
HTLV-1, Immune Response and Autoimmunity


{kind=link}
Abstract
:1. Introduction
2. Human T-lymphotropic Virus Type-1 (HTLV-1)
3. Immunological Changes in HTLV-1-Infected Patients
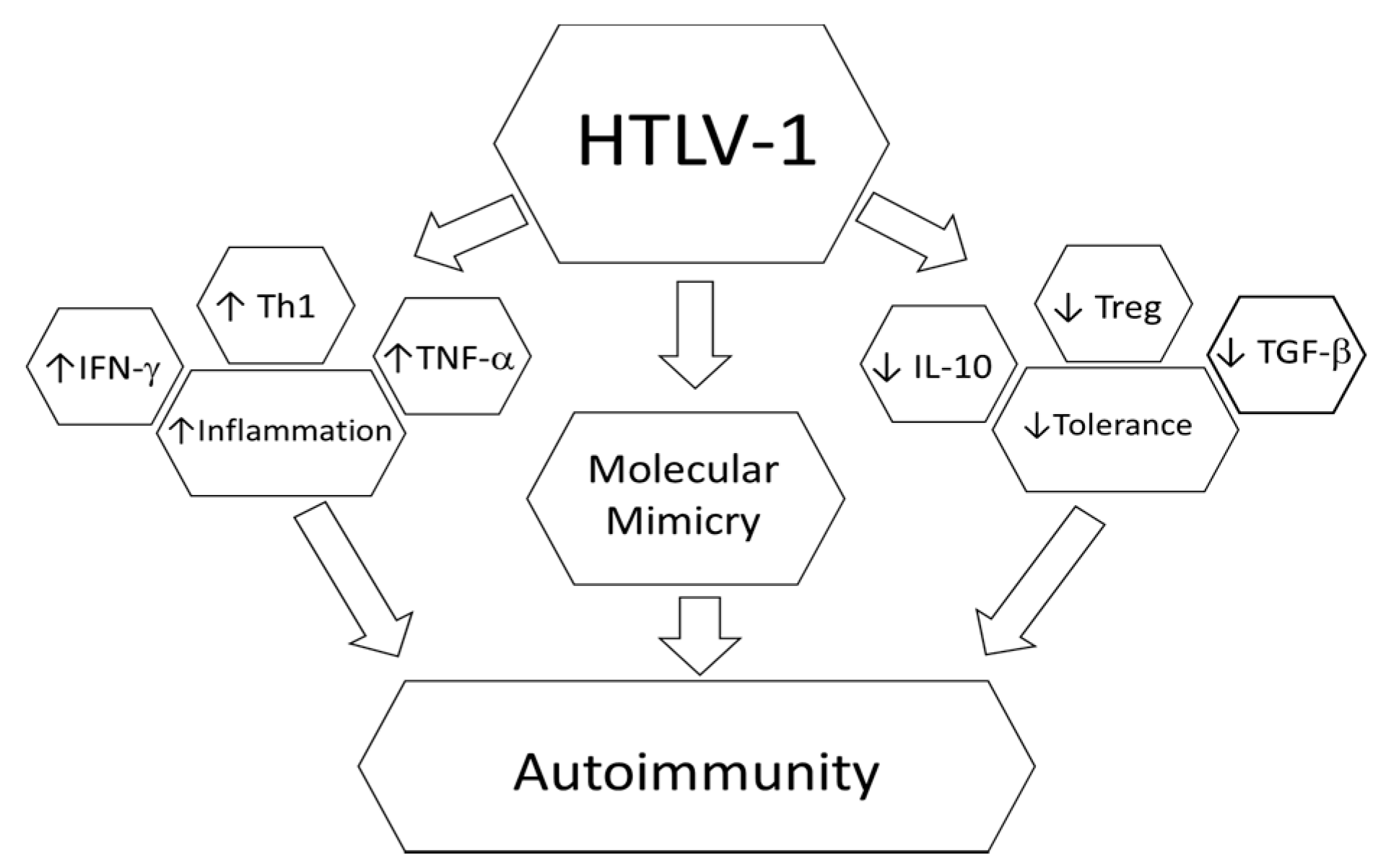
4. HTLV-1 and Autoimmunity
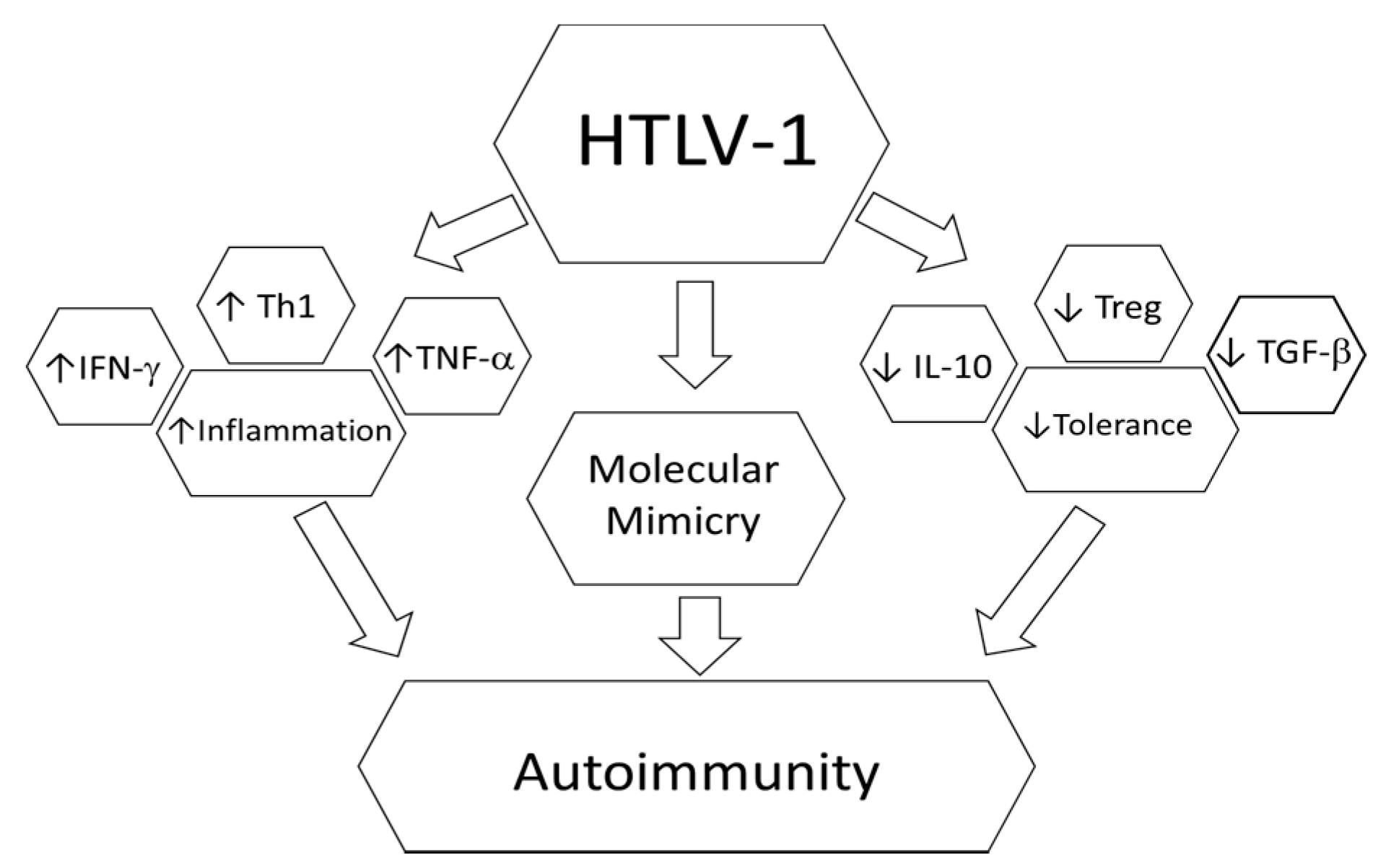
5. Rheumatoid Arthritis
6. Sjögren’s Syndrome
7. Systemic Lupus Erythematosus
8. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Von Herrath, M.G.; Fujinami, R.S.; Whitton, J.L. Microorganisms and autoimmunity: Making the barren field fertile? Nat. Rev. Microbiol. 2003, 1, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Libbey, J.E.; Fujinami, R.S. Potential triggers of MS. Results Probl. Cell Differ. 2010, 51, 21–42. [Google Scholar]
- Fujinami, R.S. Viruses and autoimmune disease—Two sides of the same coin? Trends Microbiol. 2001, 9, 377–381. [Google Scholar] [CrossRef]
- McCoy, L.; Tsunoda, I.; Fujinami, R.S. Multiple sclerosis and virus induced immune responses: Autoimmunity can be primed by molecular mimicry and augmented by bystander activation. Autoimmunity 2006, 39, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Sfriso, P.; Ghirardello, A.; Botsios, C.; Tonon, M.; Zen, M.; Bassi, N.; Bassetto, F.; Doria, A. Infections and autoimmunity: The multifaceted relationship. J. Leukoc. Biol. 2010, 87, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Araujo, A.Q.; Silva, S.T. The HTLV-1 neurological complex. Lancet Neurol. 2006, 5, 1068–1076. [Google Scholar] [CrossRef]
- Gessain, A.; Barin, F.; Vernant, J.C.; Gout, O.; Maurs, L.; Calender, A.; de Thé, G. Antibodies to human T lymphotropicvírus type I in patients with tropical spastic paraparesis. Lancet 1985, 2, 407–410. [Google Scholar] [CrossRef]
- Román, G.C.; Osame, M. Identity of HTLV-I-associated tropical spastic paraparesis and HTLV-I-associated myelopathy. Lancet 1988, 1. [Google Scholar] [CrossRef]
- Romanelli, L.C.F.; Caramelli, P.; Proietti, A.B.F.C. Human T-cell leukemia virus type 1 (HTLV-1): When do we suspect of infection? Rev. Assoc. Med. Brás. 2010, 56, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Cooper, S.A.; van der Loeff, M.S.; Taylor, G.P. The neurology of HTLV-1 infection. Pract. Neurol. 2009, 9, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Gessain, A.; Mahieux, R. Tropical spastic paraparesis and HTLV-1 associated myelopathy: Clinical, epidemiological, virological and therapeutic aspects. Rev. Neurol. 2012, 168, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Boxus, M.; Willems, L. Mechanisms of HTLV-1 persistence and transformation. Br. J. Cancer 2009, 101, 1497–1501. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, H. Cellular factors involved in HTLV-1 entry and pathogenicit. Front. Microbiol. 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Lairmore, M.D.; Anupam, R.; Bowden, N.; Haines, R.; Haynes, R.A., II; Ratner, L.; Green, P.L. Molecular determinants of human T-lymphotropic virus type 1 transmission and spread. Viruses 2011, 3, 1131–1165. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, E.; Yamano, Y.; Jacobson, S. Neuroimmunity of HTLV-I Infection. J. Neuroimmune Pharmacol. 2010, 5, 310–325. [Google Scholar] [CrossRef] [PubMed]
- Best, I.; López, G.; Verdonck, K.; González, E.; Tipismana, M.; Gotuzzo, E.; Vanham, G.; Clark, D. IFN-γ production in response to Tax 161-233, and frequency of CD4+ Foxp3+ and Lin HLA-DRhigh CD123+ cells, discriminate HAM/TSP patients from asymptomatic HTLV-1-carriers in a Peruvian population. Immunology 2009, 128, e777–e786. [Google Scholar] [CrossRef] [PubMed]
- Marriott, S.J.; Semmes, O.J. Impact of HTLV-I Tax on cell cycle progression and the cellular DNA damage repair response. Oncogene 2005, 24, 5986–5995. [Google Scholar] [CrossRef] [PubMed]
- Grassmann, R.; Aboud, M.; Jeang, K.T. Molecular mechanisms of cellular transformation by HTLV-1 Tax. Oncogene 2005, 24, 5976–5985. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Ren, T.; Sun, S.C. New insight into the oncogenic mechanism of the retroviral oncoprotein Tax. Protein Cell 2012, 3, 581–589. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Higuchi, M.; Fujii, M. Distinct functions of HTLV-1 Tax1 from HTLV-2 Tax2 contribute key roles to viral pathogenesis. Retrovirology 2009, 6. [Google Scholar] [CrossRef] [PubMed]
- Jeang, K.T. HTLV-1 and adult T-cell leukemia: Insights into viral transformation of cells 30 years after virus discovery. J. Formos. Med. Assoc. 2010, 109, 688–693. [Google Scholar] [CrossRef]
- Beimling, P.; Moelling, K. Direct interaction of CREB protein with 21 bp Tax-response elements of HTLV-ILTR. Oncogene 1992, 7, 257–262. [Google Scholar] [PubMed]
- Satou, Y.; Yasunaga, J.; Yoshida, M.; Matsuoka, M. HTLV-I basic leucine zipper factor gene mRNA supports proliferation of adult T cell leukemia cells. Proc. Natl. Acad. Sci. USA 2006, 103, 720–725. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, M. Human T-cell leukemia virus type I (HTLV-I) infection and the onset of adult T-cell leukemia (ATL). Retrovirology 2005, 2. [Google Scholar] [CrossRef] [PubMed]
- Cabral, F.; Arruda, L.B.; de Araújo, M.L.; Montanheiro, P.; Smid, J.; de Oliveira, A.C.; Duarte, A.J.; Casseb, J. Detection of human T-cell lymphotropic virus type 1 in plasma samples. Virus Res. 2012, 163, 87–90. [Google Scholar] [CrossRef] [PubMed]
- Journo, C.; Mahieux, R. HTLV-1 and innate immunity. Viruses 2011, 3, 1374–1394. [Google Scholar] [CrossRef] [PubMed]
- Nagai, M.; Yamano, Y.; Brennan, M.B.; Mora, C.A.; Jacobson, S. Increased HTLV-I proviral load and preferential expansion of HTLV-I Tax-specific CD8+ T cells in cerebrospinal fluid from patients with HAM/TSP. Ann. Neurol. 2001, 50, 807–812. [Google Scholar] [CrossRef] [PubMed]
- Gallo, R.C. Research and discovery of the first human cancer virus, HTLV-1. Best Pract. Res. Clin. Haematol. 2011, 24, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Morgan, O. HTLV-1 associated myelopathy/tropical spastic paraparesis: How far have we come? West Indian Med. J. 2011, 60, 505–512. [Google Scholar] [PubMed]
- Ahuja, J.; Lepoutre, V.; Wigdahl, B.; Khan, Z.K.; Jain, P. Induction of pro-inflammatory cytokines by human T-cell leukemia virus type-1 Tax protein as determined by multiplexed cytokine protein array analyses of human dendritic cells. Biomed. Pharmacother. 2007, 61, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Yamano, Y.; Araya, N.; Sato, T.; Utsunomiya, A.; Azakami, K.; Hasegawa, D.; Izumi, T.; Fujita, H.; Aratani, S.; Yagishita, N.; et al. Abnormally high levels of virus-infected INF-γ+CCR4+CD4+CD25+ T cells in a retrovirus-associated neuroinflammatory disorder. PLoS ONE 2009, 4. [Google Scholar] [CrossRef] [PubMed]
- Oliére, S.; Douville, R.; Sze, A.; Belgnaoui, S.M.; Hiscott, J. Modulation of innate immune responses during human T-cell leukemia virus (HTLV-1) pathogenesis. Cytokine Growth Factor Rev. 2011, 22, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Puccioni-Sohler, M.; Gasparetto, E.; Cabral-Castro, M.J.; Slatter, C.; Vidal, C.M.; Cortes, R.D.; Rosen, B.R.; Mainero, C. HAM/TSP: Association between white matter lesions on magnetic resonance imaging, clinical and cerebrospinal fluid findings. Arq. Neuropsiquiatr. 2012, 70, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Saito, M. Immunogenetics and the pathological mechanisms of human T-cell leukemia virustype 1-(HTLV-1-) associated myelopathy/tropical spastic paraparesis (HAM/TSP). Interdiscip. Perspect. Infect. Dis. 2010, 2010. [Google Scholar] [CrossRef]
- Shoeibi, A.; Etemadi, M.; Ahmadi, A.M.; Amini, M.; Boostani, R. “HTLV-I Infection” twenty-year research in neurology department of mashhad university of medical sciences. Iran. J. Basic Med. Sci. 2013, 16, 202–207. [Google Scholar] [PubMed]
- Satou, Y.; Matsuoka, M. HTLV-1 and the host immune system: How the virus disrupts immune regulation, leading to HTLV-1 associated diseases. J. Clin. Exp. Hematop. 2010, 50, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Fuzii, H.T.; Dias, G.A.S.; Barros, R.J.; Falcão, L.F.; Quaresma, J.A. Immunopathogenesis of HTLV-1-assoaciated myelopathy/tropical spastic paraparesis (HAM/TSP). Life Sci. 2014, 104, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Azran, I.; Schavinsky-Khrapunsky, Y.; Aboud, M. Role of Tax protein in human T-cell leukemia virus type-I leukemogenicity. Retrovirology 2004, 1. [Google Scholar] [CrossRef] [PubMed]
- Kibler, K.V.; Jeang, K.T. CREB/ATF-Dependent repression of cyclin A by human T-cell leukemia virus Type 1 Tax protein. J. Virol. 2001, 75, 2161–2173. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, J.; Ohshima, T.; Isono, O.; Shimotohno, K. HTLV-1 HBZ suppresses AP-1 activity by impairing both the DNA-binding ability and the stability of c-Jun protein. Oncogene 2005, 24, 1001–1010. [Google Scholar] [CrossRef] [PubMed]
- Wildin, R.S.; Freitas, A. IPEX and FOXP3: Clinical and research perspectives. J. Autoimmun. 2005, 25, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Bacchetta, R.; Gambineri, E.; Roncarolo, M.G. Role of regulatory T cells and FOXP3 in human diseases. J. Allergy Clin. Immunol. 2007, 120, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Toulza, F.; Heaps, A.; Tanaka, Y.; Taylor, G.P.; Bangham, C.R. High frequency of CD4+FoxP3+ cells in HTLV-1 infection: Inverse correlation with HTLV-1-especific CTL response. Blood 2008, 111, 5047–5053. [Google Scholar] [CrossRef] [PubMed]
- Brito-Melo, G.E.; Peruhype-Magalhães, V.; Teixeira-Carvalho, A.; Barbosa-Stancioli, E.F.; Carneiro-Proietti, A.B.; Catalan-Soares, B.; Ribas, J.G.; Martins-Filho, O.A. IL-10 produced by CD4+ and CD8+ T cells emerge as a putative immunoregulatory mechanism to counterbalance the monocyte-derived TNF-α and guarantee asymptomatic clinical status during chronic HTLV-I infection. Clin. Exp. Immunol. 2007, 147, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Santos, S.B.; Muniz, A.L.; Carvalho, E.M. HTLV-1-associated myelopathy Immunopathogenesis. Gaz. Méd. Bahia 2009, 79, 11–17. [Google Scholar]
- Palmer, D.C.; Restifo, N.P. Suppressors of cytokine signaling (SOCS) in T cell differentiation, maturation, and function. Trends Immunol. 2009, 30, 592–602. [Google Scholar] [CrossRef] [PubMed]
- Tamiya, T.; Kashiwagi, I; Takahashi, R.; Yasukawa, H.; Yoshimura, A. Suppressors of cytokine signaling (SOCS) proteins and JAK/STAT pathways: Regulation of T-cell inflammation by SOCS1 and SOCS3. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 980–985. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, A.; Suzuki, M.; Sakaguchi, R.; Hanada, T.; Yasukawa, H. SOCS, inflammation, and autoimmunity. Front. Immunol. 2012, 12. [Google Scholar] [CrossRef] [PubMed]
- Nishiura, Y.; Nakamura, T.; Fukushima, N.; Moriuchi, R.; Katamine, S.; Eguchi, K. Increased mRNA expression of Th1-cytokine signaling molecules in patients with HTLV-I-associated myelopathy/tropical spastic paraparesis. Tohoku J. Exp. Med. 2004, 204, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Araújo, A.Q.; Leite, A.C.; Lima, M.A.; Silva, M.T. HTLV-1 and neurological conditions: When to suspect and when to order a diagnostic test for HTLV-1 infection? Arq. Neuropsiquiatr. 2009, 67, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Castro-Costa, C.M.; Araújo, A.Q.; Câmara, C.C.; Ferreira, A.S.; Santos, T.J.; Castro-Costa, S.B.; Alcântara, R.N.; Taylor, G.P. Pain in tropical spastic paraparesis/HTLV-I associated myelopathy patients. Arq. Neuropsiquiatr. 2009, 67, 866–870. [Google Scholar] [CrossRef] [PubMed]
- Heraud, J.M.; Merien, F.; Mortreux, F.; Mahieux, R.; Kazanji, M. Immunological changes and cytokine gene expression during primary infection with human T-cell leukaemia virus type 1 in squirrel monkeys (Saimiri sciureus). Virology 2007, 361, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Michaëlsson, J.; Barbosa, H.M.; Jordan, K.A.; Chapman, J.M.; Brunialti, M.K.; Neto, W.K.; Nukui, Y.; Sabino, E.C.; Chieia, M.A.; Oliveira, A.S.; et al. The frequency of CD127low expressing CD4+CD25high T regulatory cells is inversely correlated with human T lymphotrophic virus type-1 (HTLV-1) proviral load in HTLV-1-infection and HTLV-1-associated myelopathy/tropical spastic paraparesis. BMC Immunol. 2008, 9. [Google Scholar] [CrossRef] [PubMed]
- Montes, M.; Sanchez, C.; Verdonck, K.; Lake, J.E.; Gonzalez, E.; Lopez, G.; Terashima, A.; Nolan, T.; Lewis, D.E.; Gotuzzo, E.; et al. Regulatory T cell expansion in HTLV-1 and strongyloidiasis co-infection is associated with reduced IL-5 responses to Strongyloides stercoralis antigen. PLoS Negl. Trop. Dis. 2009, 3. [Google Scholar] [CrossRef] [PubMed]
- Santos, S.B.; Porto, A.F.; Muniz, A.L.; Jesus, A.R.; Magalhães, E.; Melo, A.; Dutra, W.O.; Gollob, K.J.; Carvalho, E.M. Exacerbated inflammatory cellular imune response characteristics of HAM/TSP is observed in a large proportionof HTLV-I asymptomatic carriers. BMC Infect. Dis. 2004, 4. [Google Scholar] [CrossRef]
- Brito-Melo, G.E.; Martins-Filho, O.A.; Carneiro-Proietti, A.B.; Catalan-Soares, B.; Ribas, J.G.; Thorum, G.W.; Barbosa-Stancioli, E.F. Phenotypic study of peripheral blood leucocytes in HTLV-I-infected individuals from Minas Gerais, Brazil. Scand. J. Immunol. 2002, 55, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Coutinho, R.J.; Grassi, M.F.; Korngold, A.B.; Olavarria, V.N.; Galvão-Castro, B.; Mascarenhas, R.E. Human T lymphotropic virus type 1 (HTLV-1) proviral load induces activation of T-lymphocytes in asymptomatic carriers. BMC Infect. Dis. 2014, 22. [Google Scholar] [CrossRef] [PubMed]
- Walker, L.S.; Abbas, A.K. The enemy within: Keeping self-reactive T cells at bay in the periphery. Nat. Rev. Immunol. 2002, 2, 11–19. [Google Scholar] [CrossRef] [PubMed]
- O’Garra, A.; Vieira, P. Regulatory T cells and mechanisms of immune system control. Nat. Med. 2004, 10, 801–805. [Google Scholar] [CrossRef] [PubMed]
- Moudgil, K.D.; Choubey, D. Cytokines in autoimmunity: Role in induction, regulation, and treatment. J. Interferon Cytokine Res. 2011, 31, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Parks, C.G.; Miller, F.W.; Pollard, K.M.; Selmi, C.; Germolec, D.; Joyce, K.; Rose, N.R.; Humble, M.C. Expert panel workshop consensus statement on the role of the environment in the development of autoimmune disease. Int. J. Mol. Sci. 2014, 15, 14269–14297. [Google Scholar] [CrossRef] [PubMed]
- Cárdenas-Roldán, J.; Rojas-Villarraga, A.; Anaya, J.M. How do autoimmune diseases cluster in families? A systematic review and meta-analysis. BMC. Med. 2013, 73. [Google Scholar] [CrossRef]
- Lossius, A.; Johansen, J.N.; Torkildsen, O.; Vartdal, F.; Holmøy, T. Epstein-Barr virus in systemic lupus erythematosus, rheumatoid arthritis and multiple sclerosis-association and causation. Viruses 2012, 4, 3701–3730. [Google Scholar] [CrossRef] [PubMed]
- Getts, D.R.; Chastain, E.M.; Terry, R.L.; Miller, S.D. Virus infection, antiviral immunity, and autoimmunity. Immunol. Rev. 2013, 255, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Levin, M.C.; Lee, S.M.; Kalume, F.; Morcos, Y.; Dohan, F.C.; Hasty, K.A.; Callaway, J.C.; Zunt, J.; Desiderio, D.; Stuart, J.M. Autoimmunity due to molecular mimicry as a cause of neurological disease. Nat. Med. 2002, 8, 509–513. [Google Scholar] [CrossRef] [PubMed]
- García-Vallejo, F.; Domínguez, M.C.; Tamayo, O. Autoimmunity and molecular mimicry in tropical spastic paraparesis/human T-lymphotropic virus-associated myelopathy. Braz. J. Med. Biol. Res. 2005, 38, 241–250. [Google Scholar] [PubMed]
- Alamanos, Y.; Voulgari, P.V.; Drosos, A.A. Incidence and prevalence of rheumatoid arthritis, based on the 1987 American College of Rheumatology criteria: A systematic review. Semin. Arthritis Rheum. 2006, 36, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Mota, L.M.H.; Laurindo, I.M.M.; Santos Neto, L.L. Artrite reumatoide inicial–conceitos. Rev. Assoc. Med. Bras. 2010, 56, 227–229. [Google Scholar] [CrossRef] [PubMed]
- Coenen, M.J.H.; Gregersen, P.K. Rheumatoid arthritis: A view of the current genetic landscape. Genes Immun. 2008, 77, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Firestein, G.S. Evolving concepts of rheumatoid arthritis. Nature 2003, 5, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Feldmann, M.; Brennan, F.M. Rheumatoid arthritis. Cell 1996, 85, 307–310. [Google Scholar] [CrossRef]
- Meyer, P.W.; Hodkinson, B.; Ally, M.; Musenge, E.; Wadee, A.A.; Fickl, H.; Tikly, M.; Anderson, R. Circulating cytokine profiles and their relationships with autoantibodies, acute phase reactants, and disease activity in patients with rheumatoid arthritis. Med. Inflamm. 2010, 2010. [Google Scholar] [CrossRef] [PubMed]
- Steiner, G.; Tohidast-Akrad, M.; Witzmann, G.; Vesely, M.; Studnicka-Benke, A.; Gal, A.; Kunaver, M.; Zenz, P.; Smolen, J.S. Cytokine production by synovial T cells in rheumatoid arthritis. Rheumatology 1999, 38, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Nishioka, K.; Nakajima, T.; Hasunuma, T.; Sato, K. Rheumatic manifestation of human leukemia virus infection. Rheum. Dis. Clin. N. Am. 1993, 19, 489–503. [Google Scholar]
- Yakova, M.; Lézin, A.; Dantin, F.; Lagathu, G.; Olindo, S.; Jean-Baptiste, G.; Arfi, S.; Césaire, R. Increased proviral load in HTLV-1-infected patients with rheumatoid arthritis or connective tissue disease. Retrovirology 2005, 1. [Google Scholar] [CrossRef]
- Firestein, G.S.; Zvaifler, N.J. Rheumatoid arthritis: A disease of disordered immunity. In Inflammation: Basic Principles and Clinical Correlates, 2nd ed.; Raven Press Ltd.: New York, NY, USA, 1992; pp. 959–975. [Google Scholar]
- Eguchi, K.; Origuchi, T.; Takashima, H.; Iwata, K.; Katamine, S.; Nagataki, S. High seroprevalence of anti-HTLV-1 antibody in rheumatoid arthritis. Arthritis Rheum. 1996, 39, 463–466. [Google Scholar] [CrossRef] [PubMed]
- Brzustewicz, E.; Bryl, E. The role of cytokines in the pathogenesis of rheumatoid arthritis—Practical and potential application of cytokines as biomarkers and targets of personalized therapy. Cytokine 2015, 76, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Ijichi, S.; Matsuda, T.; Maruyama, I.; Izumihara, T.; Kojima, K.; Niimura, T.; Maruyama, Y.; Sonoda, S.; Yoshida, A.; Osame, M. Arthritis in a human T lymphotropic virus type I (HTLV-I) carrier. Ann. Rheum. Dis. 1990, 49, 718–721. [Google Scholar] [CrossRef] [PubMed]
- Fox, R.I.; Tornwall, J.; Maruyama, T.; Stern, M. Evolving concepts of diagnosis, pathogenesis, and therapy of Sjogren’s syndrome. Curr. Opin. Rheumatol. 1998, 10, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Ambrosi, A.; Wahren-Herlenius, M. Update on the immunobiology of Sjögren′s syndrome. Curr. Opin. Rheumatol. 2015, 27, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, K.; Matsuoka, N.; Ida, H.; Nakashima, M.; Sakai, M.; Sakito, S.; Kawakami, A.; Terada, K.; Shimada, H.; Kawabe, Y. Primary Sjögren’s syndrome with antibodies to HTLV-1: Clinical and laboratory features. Ann. Rheum. Dis. 1992, 51, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Vernant, J.C.; Buisson, G.; Magdeleine, J.; Thore, J.; Jouannelle, A.; Neisson-Vernant, C.; Monplaisir, N. T-lymphocyte alveolitis, tropical spastic paresis, and Sjogren syndrome. Lancet 1988, 1. [Google Scholar] [CrossRef]
- Nakamura, H.; Takahashi, Y.; Yamamoto-Fukuda, T.; Horai, Y.; Nakashima, Y.; Arima, K.; Nakamura, T.; Koji, T.; Kawakami, A. Direct infection of primary salivary gland epithelial cells by human T lymphotropic virus type I in patients with Sjögren′s syndrome. Arthritis Rheumatol. 2015, 67, 1096–1106. [Google Scholar] [CrossRef] [PubMed]
- Scofield, L.; Alarcón, G.S.; Cooper, G.S. Employment and disability issues in systemic lupus erythematosus: A review. Arthritis Care Res. 2008, 59, 1475–1479. [Google Scholar] [CrossRef] [PubMed]
- Cooper, G.S.; Gilbert, K.M.; Greidinger, E.L.; James, J.A.; Pfau, J.C.; Reinlib, L.; Richardson, B.C.; Rose, N.R. Recent advances and opportunities in research on lupus: Environmental influences and mechanisms of disease. Cien. Saude Colet. 2009, 14, 1865–1876. [Google Scholar] [CrossRef] [PubMed]
- Tiffin, N.; Adeyemo, A.; Okpechi, I. A diverse array of genetic factors contribute to the pathogenesis of systemic lupus erythematosus. Orphanet J. Rare Dis. 2013, 7. [Google Scholar] [CrossRef] [PubMed]
- Yap, D.Y.; Lai, K.N. Cytokines and their roles in the pathogenesis of systemic lupus erythematosus: From basics to recent advances. J. Biomed. Biotechnol. 2010, 2010. [Google Scholar] [CrossRef] [PubMed]
- Shirdel, A.; Hashemzadeh, K.; Sahebari, M.; Rafatpanah, H.; Hatef, M.; Rezaieyazdi, Z.; Mirfeizi, Z.; Faridhosseini, R. Is there any association between human lymphotropic virus type I (HTLV-I) infection and systemic lupus erythematosus? Iran. J. Basic Med. Sci. 2013, 16, 252–257. [Google Scholar] [PubMed]
- Sugimoto, T.; Okamoto, M.; Koyama, T.; Takashima, H.; Saeki, M.; Kashiwagi, A.; Horie, M. The occurrence of systemic lupus erythematosus in an asymptomatic carrier of human T-cell lymphotropic virus type I. Clin. Rheumatol. 2007, 26, 1005–1007. [Google Scholar] [CrossRef] [PubMed]
- Akimoto, M.; Matsushita, K.; Suruga, Y.; Aoki, N.; Ozaki, A.; Uozumi, K.; Tei, C.; Arima, N. Clinical manifestations of human T lymphotropic virus type I-infected patients with systemic lupus erythematosus. J. Rheumatol. 2007, 34, 1841–1848. [Google Scholar] [PubMed]
- Magistrelli, C.; Samoilova, E.; Agarwal, R.K.; Banki, K.; Ferrante, P.; Vladutiu, A.; Phillips, P.E.; Perl, A. Polymorphic genotypes of the HRES-1 human endogenous retrovirus locus correlate with systemic lupus erythematosus and autoreactivity. Immunogenetics 1999, 49, 829–834. [Google Scholar] [CrossRef] [PubMed]
- Olsen, R.G.; Tarr, M.J.; Mathes, L.E.; Whisler, R.; du Plessis, D.; Schulz, E.J.; Blakeslee, J.R. Serological and virological evidence of human T-lymphotropic virus in systemic lupus erythematosus. Med. Microbiol. Immunol. 1987, 176, 53–64. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quaresma, J.A.S.; Yoshikawa, G.T.; Koyama, R.V.L.; Dias, G.A.S.; Fujihara, S.; Fuzii, H.T. HTLV-1, Immune Response and Autoimmunity. Viruses 2016, 8, 5. https://doi.org/10.3390/v8010005
Quaresma JAS, Yoshikawa GT, Koyama RVL, Dias GAS, Fujihara S, Fuzii HT. HTLV-1, Immune Response and Autoimmunity. Viruses. 2016; 8(1):5. https://doi.org/10.3390/v8010005
Chicago/Turabian StyleQuaresma, Juarez A S, Gilberto T Yoshikawa, Roberta V L Koyama, George A S Dias, Satomi Fujihara, and Hellen T Fuzii. 2016. "HTLV-1, Immune Response and Autoimmunity" Viruses 8, no. 1: 5. https://doi.org/10.3390/v8010005
APA StyleQuaresma, J. A. S., Yoshikawa, G. T., Koyama, R. V. L., Dias, G. A. S., Fujihara, S., & Fuzii, H. T. (2016). HTLV-1, Immune Response and Autoimmunity. Viruses, 8(1), 5. https://doi.org/10.3390/v8010005