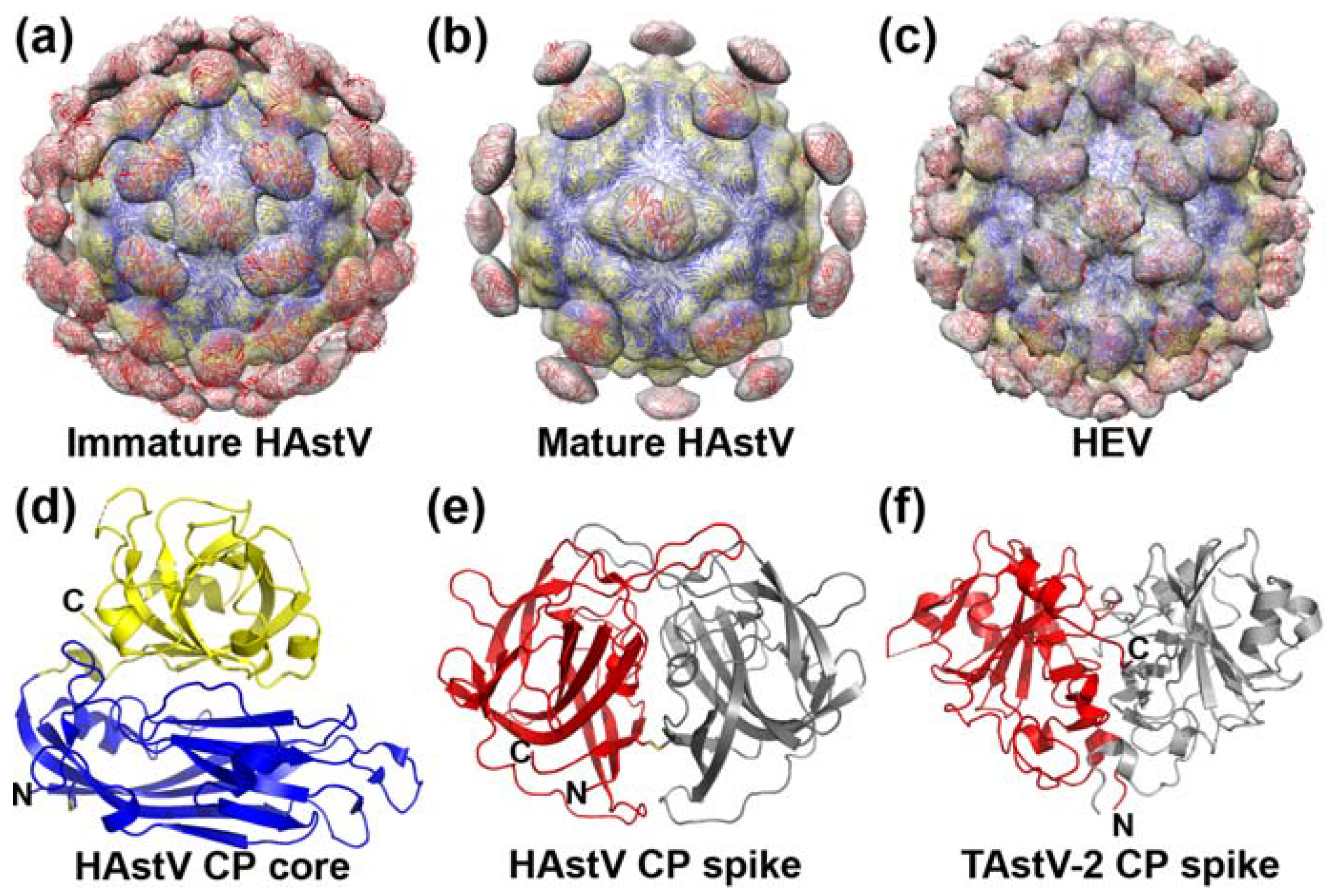
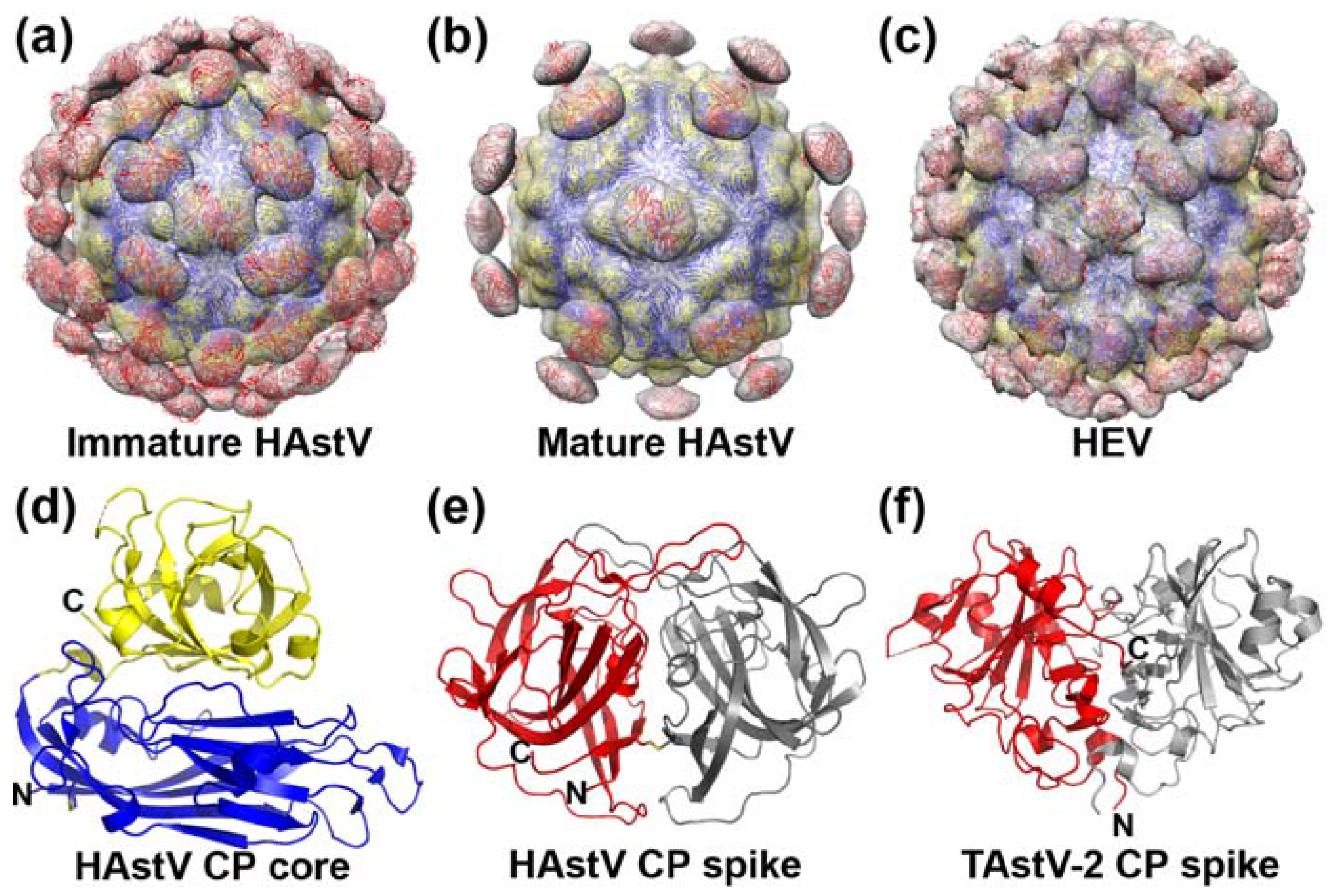
3. Capsid Structure
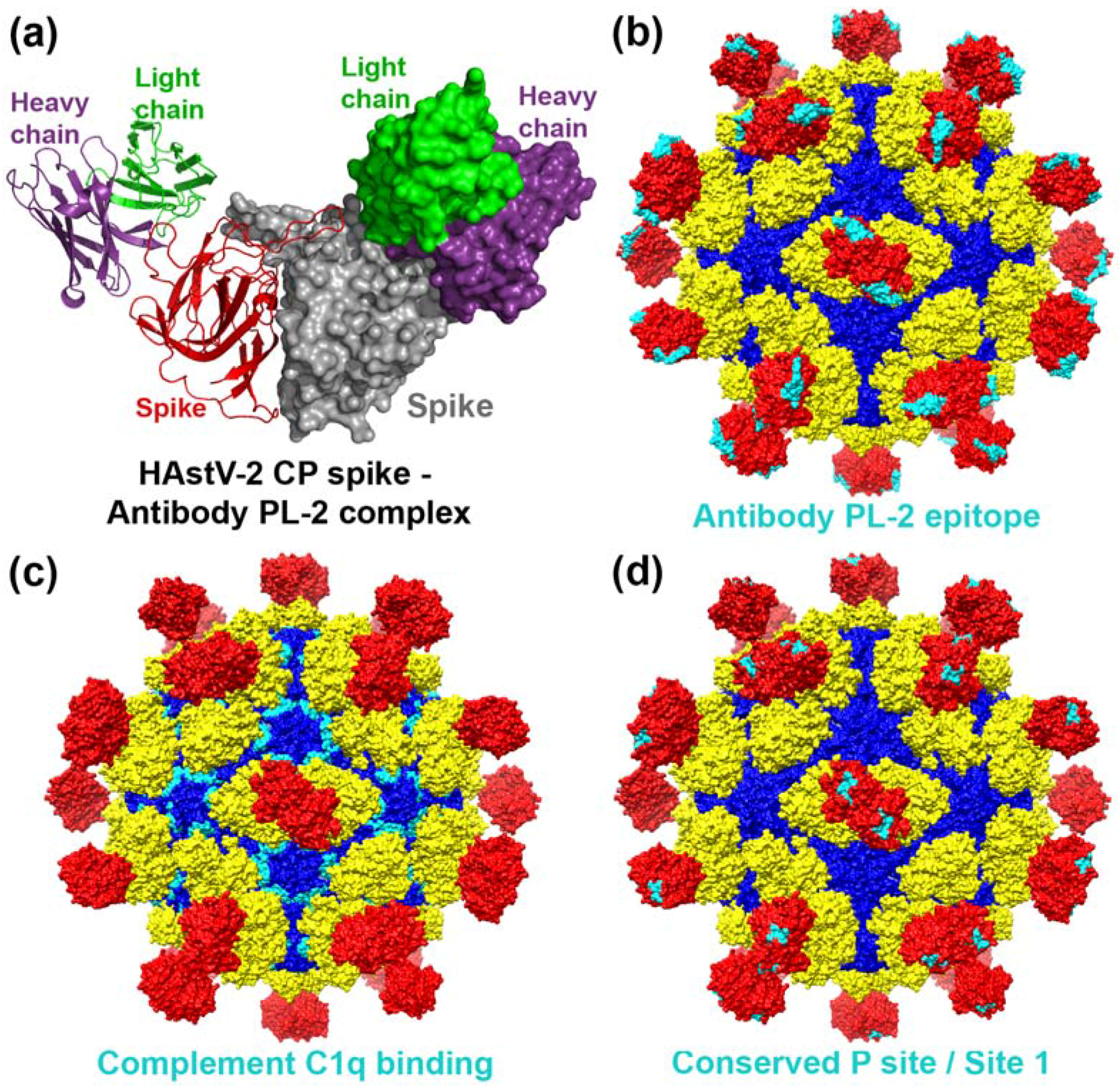
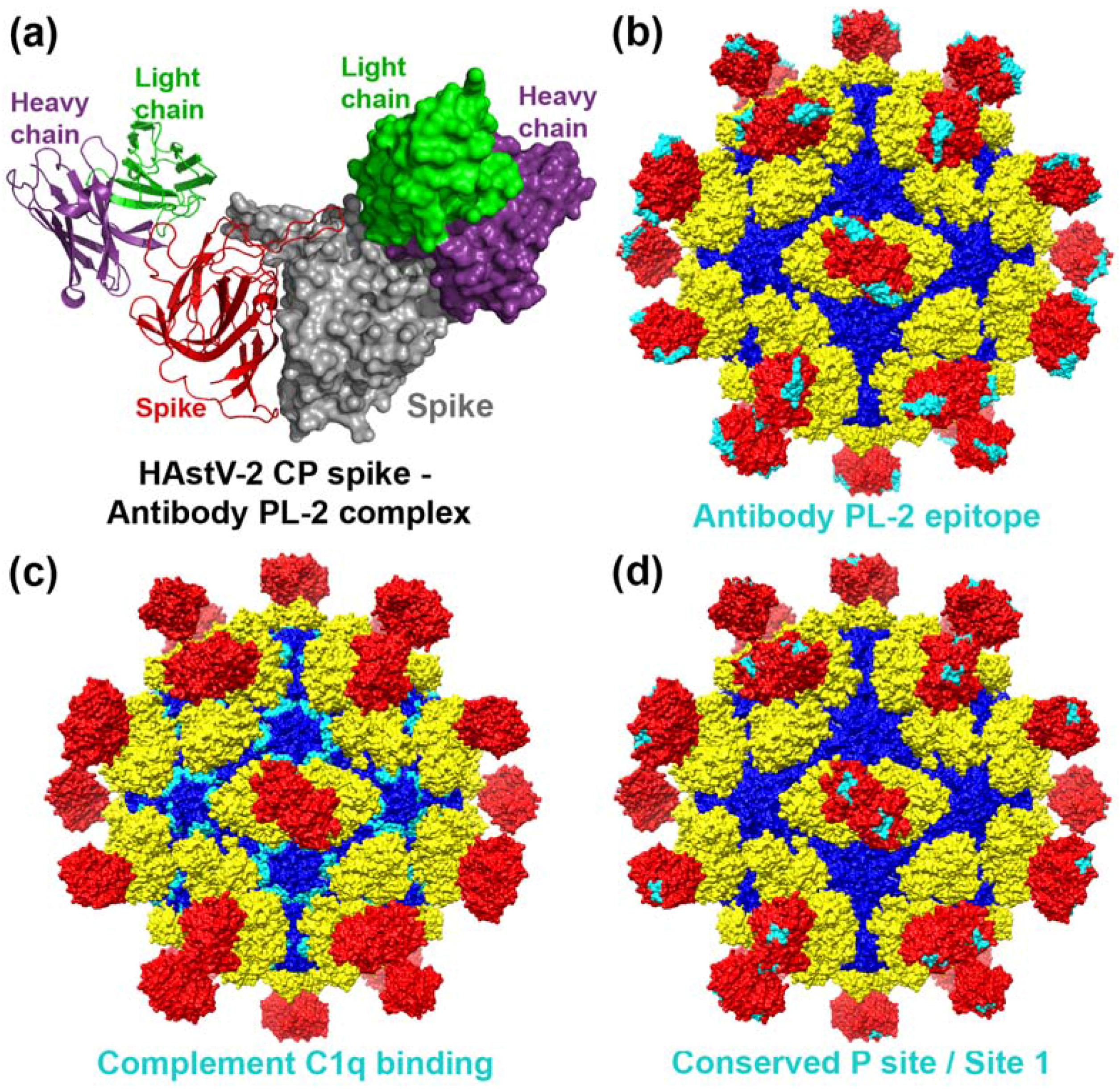
4. Capsid Interactions with Antibodies
5. Capsid Interactions with Complement
6. Virus Entry into Cells
7. Conclusions
- What is the role of the acidic region at the C-terminus of the CP?
- What is the mechanism of astrovirus release from the host cell?
- Which extracellular protease(s) play a role in HAstV maturation in vivo?
- What is the mechanism by which protease maturation increases HAstV infectivity?
- Which of the protease cleavage sites are essential for HAstV infectivity?
- Are the spike dimers homodimeric for VP27 or heterodimeric for VP25 and VP27?
- What are the structures of CPs from other sequence-divergent astroviruses?
- What are the high-resolution structures of the immature and mature astrovirus particles?
- Where do other neutralizing and non-neutralizing antibodies bind?
- What are the mechanisms by which neutralizing antibodies block infection?
- Do antibodies protect from astrovirus infection in vivo?
- What is the role of CP inhibition of complement activation in astrovirus pathogenesis?
- What is the astrovirus host cell receptor and do astroviruses use co-receptors?
- Do glycans play a role in virus cell entry?
- Where are the receptor binding site(s) on the CP surface?
- What viral factors trigger activation of the ERK1/2 and PI3K pathways and how do they facilitate virus entry and replication?
- How and where is the astrovirus genome released into the cytoplasm?
- How does the CP modulate tight junctions and intestinal barrier permeability?
Author Contributions
Conflicts of Interest
References
- Mendez, E.; Arias, C.F. Astroviruses. In Fields Virology, 6th ed.; Lippincott Williams & Wilkins: New York, NY, USA, 2013; Volume 1, pp. 609–628. [Google Scholar]
- Schultz-Cherry, S. Astrovirus Research: Essential Ideas, Everyday Impacts, Future Directions; Springer: New York, NY, USA, 2013; pp. XIV, 185. [Google Scholar]
- Lewis, T.L.; Greenberg, H.B.; Herrmann, J.E.; Smith, L.S.; Matsui, S.M. Analysis of astrovirus serotype 1 RNA, identification of the viral RNA-dependent RNA polymerase motif, and expression of a viral structural protein. J. Virol. 1994, 68, 77–83. [Google Scholar] [PubMed]
- Monroe, S.S.; Jiang, B.; Stine, S.E.; Koopmans, M.; Glass, R.I. Subgenomic RNA sequence of human astrovirus supports classification of astroviridae as a new family of RNA viruses. J. Virol. 1993, 67, 3611–3614. [Google Scholar] [PubMed]
- Monroe, S.S.; Stine, S.E.; Gorelkin, L.; Herrmann, J.E.; Blacklow, N.R.; Glass, R.I. Temporal synthesis of proteins and RNAs during human astrovirus infection of cultured cells. J. Virol. 1991, 65, 641–648. [Google Scholar] [PubMed]
- Willcocks, M.M.; Carter, M.J. Identification and sequence determination of the capsid protein gene of human astrovirus serotype 1. FEMS Microbiol. Lett. 1993, 114, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Geigenmuller, U.; Ginzton, N.H.; Matsui, S.M. Studies on intracellular processing of the capsid protein of human astrovirus serotype 1 in infected cells. J. Gen. Virol. 2002, 83, 1691–1695. [Google Scholar] [CrossRef] [PubMed]
- Geigenmuller, U.; Chew, T.; Ginzton, N.; Matsui, S.M. Processing of nonstructural protein 1a of human astrovirus. J. Virol. 2002, 76, 2003–2008. [Google Scholar] [CrossRef] [PubMed]
- Mendez, E.; Aguirre-Crespo, G.; Zavala, G.; Arias, C.F. Association of the astrovirus structural protein VP90 with membranes plays a role in virus morphogenesis. J. Virol. 2007, 81, 10649–10658. [Google Scholar] [CrossRef] [PubMed]
- Banos-Lara Mdel, R.; Mendez, E. Role of individual caspases induced by astrovirus on the processing of its structural protein and its release from the cell through a non-lytic mechanism. Virology 2010, 401, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Mendez, E.; Fernandez-Luna, T.; Lopez, S.; Mendez-Toss, M.; Arias, C.F. Proteolytic processing of a serotype 8 human astrovirus ORF2 polyprotein. J. Virol. 2002, 76, 7996–8002. [Google Scholar] [CrossRef] [PubMed]
- Mendez, E.; Salas-Ocampo, E.; Arias, C.F. Caspases mediate processing of the capsid precursor and cell release of human astroviruses. J. Virol. 2004, 78, 8601–8608. [Google Scholar] [CrossRef] [PubMed]
- McGuffin, L.J.; Bryson, K.; Jones, D.T. The psipred protein structure prediction server. Bioinformatics 2000, 16, 404–405. [Google Scholar] [CrossRef] [PubMed]
- Bass, D.M.; Qiu, S. Proteolytic processing of the astrovirus capsid. J. Virol. 2000, 74, 1810–1814. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.W.; Kurtz, J.B. Serial propagation of astrovirus in tissue culture with the aid of trypsin. J. Gen. Virol. 1981, 57, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Fauquier, A.; Carrascosa, A.L.; Carrascosa, J.L.; Otero, A.; Glass, R.I.; Lopez, J.A.; San Martin, C.; Melero, J.A. Characterization of a human astrovirus serotype 2 structural protein (VP26) that contains an epitope involved in virus neutralization. Virology 1994, 201, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Krishna, N.K. Identification of structural domains involved in astrovirus capsid biology. Viral Immunol. 2005, 18, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Appleton, H.; Higgins, P.G. Letter: Viruses and gastroenteritis in infants. Lancet 1975, 1, 1297. [Google Scholar] [CrossRef]
- Madeley, C.R.; Cosgrove, B.P. Letter: 28 nm particles in faeces in infantile gastroenteritis. Lancet 1975, 2, 451–452. [Google Scholar] [CrossRef]
- Risco, C.; Carrascosa, J.L.; Pedregosa, A.M.; Humphrey, C.D.; Sanchez-Fauquier, A. Ultrastructure of human astrovirus serotype 2. J. Gen. Virol. 1995, 76 (Pt 8), 2075–2080. [Google Scholar] [CrossRef] [PubMed]
- Dryden, K.A.; Tihova, M.; Nowotny, N.; Matsui, S.M.; Mendez, E.; Yeager, M. Immature and mature human astrovirus: Structure, conformational changes, and similarities to hepatitis e virus. J. Mol. Biol. 2012, 422, 650–658. [Google Scholar] [CrossRef] [PubMed]
- Bass, D.M.; Upadhyayula, U. Characterization of human serotype 1 astrovirus-neutralizing epitopes. J. Virol. 1997, 71, 8666–8671. [Google Scholar] [PubMed]
- Willcocks, M.M.; Carter, M.J.; Laidler, F.R.; Madeley, C.R. Growth and characterisation of human faecal astrovirus in a continuous cell line. Arch. Virol. 1990, 113, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Li, T.C.; Mayazaki, N.; Simon, M.N.; Wall, J.S.; Moore, M.; Wang, C.Y.; Takeda, N.; Wakita, T.; Miyamura, T.; et al. Structure of hepatitis e virion-sized particle reveals an RNA-dependent viral assembly pathway. J. Biol. Chem. 2010, 285, 33175–33183. [Google Scholar] [CrossRef] [PubMed]
- York, R.L.; Yousefi, P.A.; Bogdanoff, W.; Haile, S.; Tripathi, S.; DuBois, R.M. Structural, mechanistic, and antigenic characterization of the human astrovirus capsid. J. Virol. 2016, 90, 2254–2263. [Google Scholar] [CrossRef] [PubMed]
- DuBois, R.M.; Freiden, P.; Marvin, S.; Reddivari, M.; Heath, R.J.; White, S.W.; Schultz-Cherry, S. Crystal structure of the avian astrovirus capsid spike. J. Virol. 2013, 87, 7853–7863. [Google Scholar] [CrossRef] [PubMed]
- Toh, Y.; Harper, J.; Dryden, K.A.; Yeager, M.; Arias, C.F.; Mendez, E.; Tao, Y.J. Crystal structure of the human astrovirus capsid protein. J. Virol. 2016, 90, 9008–9017. [Google Scholar] [CrossRef] [PubMed]
- Bogdanoff, W.A.; Campos, J.; Perez, E.I.; Yin, L.; Alexander, D.L.; DuBois, R.M. Structure of a human astrovirus capsid—Antibody complex and mechanistic insights into virus neutralization. J. Virol. 2016, 91, e01859-16. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Dong, L.; Mendez, E.; Tao, Y. Crystal structure of the human astrovirus capsid spike. Proc. Natl. Acad. Sci. USA 2011, 108, 12681–12686. [Google Scholar] [CrossRef] [PubMed]
- Guu, T.S.; Liu, Z.; Ye, Q.; Mata, D.A.; Li, K.; Yin, C.; Zhang, J.; Tao, Y.J. Structure of the hepatitis e virus-like particle suggests mechanisms for virus assembly and receptor binding. Proc. Natl. Acad. Sci. USA 2009, 106, 12992–12997. [Google Scholar] [CrossRef] [PubMed]
- Burbelo, P.D.; Ching, K.H.; Esper, F.; Iadarola, M.J.; Delwart, E.; Lipkin, W.I.; Kapoor, A. Serological studies confirm the novel astrovirus HMOAstV-C as a highly prevalent human infectious agent. PLoS ONE 2011, 6, e22576. [Google Scholar] [CrossRef] [PubMed]
- Cubitt, W.D.; Mitchell, D.K.; Carter, M.J.; Willcocks, M.M.; Holzel, H. Application of electronmicroscopy, enzyme immunoassay, and RT-PCR to monitor an outbreak of astrovirus type 1 in a paediatric bone marrow transplant unit. J. Med. Virol. 1999, 57, 313–321. [Google Scholar] [CrossRef]
- Koopmans, M.P.; Bijen, M.H.; Monroe, S.S.; Vinje, J. Age-stratified seroprevalence of neutralizing antibodies to astrovirus types 1 to 7 in humans in the netherlands. Clin. Diagn. Lab. Immunol. 1998, 5, 33–37. [Google Scholar] [PubMed]
- Kurtz, J.; Lee, T. Astrovirus gastroenteritis age distribution of antibody. Med. Microbiol. Immunol. 1978, 166, 227–230. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, J.B.; Lee, T.W.; Craig, J.W.; Reed, S.E. Astrovirus infection in volunteers. J. Med. Virol. 1979, 3, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Noel, J.; Cubitt, D. Identification of astrovirus serotypes from children treated at the hospitals for sick children, London 1981–93. Epidemiol. Infect. 1994, 113, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.K.; Matson, D.O.; Cubitt, W.D.; Jackson, L.J.; Willcocks, M.M.; Pickering, L.K.; Carter, M.J. Prevalence of antibodies to astrovirus types 1 and 3 in children and adolescents in norfolk, virginia. Pediatr. Infect. Dis. J. 1999, 18, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.K. Astrovirus gastroenteritis. Pediatr. Infect. Dis. J. 2002, 21, 1067–1069. [Google Scholar] [CrossRef] [PubMed]
- Bjorkholm, M.; Celsing, F.; Runarsson, G.; Waldenstrom, J. Successful intravenous immunoglobulin therapy for severe and persistent astrovirus gastroenteritis after fludarabine treatment in a patient with waldenstrom’s macroglobulinemia. Int. J. Hematol. 1995, 62, 117–120. [Google Scholar] [CrossRef]
- Marvin, S.A.; Huerta, C.T.; Sharp, B.; Freiden, P.; Cline, T.D.; Schultz-Cherry, S. Type I interferon response limits astrovirus replication and protects against increased barrier permeability in vitro and in vivo. J. Virol. 2015, 90, 1988–1996. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, J.E.; Hudson, R.W.; Perron-Henry, D.M.; Kurtz, J.B.; Blacklow, N.R. Antigenic characterization of cell-cultivated astrovirus serotypes and development of astrovirus-specific monoclonal antibodies. J. Infect. Dis. 1988, 158, 182–185. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, J.B.; Lee, T.W. Human astrovirus serotypes. Lancet 1984, 2, 1405. [Google Scholar] [CrossRef]
- Lee, T.W.; Kurtz, J.B. Human astrovirus serotypes. J. Hyg. 1982, 89, 539–540. [Google Scholar] [CrossRef] [PubMed]
- Bogdanoff, W.A.; Morgenstern, D.; Bern, M.; Ueberheide, B.M.; Sanchez-Fauquier, A.; DuBois, R.M. De novo sequencing and resurrection of a human astrovirus-neutralizing antibody. ACS Infect. Dis. 2016, 2, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Gronemus, J.Q.; Hair, P.S.; Crawford, K.B.; Nyalwidhe, J.O.; Cunnion, K.M.; Krishna, N.K. Potent inhibition of the classical pathway of complement by a novel C1q-binding peptide derived from the human astrovirus coat protein. Mol. Immunol. 2010, 48, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Koci, M.D.; Moser, L.A.; Kelley, L.A.; Larsen, D.; Brown, C.C.; Schultz-Cherry, S. Astrovirus induces diarrhea in the absence of inflammation and cell death. J. Virol. 2003, 77, 11798–11808. [Google Scholar] [CrossRef] [PubMed]
- Sebire, N.J.; Malone, M.; Shah, N.; Anderson, G.; Gaspar, H.B.; Cubitt, W.D. Pathology of astrovirus associated diarrhoea in a paediatric bone marrow transplant recipient. J. Clin. Pathol. 2004, 57, 1001–1003. [Google Scholar] [CrossRef] [PubMed]
- Bonaparte, R.S.; Hair, P.S.; Banthia, D.; Marshall, D.M.; Cunnion, K.M.; Krishna, N.K. Human astrovirus coat protein inhibits serum complement activation via C1, the first component of the classical pathway. J. Virol. 2008, 82, 817–827. [Google Scholar] [CrossRef] [PubMed]
- Hair, P.S.; Gronemus, J.Q.; Crawford, K.B.; Salvi, V.P.; Cunnion, K.M.; Thielens, N.M.; Arlaud, G.J.; Rawal, N.; Krishna, N.K. Human astrovirus coat protein binds C1q and MBL and inhibits the classical and lectin pathways of complement activation. Mol. Immunol. 2010, 47, 792–798. [Google Scholar] [CrossRef] [PubMed]
- Lopez, S.; Arias, C.F. Multistep entry of rotavirus into cells: A versaillesque dance. Trends Microbiol. 2004, 12, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Triantafilou, K.; Triantafilou, M. Lipid raft microdomains: Key sites for coxsackievirus A9 infectious cycle. Virology 2003, 317, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Wu, E.; Nemerow, G.R. Virus yoga: The role of flexibility in virus host cell recognition. Trends Microbiol. 2004, 12, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Brinker, J.P.; Blacklow, N.R.; Herrmann, J.E. Human astrovirus isolation and propagation in multiple cell lines. Arch. Virol. 2000, 145, 1847–1856. [Google Scholar] [CrossRef] [PubMed]
- Geigenmuller, U.; Ginzton, N.H.; Matsui, S.M. Construction of a genome-length cdna clone for human astrovirus serotype 1 and synthesis of infectious RNA transcripts. J. Virol. 1997, 71, 1713–1717. [Google Scholar] [PubMed]
- Casorla, L.; López, T.; Parrón, J.A.; Sánchez, L.; Arias, C.F. Human astrovirus infection is not affected by bovine milk fractions. Unpublished.
- Funtional Glycomics Gateway. Available online: http://www.functionalglycomics.org (accessed on 17 January 2017).
- Mendez, E.; Munoz-Yanez, C.; Sanchez-San Martin, C.; Aguirre-Crespo, G.; Banos-Lara Mdel, R.; Gutierrez, M.; Espinosa, R.; Acevedo, Y.; Arias, C.F.; Lopez, S. Characterization of human astrovirus cell entry. J. Virol. 2014, 88, 2452–2460. [Google Scholar] [CrossRef] [PubMed]
- Donelli, G.; Superti, F.; Tinari, A.; Marziano, M.L. Mechanism of astrovirus entry into graham 293 cells. J. Med. Virol. 1992, 38, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Moser, L.A.; Schultz-Cherry, S. Suppression of astrovirus replication by an erk1/2 inhibitor. J. Virol. 2008, 82, 7475–7482. [Google Scholar] [CrossRef] [PubMed]
- Tange, S.; Zhou, Y.; Nagakui-Noguchi, Y.; Imai, T.; Nakanishi, A. Initiation of human astrovirus type 1 infection was blocked by inhibitors of phosphoinositide 3-kinase. Virol. J. 2013, 10, 153. [Google Scholar] [CrossRef] [PubMed]
- Caballero, S.; Guix, S.; Ribes, E.; Bosch, A.; Pinto, R.M. Structural requirements of astrovirus virus-like particles assembled in insect cells. J. Virol. 2004, 78, 13285–13292. [Google Scholar] [CrossRef] [PubMed]
- Moser, L.A.; Carter, M.; Schultz-Cherry, S. Astrovirus increases epithelial barrier permeability independently of viral replication. J. Virol. 2007, 81, 11937–11945. [Google Scholar] [CrossRef] [PubMed]
- Meliopoulos, V.A.; Marvin, S.A.; Freiden, P.; Moser, L.A.; Nighot, P.; Ali, R.; Blikslager, A.; Reddivari, M.; Heath, R.J.; Koci, M.D.; et al. Oral administration of astrovirus capsid protein is sufficient to induce acute diarrhea in vivo. mBio 2016, 7, e01494-16. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arias, C.F.; DuBois, R.M. The Astrovirus Capsid: A Review. Viruses 2017, 9, 15. https://doi.org/10.3390/v9010015
Arias CF, DuBois RM. The Astrovirus Capsid: A Review. Viruses. 2017; 9(1):15. https://doi.org/10.3390/v9010015
Chicago/Turabian StyleArias, Carlos F., and Rebecca M. DuBois. 2017. "The Astrovirus Capsid: A Review" Viruses 9, no. 1: 15. https://doi.org/10.3390/v9010015
APA StyleArias, C. F., & DuBois, R. M. (2017). The Astrovirus Capsid: A Review. Viruses, 9(1), 15. https://doi.org/10.3390/v9010015