
Myxoma Virus and the Leporipoxviruses: An Evolutionary Paradigm


Abstract
:1. Introduction
2. MYXV and the Leporipoxviruses

| Virus | Natural Host | Disease Caused in Natural Host | Geographic Distribution of Natural Host |
|---|---|---|---|
| Myxoma virus | Sylvilagus brasiliensis (tapeti) | Cutaneous fibroma | South & Central America |
| Californian Myxoma virus | Sylvilagus bachmani (brush rabbit) | Cutaneous fibroma | West coast USA & Mexico |
| Rabbit (Shope) fibroma virus | Sylvilagus floridanus (eastern cottontail) | Cutaneous fibroma | Eastern & central North America to Central and South America |
| Squirrel fibroma virus | Sciurus carolinensis (eastern gray squirrel) 1 | Cutaneous epitheliofibroma; may be generalized and involve internal organs | Eastern North America; introduced in Europe and Britain |
| Hare fibroma virus | Lepus europaeus (European brown hare) 2 | Cutaneous fibroma | Europe but widely introduced to other countries |
| Western squirrel fibroma virus | Sciurus griseus griseus (western gray squirrel) | Cutaneous thickening | West coast of North America |
2.1. The Biology of the Leporipoxviruses in Their Natural Hosts
2.1.1. Myxoma Virus (MYXV)
2.1.2. Californian MYXV (Cal MYXV)
2.1.3. Rabbit Fibroma Virus (RFV)
2.1.4. Hare Fibroma Virus
2.1.5. Squirrel Fibroma Virus
2.2. Species Specificity of Leporipoxviruses
2.3. Genome Sequences of MYXV and Related Leporipoxviruses
| Gene | Protein Function (no. of Amino Acids; Transcription Time: E [Early], L [Late]) | Reference | Effect on Virulence of Gene Disruption |
|---|---|---|---|
| M001L/R | chemokine binding (260; E) | [68] | Generalized myxomatosis; 1/6 survived |
| M002L/R | TNF binding; antiapoptosis (326; E) | [69,70,71,72] | Moderate to severe myxomatosis; 5/8 animals survived |
| M003.1L/R | VACV 1 B15 orthologue; Bcl-2 fold (151; E) | [73] | Not determined |
| M004L/R | RDEL motif; antiapoptosis (237; E) | [74,75] | Small rapidly resolved primary lesions; 1/8 rabbits had a secondary; all animals recovered |
| M005L/R | Antiapoptosis; E3 Ub ligase(483; E) | [76] | Primary lesion only; rapid resolution; no signs of clinical myxomatosis |
| M006L/R | BTB/kelch domains; putative E3 Ub ligase (509; E) | [77] | Not determined |
| M007L/R | Secreted IFN- γ binding protein; chemokine binding (263; E) | [78,79] | 12/13 rabbits mild to moderate disease; lymphocyte infiltration |
| M008L/R | BTB/kelch domains; putative E3 Ub ligase (515; E) | [77] | Not determined |
| M008.1L/R | Serp 1; secreted serine proteinase inhibitor (369; L) | [80] | Moderate to severe generalized myxomatosis; 5/8 rabbits recovered from infection; enhanced inflammatory response |
| M009L | BTB/kelch domains; putative E3 Ub ligase (509; E) | [77] | Not determined |
| M010L | Epidermal growth factor homologue (85; E) | [81] | Generalized myxomatosis; 75% of animals recovered |
| M011L | Antiapoptotic factor (166; E) | [81,82] | All rabbits survived; large protuberant demarcated primary; large secondaries; mild conjunctivitis/rhinitis |
| M013L | Pyrin domain inflammasome (126; E) | [83] | Mild clinical signs rapidly resolved; small secondaries; no mortality; rapid inflammatory response |
| M014L | BTB/kelch domains; putative E3 Ub ligase (517; E) | [77] | Not determined |
| M029L | Type I interferon resistance/PKR inhibition; RNA helicase A binding (115; E) | [84,85] | Abortive infection |
| M036L | VACV O1 orthologue; ERK1/2 signal enhancement (680; E) | [86] | Not determined |
| M062R | Host range (158;E/L ) | [87] | abortive infection in rabbits and rabbit cells |
| M063R | Host range (215; E) | [88] | No virus replication in rabbits and rabbit cells |
| M064R | Virion component (203; E/L) | [89] | Slower progression of disease but lethal in rabbits |
| M104L | potential immunomodulatory (53; L) | [59] | Not determined |
| M121R | NK cell receptor homologue (176; E) | [59] | Not determined |
| M122R | NK cell receptor homologue (172; L) | [59] | Not determined |
| M128L | CD47 homologue; macrophage inhibition (281; L) | [90] | Mild generalized disease; rapid resolution; no deaths |
| M130R | Unknown function; localized to ER/Golgi, glycosylated but not secreted (122; L) | [91] | Generalized myxomatosis but no deaths |
| M131R | Superoxide dismutase inhibition (163; L) | [92,93,94] | All animals euthanized days 10-11; RFV is attenuated |
| M135R | Immunomodulatory (178; E) | [95] | Mild disease with little generalization; all survived |
| M136R | Homology to VACV A52; Bcl-2 fold (179; L?) | [59,73,96] | Not determined |
| M138L | Sialyltransferase (290; E) | [97] | Severe fatal myxomatosis; survival time prolonged |
| M139R | Homology to VACV A52; Bcl-2 fold (188; E) | [73] | Not determined |
| M140R | BTB/kelch domains; putative E3 Ub ligase (553; E?) | [59,77] | Not determined |
| M141R | OX-2 homologue (218; E) | [98] | Mild generalized disease, rapid resolution, all survived; increased macrophage and T cell activation |
| M143R | RING-E3 Ub ligase; possible apoptosis regulator (234; L) | [99,100] | Not determined |
| M146R | VACV N1 orthologue;TLR signal inhibition; Bcl-2 fold antiapoptosis (108; E?) | [59,73,96] | Not determined |
| M148R | Ankyrin repeat; putative E3 Ub ligase (675; L) | [77,101] | Moderate generalized; 2/5 rabbits euthanized at 21 days; mononuclear inflammatory response |
| M149R | Ankyrin repeat; putative E3 Ub ligase (490; E/L?) | [77,101] | Moderate generalized with delayed secondaries; 5/5 rabbits survived |
| M150R | NF-κB inhibition; E3 Ub ligase (494; E) | [77,102,103] | Rapid inflammatory response at primary site; few small secondaries; no respiratory disease; 12/12 recovered by d21 |
| M151R | Serp 2 (333; E) | [104] | Primary lesion but few or no secondary lesions; 7/10 recovered |
| M152R | Serp 3 (266; L) | [105] | 4/10 infected rabbits recovered; 6/10 euthanized because of respiratory disease; no secondary lesions |
| M153R | MHC downreg; E3 Ub ligase (206; E) | [106,107,108] | Generalized myxomatosis; 4/12 rabbits euthanized day 14, the remainder recovered |
| M154L | Downregulation of NFκB? Vac M2 orthologue (214; E) | [59,109] | Not determined |
| M156R | interferon resistance; eIF2α homologue (102; L/E?) | [110] | Not determined |
| Lu Gene | Function | RFV Compared to Lu [59,61] |
|---|---|---|
| M000.5L/R | Unknown | Missing from RFV |
| M008.1L/R | Secreted serine proteinase inhibitor (Serp 1) | Fragmented in RFV |
| M023R | Unknown | 35 aa in RFV; 61 aa in MYXV |
| M119L | unknown | N-terminal truncation |
| M129R | Unknown | 78 aa in RFV; 136 aa in MYXV |
| M135R | Immunomodulatory | Fragmented in RFV |
| M136R | Possible immunomodulatory | Fragmented in RFV |
| M139R | Possible immunomodulatory | Fragmented in RFV |
| M150R | NFκB signal inhibition | Fragmented in RFV |
| M152R | Serp 3 | Fragmented in RFV |
| M156R | eIF2α homologue (IFN resistance) | Truncated N-terminus; duplicated in RFV |
| Lu Gene | Function | MSW Compared to Lu [62] |
|---|---|---|
| M000.5L/R | Unknown | ATG but no ORF in MSW |
| M008.1L/R | Secreted serine proteinase inhibitor (Serp 1) | Multiple stop codons in MSW |
| M009L | BTB/kelch domains; potential E3 Ub ligase | Loss of 845 nts and multiple indels in MSW |
| M023R | Unknown | Mutation of ATG and disruption of ORF in MSW |
| M077L | Assembly complex | 23 aa extra at N-terminus in MSW |
| M119L | Unknown | N-terminal truncation in MSW |
| M131R | Superoxide dismutase homologue | Multiple stop codons in MSW |
| M152R | Serp 3 | Multiple stop codons in MSW |
| M156R | eIF2α homologue (IFN resistance) | Truncated N-terminus; gene duplicated in MSW |
3. Pathogenesis of MYXV in European Rabbits
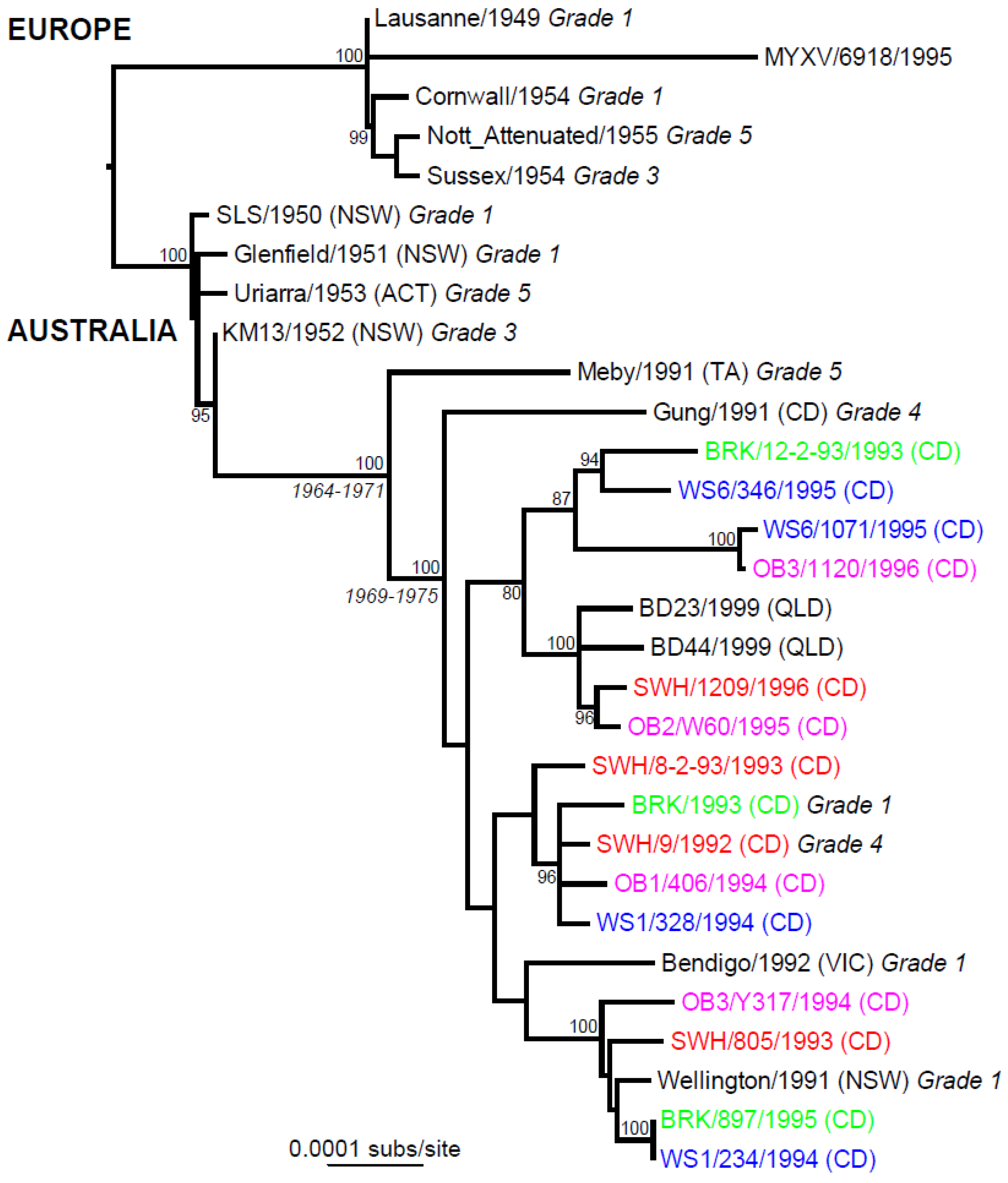
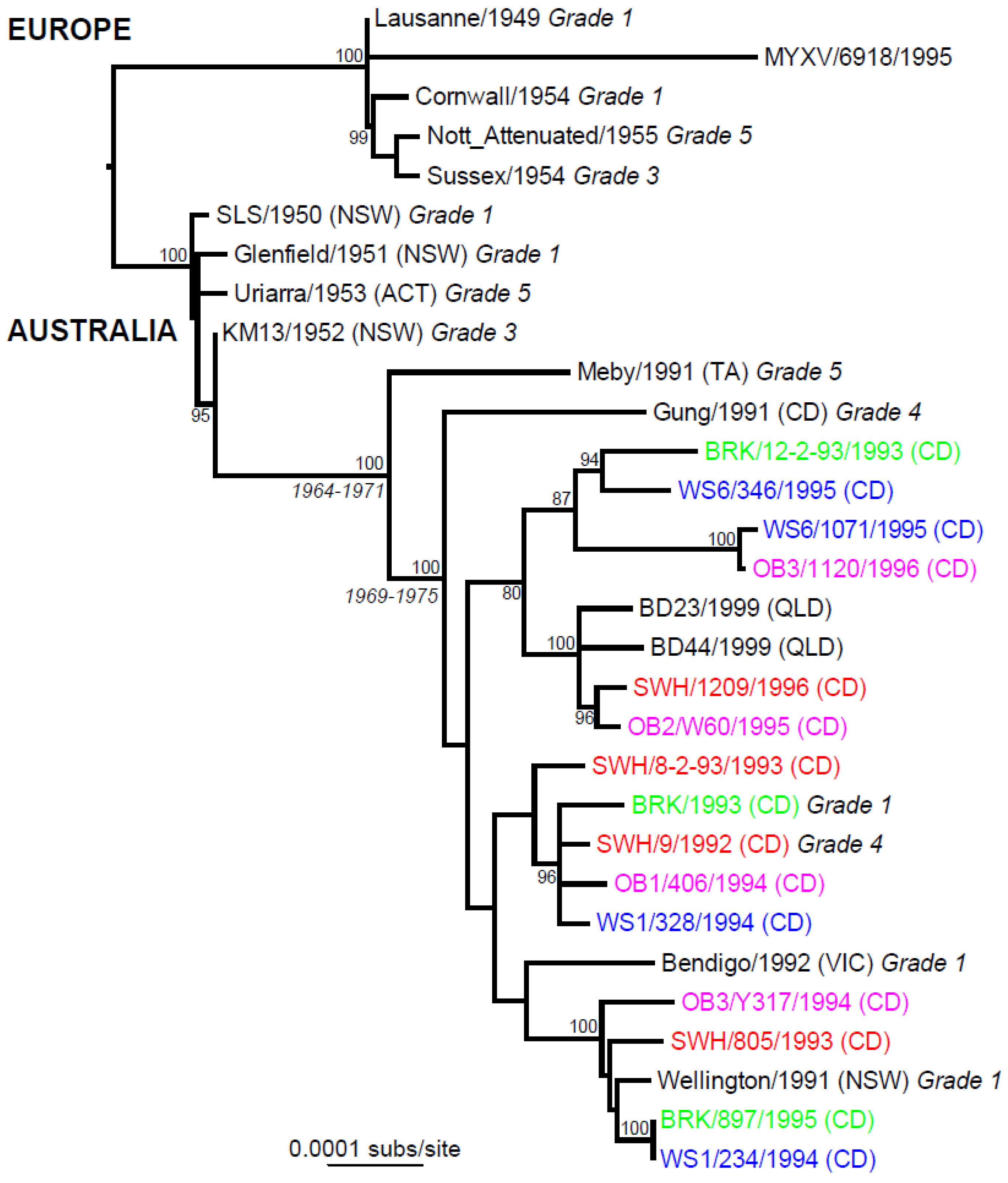
4. Myxoma Virus Evolution in Australia
4.1. The European Rabbit and Initial Trials with MYXV
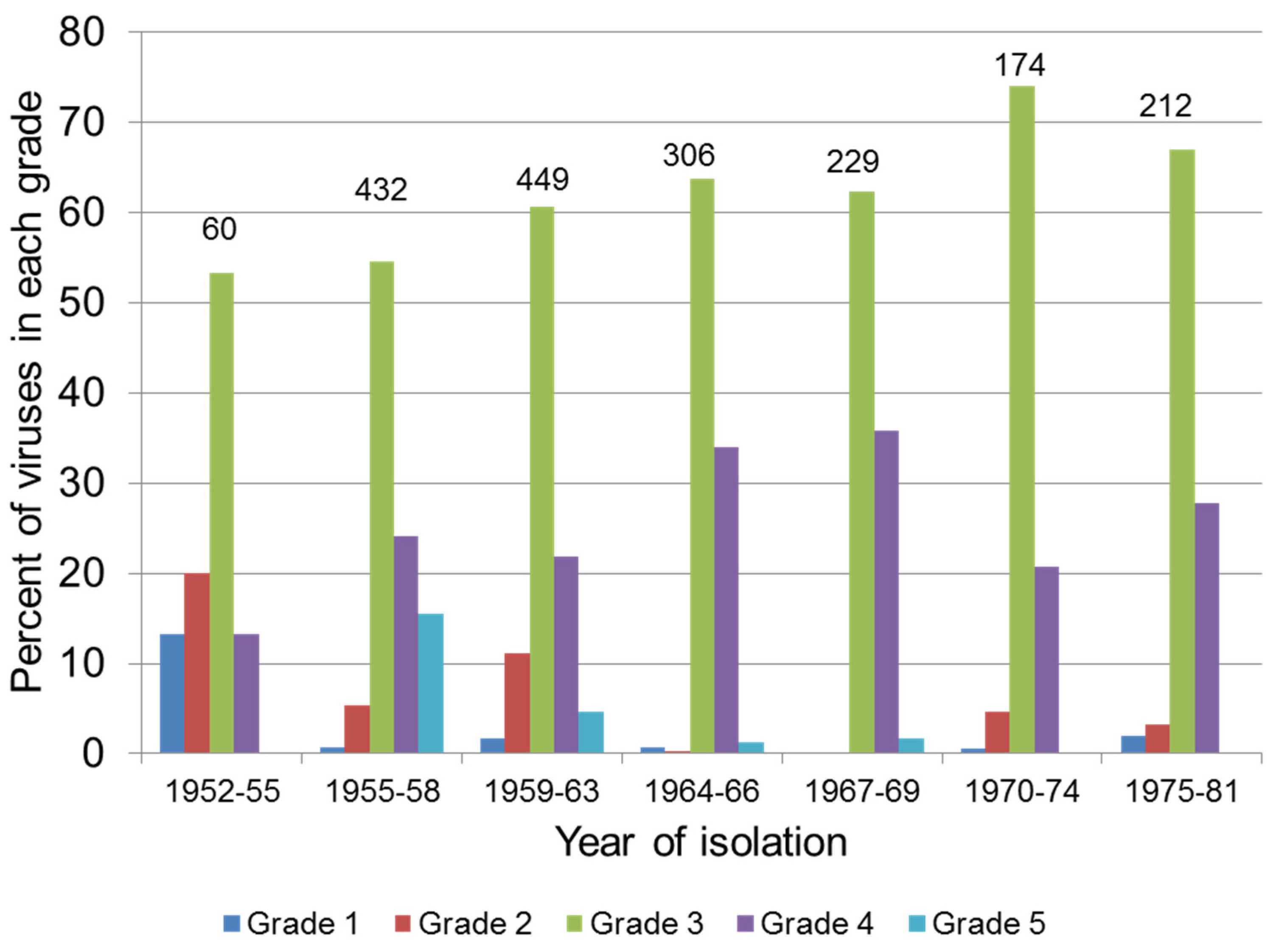
4.2. Attenuation of Field Strains of MYXV
| Virulence Grade | Case Fatality Rate (%) | Average Survival Time (Days) 1 |
|---|---|---|
| 1 | 99.5 | ≤13 |
| 2 | 95–99 | 14–16 |
| 3A | 90–95 | 17–22 |
| 3B | 70–90 | 23–29 |
| 4 | 50–70 | 29–50 |
| 5 | <50 | Not determined |
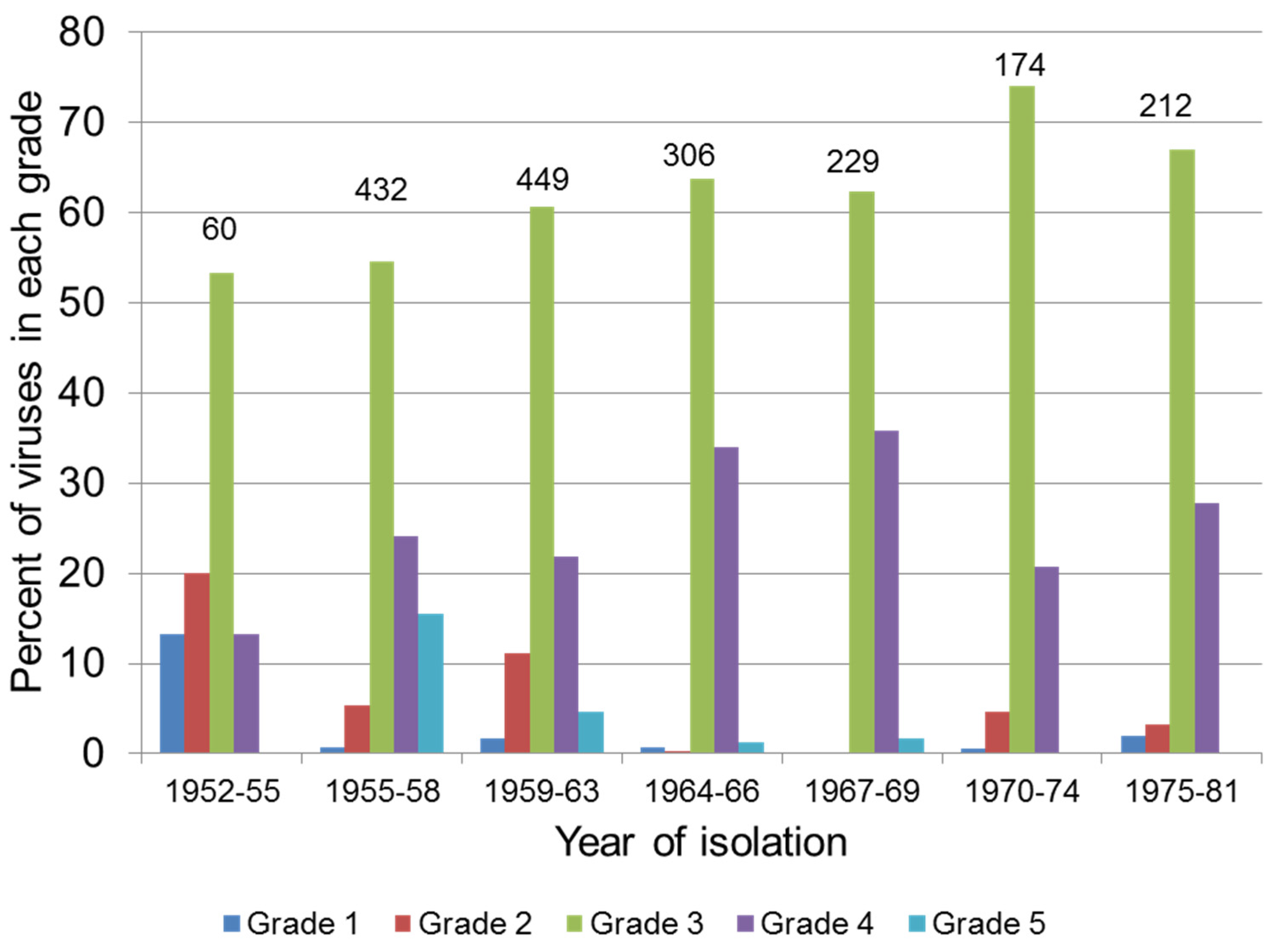
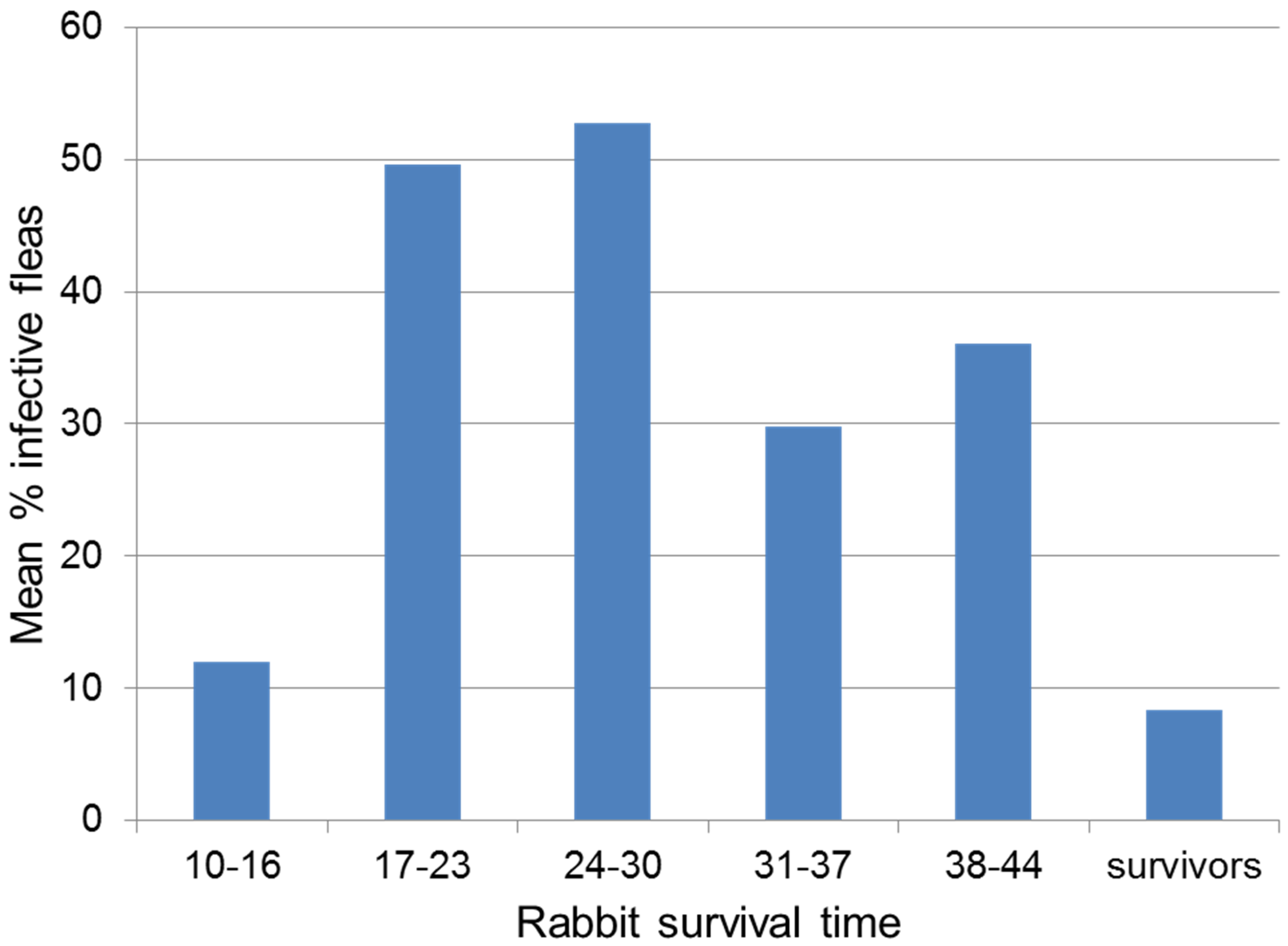
4.3. Selection for Resistance in the Rabbit Population
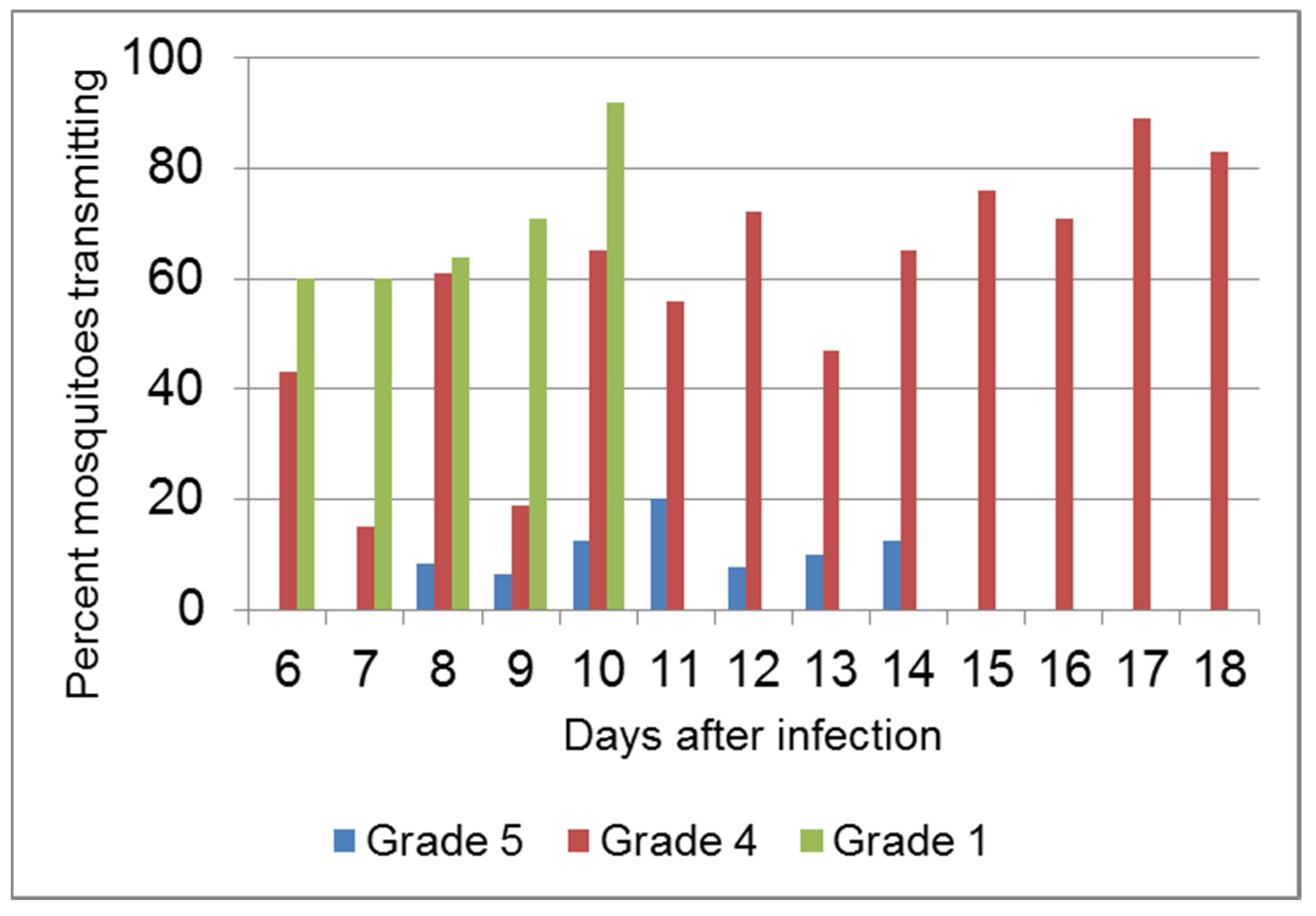
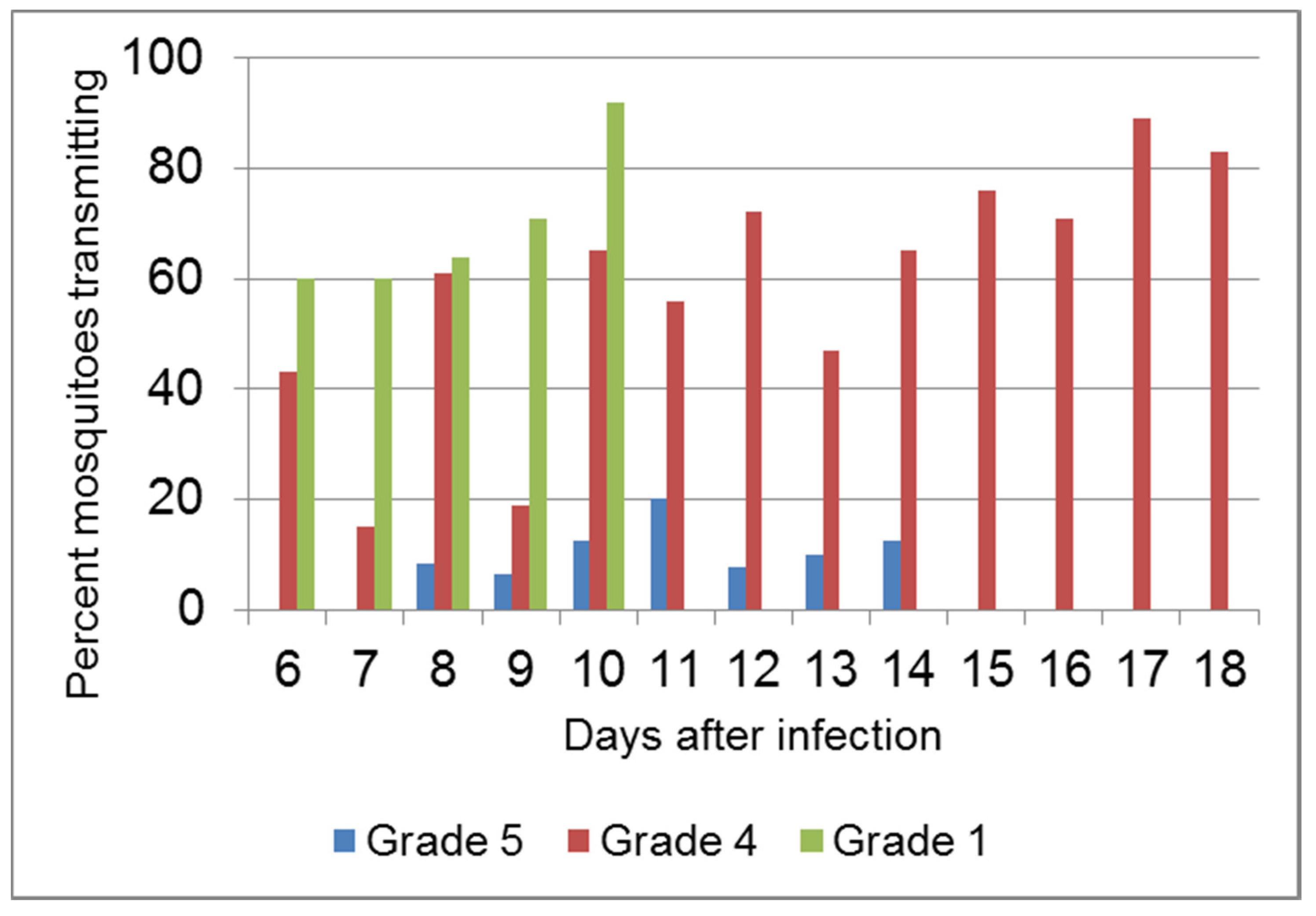
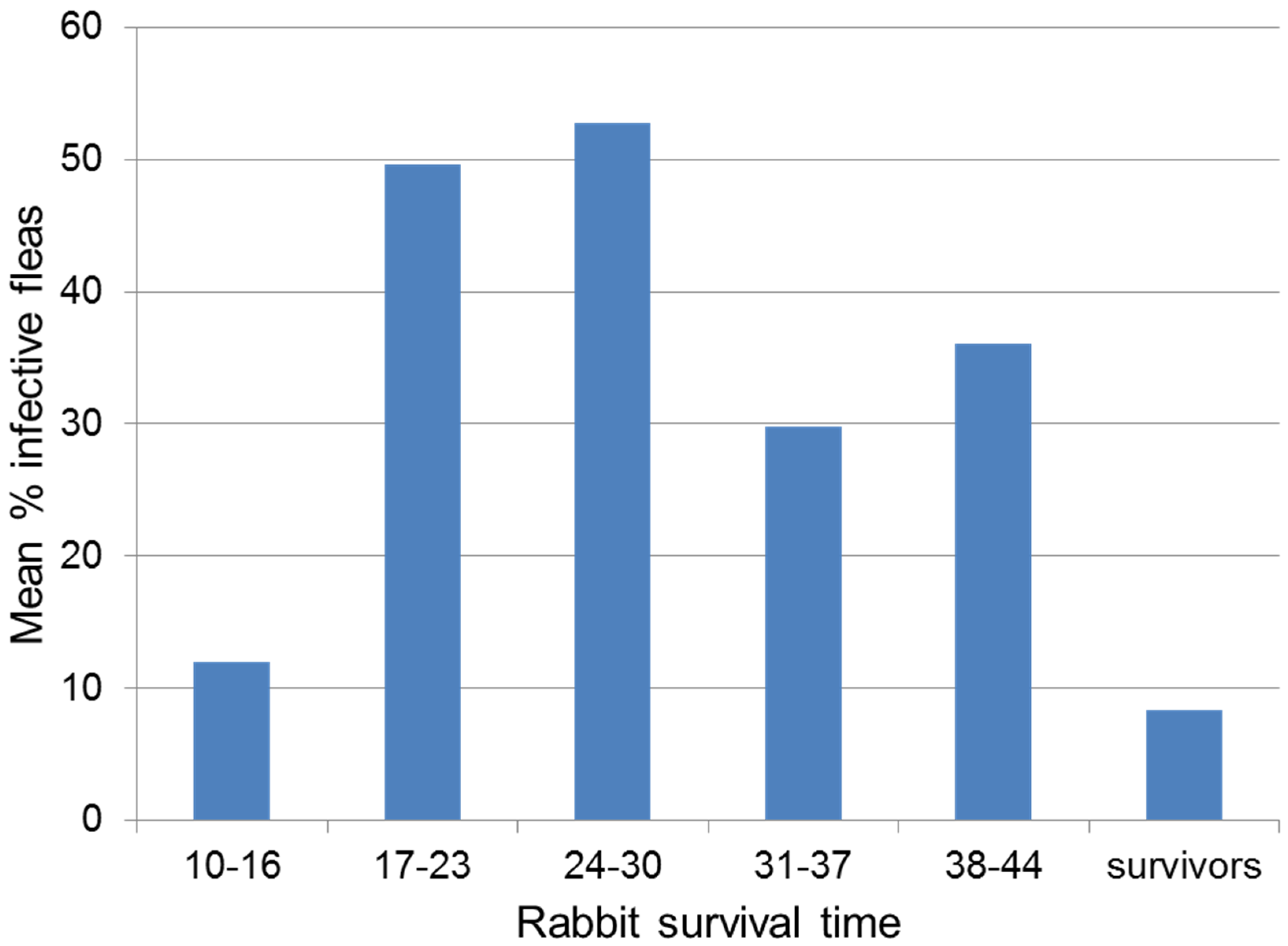
4.4. Virulence and Transmission
5. MYXV Evolution in Europe
5.1. Introduction and Spread
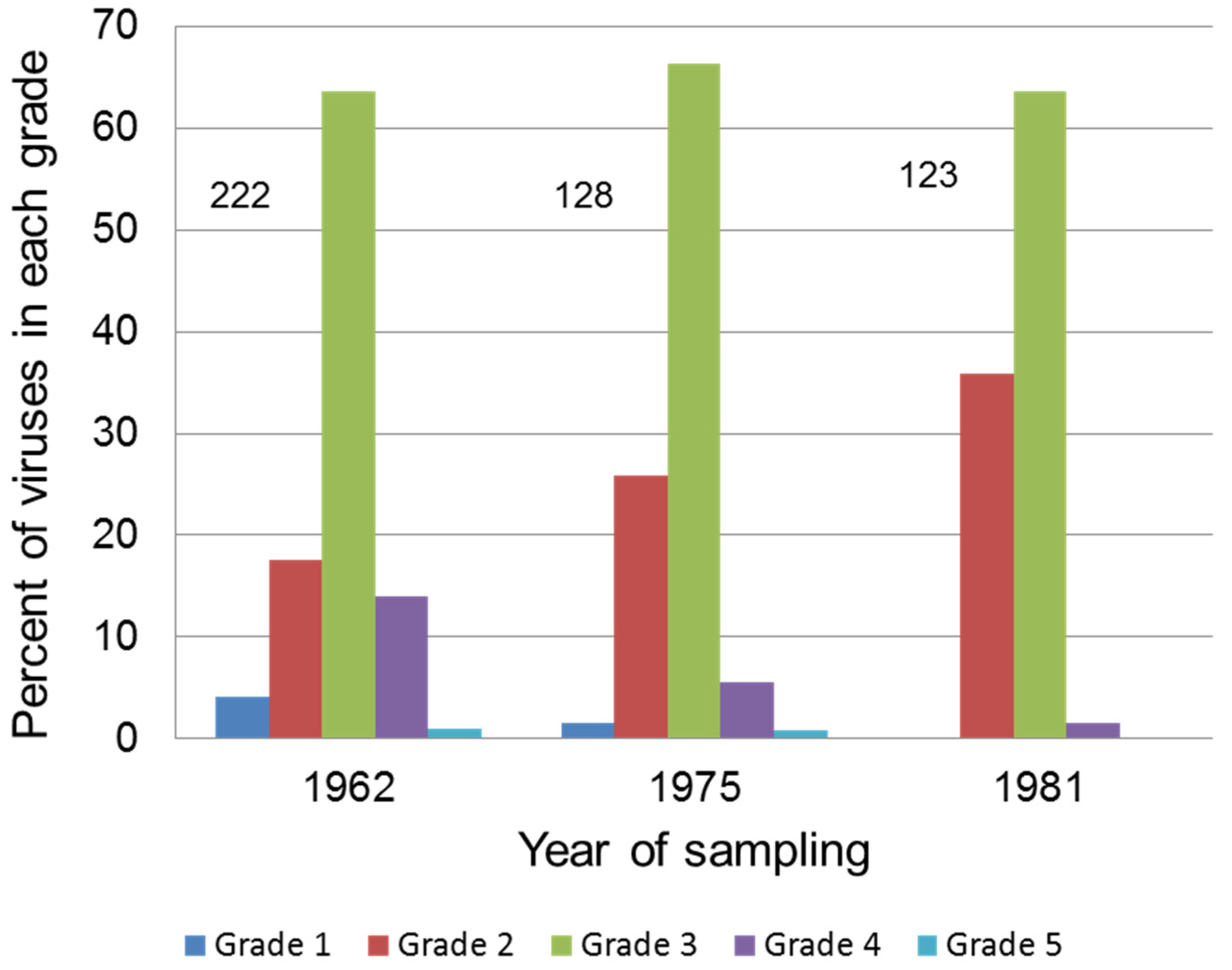
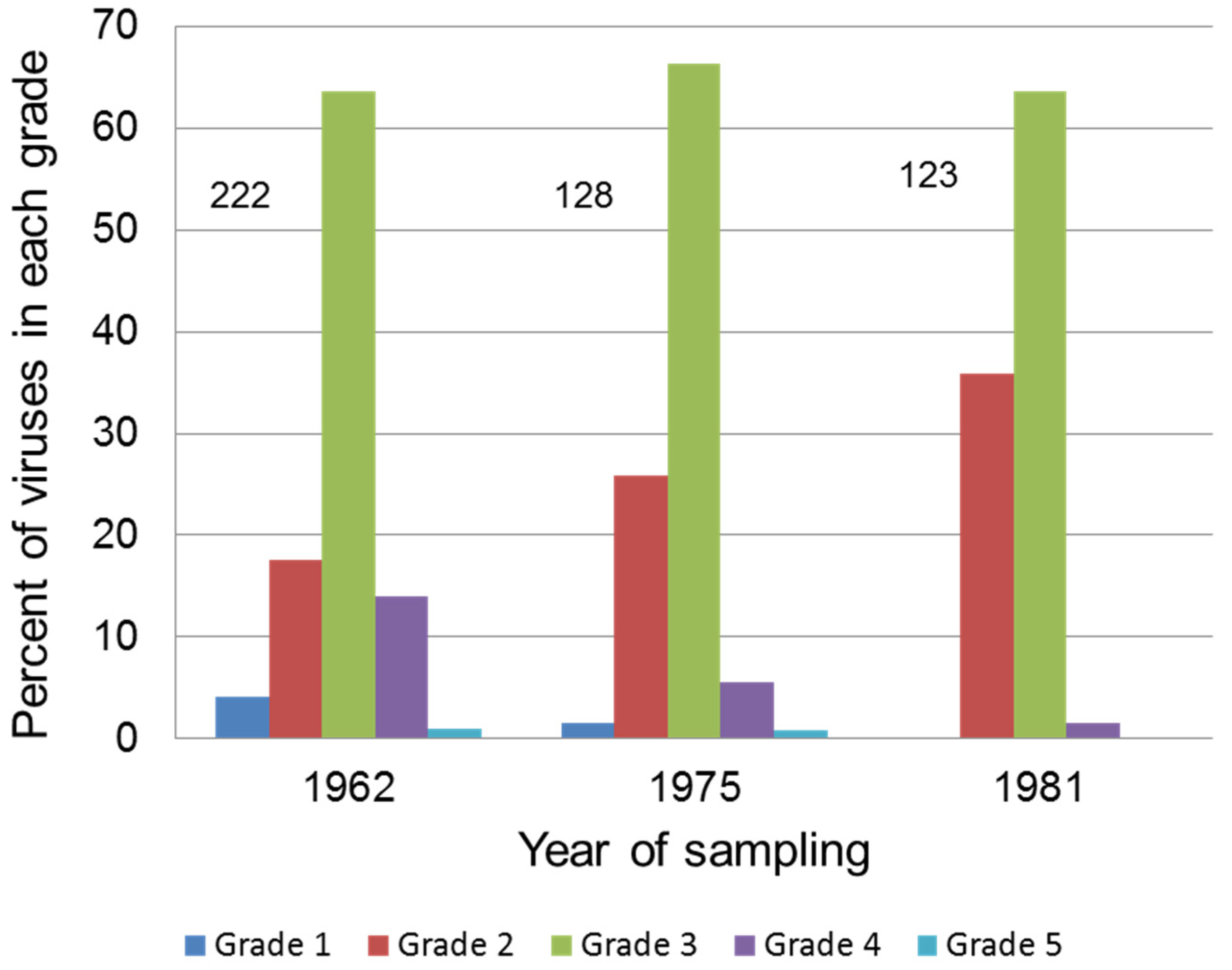
5.2. Attenuation in Europe
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
5.3. Selection for Resistance in Europe
5.4. Evolution of the Phenotype of Myxomatosis in Europe
5.5. Evolution of Vaccine Strains
6. Molecular Evolution of MYXV in Australia and Europe
| Gene | Function (Protein Size aa) | Mutation | Effect | Virus (Virulence Grade if Determined) |
|---|---|---|---|---|
| M000.5L/R | Undetermined (72) | G del | Frameshift from aa 58 and readthrough stop codon adds 5 aa at C terminus | BD44 |
| M005L/R | apoptosis inhibition/host range (483) | C insert (homopol) | Frameshift from aa 284 stop after aa 317; loss of C-terminal F box | WS6 346 |
| M005L/R | apoptosis inhibition/host range (483) | C insert | Frameshift; stop after aa 73 | Ur (5) |
| M008.1L | Secreted Serpin (369) | CC insert (homopol) | Stop after aa 299; loss of active site | BD44 |
| M009L | BTB/kelch domains; putative Ub ligase (509) | A del (homopol) | Additional indel in disrupted ORF | SWH 8/2/93 |
| M009L | BTB/kelch domains; putative Ub ligase (509) | TA insert | Additional indel in disrupted ORF | BRK (1) |
| M009L | BTB/kelch domains; putative Ub ligase (509) | A insert (homopol) | Early stop at aa 146—loss of all kelch domains | All recent Australian except Bendigo (1) |
| M009L | BTB/kelch domains; putative Ub ligase (509) | A del | Early stop at aa 114; additional indel in disrupted ORF | WS6 1071; OB31120 |
| M012L | dUTPpyrophosphatase (148) | 13 nt deletion | Stop after aa 70 | OB3Y317 |
| M014L | BTB/kelch domains; putative Ub ligase (517) | G insert (homopol) | Early stop at aa 477—loss of last kelch domain | Glenfield (1); KM13 (3); Ur (5) |
| M018L | cytoplasmic protein; VACV 1 F8 orthologue (66) | TT insert | Frameshift from aa 60; readthrough adds 20 aa at C-terminus | OB3Y317 |
| M036L | VACV O1 orthologue (680) | 92 nt del | Stop after aa 212 | BRK (1) |
| M036L | VACV O1 orthologue (680) | T insert | Stop after aa 442 | Sussex (3); Nottingham (5) |
| M061R | Thymidine kinase (178) | T insert (homopol) | Readthrough adds LKY to C-terminus | WS1 234 |
| M083L | Carbonic anhydrase homologue/structural (286) | G insert (homopol) | Corrects G deletion in SLS; restores ORF | All recent Australian |
| M130R | Undetermined/virulence (122) | G insert (homopol) | Stop after aa 15 | Glenfield (1) |
| M134R | Transmembrane protein (2000) | A insert (homopol) | Stop at aa 1953; retains predicted C-terminal transmembrane domain | Ur (5); Nottingham (5) |
| M147R | S/T protein kinase (288) | GT del (rpt seq) | Stop after aa 134 | BD23 |
| M150R | NF-κB inhibition (494) | TG insert | Stop after aa 196 | Nottingham (5) |
| M152R | Serp 3 (266/273) | A del (homopol) | Stop after aa 271 | WS6 1071; OB31120 |
| M153R | Ub ligase/MHC-1 downregulation (206) | G del (homopol) | stop after aa 118 | Glenfield (1) |
| M153R | Ub ligase/MHC-1 downregulation (206) | G insert (homopol) | Stop after aa 124 | BD44 |
| M153R | Ub ligase/MHC-1 downregulation (206) | T del | stop after aa 161 | WS6 1071; OB31120 |
| M153R | Ub ligase/MHC-1 downregulation (206) | 73 nt del | Sequence read through replaces C-terminal CR domain of M153 | Meby (5) |
| M156R | eIF2α homologue; IFN resistance (102) | T del | Read through stop—extra EG at C-terminus | WS6 346; OB3Y317 |
7. Phylogeography
8. Future Evolution of Host and Pathogen
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Burnet, M. Changing Patterns; Heinemann: Melbourne, Australia, 1968. [Google Scholar]
- May, R.M.; Anderson, R.M. Epidemiology and genetics in the coevolution of parasites and hosts. Proc. R. Soc. Lond. B 1983, 219, 281–313. [Google Scholar] [CrossRef] [PubMed]
- Ratcliffe, F.N.; Myers, K.; Fennessy, B.V.; Calaby, J.H. Myxomatosis in Australia. A step towards the biological control of the rabbit. Nature 1952, 170, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Myers, K. Studies in the epidemiology of infectious myxomatosis of rabbits. II. Field experiments, August-November 1950, and the first epizootic of myxomatosis in the Riverine plain of south-eastern Australia. J. Hyg. 1954, 52, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Bull, L.B.; Mules, M.W. An investigation of myxomatosis cuniculi with special reference to the possible use of the disease to control rabbit populations in Australia. J. CSIR 1944, 17, 79–93. [Google Scholar]
- Lockley, R.M. Some experiments in rabbit control. Nature 1940, 145, 767–769. [Google Scholar] [CrossRef]
- Lockley, R.M. Failure of myxomatosis on Skokholm Island. Nature 1955, 175, 906–907. [Google Scholar] [CrossRef] [PubMed]
- Fenner, F.; Ratcliffe, F.N. Myxomatosis; Cambridge University Press: Cambridge, UK, 1965. [Google Scholar]
- Sanarelli, G. Das myxomatogene Virus. Beitrag zum Stadium der Krankheitserreger ausserhalb des Sichtbaren. Zbl. Bakt. 1898, 23, 865–873. (in German). [Google Scholar]
- Kilham, L. Metastasizing viral fibromas of gray squirrels: Pathogenesis and mosquito transmission. Am. J. Hyg. 1955, 61, 55–63. [Google Scholar] [PubMed]
- Kilham, L.; Herman, C.M.; Fisher, E.R. Naturally occurring fibromas of grey squirrels related to Shope’s rabbit fibroma. Proc. Soc. Exper. Biol. Med. 1953, 82, 298–301. [Google Scholar] [CrossRef]
- Regnery, D.C.; Marshall, I.D. Studies in the epidemiology of myxomatosis in California. IV. The susceptibility of six leporid species to Californian myxoma virus and the relative infectivity of their tumours for mosquitoes. Am. J. Epidemiol. 1971, 94, 508–513. [Google Scholar] [PubMed]
- Karstad, L.; Thorsen, J.; Davies, G.; Kaminjolo, J.S. Poxvirus fibromas on African hares. J. Wildl. Dis. 1977, 13, 245–247. [Google Scholar] [CrossRef] [PubMed]
- Fenner, F.; Woodroofe, G.M. Protection of laboratory rabbits against myxomatosis by vaccination with fibroma virus. Aust. J. Exptl. Biol. Med. Sci. 1954, 32, 653–668. [Google Scholar] [CrossRef]
- Marlier, D. Vaccination against myxoma virus infections: Are we really doing the best? Tijdschr. Diergeneeskd. 2010, 135, 194–198. [Google Scholar] [PubMed]
- Fenner, F.; Marshall, I.D. A comparison of the virulence for European rabbits (Oryctolagus cuniculus) of strains of myxoma virus recovered in the field in Australia, Europe and America. J. Hyg. 1957, 55, 149–191. [Google Scholar] [CrossRef] [PubMed]
- Silvers, L.; Inglis, B.; Labudovic, A.; Janssens, P.A.; van Leeuwen, B.H.; Kerr, P.J. Virulence and pathogenesis of the MSW and MSD strains of Californian myxoma virus in European rabbits with genetic resistance to myxoma virus compared to rabbits with no genetic resistance. Virology 2006, 348, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Woodroofe, G.M.; Fenner, F. Viruses of the myxoma-fibroma subgroup of the poxviruses. I. Plaque production in cultured cells, plaque-reduction tests and cross-protection tests in rabbits. Aust. J. Exptl. Biol. Med. Sci. 1965, 43, 123–142. [Google Scholar] [CrossRef]
- Fenner, F. Viruses of the myxoma-fibroma subgroup of the poxviruses. II. Comparison of soluble antigens by gel diffusion tests, and a general discussion of the subgroup. Aust. J. Exp. Biol. Med. Sci. 1965, 43, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Aragão, H.D.B. O virus do mixoma no coelho do mato (Sylvilagus. minensus) sua transmissao pelos Aedes. scapularis e aegypti. Mem. Inst. Oswaldo Cruz 1943, 38, 93–99. (in Portuguese). [Google Scholar]
- Howard, W.E.; Amaya, J.N. European rabbit invades western Argentina. J. Wildl. Man. 1975, 39, 757–761. [Google Scholar] [CrossRef]
- Jaksic, F.M.; Yanez, J.L. Rabbit and fox introductions in Tierra del Fuego: History and assessment of the attempts at biological control of the rabbit infestation. Biol. Conserv. 1983, 26, 367–374. [Google Scholar] [CrossRef]
- Grodhaus, G.; Regnery, D.C.; Marshall, I.D. Studies in the epidemioloy of myxomatosis in California. II. The experimental transmission of myxomatosis between brush rabbits (Sylvilagus bachmani) by several species of mosquitoes. Am. J. Hyg. 1963, 77, 205–212. [Google Scholar] [PubMed]
- Marshall, I.D.; Regnery, D.C.; Grodhaus, G. Studies in the epidemiology of myxomatosis in California. I. Observations on two outbreaks of myxomatosis in coastal California and the recovery of myxoma virus from a brush rabbit (Sylvilagus bachmani). Am. J. Hyg. 1963, 77, 195–204. [Google Scholar]
- Kessel, J.F.; Prouty, C.C.; Meyer, J.W. Occurence of infectious myxomatosis in southern California. Proc. Soc. Exp. Biol. Med. 1931, 28, 413–414. [Google Scholar] [CrossRef]
- Kessel, J.F.; Fiskm, R.T.; Prouty, C.C. Studies with the Californian Strain of the Virus of Infectious Myxomatosis. In Proceedings of the Fifth Pacific Science Congress; University of Toronto Press: Toronto, ON, Canada, 1934; pp. 2927–2939. [Google Scholar]
- Marshall, I.D.; Regnery, D.C. Myxomatosis in a Californian brush rabbit. Nature 1960, 188, 73–74. [Google Scholar] [CrossRef] [PubMed]
- Regnery, D.C.; Miller, J.H. A myxoma virus epizootic in a brush rabbit population. J. Wildl. Dis. 1972, 8, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Patton, N.M.; Holmes, H.T. Myxomatosis in domestic rabbits in Oregon. J. Am. Vet. Assoc. 1977, 171, 560–562. [Google Scholar]
- Luna, R.M.L. First report of myxomatosis in Mexico. J. Wildl. Dis. 2000, 36, 580–583. [Google Scholar] [CrossRef] [PubMed]
- Shope, R.E. A transmissible tumour-like condition in rabbits. J. Exp. Med. 1932, 56, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Shope, R.E. A filtrable virus causing a tumour-like condition in rabbits and its relationship to virus myxomatosum. J. Exp. Med. 1932, 56, 803–822. [Google Scholar] [CrossRef] [PubMed]
- Kilham, L.; Woke, P.A. Laboratory transmission of fibromas (Shope) in cottontail rabbits by means of fleas and mosquitoes. Proc. Soc. Exp. Biol. Med. 1953, 83, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Dalmat, H.T. Arthropod transmission of rabbit fibromatosis (Shope). J. Hyg. 1959, 57, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Kilham, L.; Dalmat, H.T. Host-virus-mosquito relations of Shope fibromas in cottontail rabbits. Am. J. Hyg. 1955, 61, 45–54. [Google Scholar] [PubMed]
- Kilham, L.; Fisher, E.R. Pathogenesis of fibromas in cottontail rabbits. Am. J. Hyg. 1954, 61, 45–54. [Google Scholar]
- Yuill, T.M.; Hanson, R.P. Infection of suckling cottontail rabbits with Shope’s fibroma virus. Proc. Soc. Exp. Biol. Med. 1964, 117, 376–380. [Google Scholar] [CrossRef] [PubMed]
- Joiner, G.N.; Jardine, J.H.; Gleiser, C.A. An epizootic of Shope fibromatosis in a commercial rabbitry. J. Am. Vet. Med. Assoc. 1971, 159, 1583–1587. [Google Scholar] [PubMed]
- Raflo, C.P.; Olsen, R.G.; Pakes, S.P.; Webster, W.S. Characterization of a fibroma virus isolated from naturally-occurring skin tumors in domestic rabbits. Lab. Anim. Sci. 1973, 23, 525–532. [Google Scholar] [PubMed]
- Arthur, C.P.; Louzis, C. La myxomatose du lapin en France: Une revue. Rev. Sci. Tech. Off. Int. Epiz. 1988, 7, 937–957. (in French). [Google Scholar]
- Rogers, P.M.; Arthur, C.P.; Soriguer, R.C. The rabbit in continental Europe. In The European Rabbit The History and Biology of a Successful Colonizer; Thompson, H.V., King, C.M., Eds.; Oxford University Press: Oxford, UK, 1994; pp. 22–63. [Google Scholar]
- Fenner, F. Hare fibroma virus. In Virus Infections of Rodents and Lagomorphs; Osterhaus, A.D.M.E., Ed.; Elsevier Science B.V.: Amsterdam, The Netherlands, 1994; Volume 5, pp. 77–79. [Google Scholar]
- Micozzi, G.; Palarchi, M. Spontaneous myxomatosis in hares. Vet. Ital. 1965, 26, 361–362. [Google Scholar]
- Barlow, A.; Lawrence, K.; Everest, D.; Dastjerdi, A.; Finnegan, C.; Steinbach, F. Confirmation of myxomatosis in a European brown hare in Great Britain. Vet. Rec. 2014, 175, 75–76. [Google Scholar] [CrossRef] [PubMed]
- Kirschstein, R.L.; Rabson, A.S.; Kilham, L. Pulmonary lesions produced by fibroma viruses in squirrels and rabbits. Cancer Res. 1958, 18, 1340–1344. [Google Scholar] [PubMed]
- King, J.M.; Woolf, A.; Shively, J.N. Naturally occurring squirrel fibroma virus with involvement of internal organs. J. Wildl. Dis. 1972, 8, 321–324. [Google Scholar] [CrossRef] [PubMed]
- Hirth, R.S.; Wyand, D.S.; Osborne, A.D.; Burke, C.N. Epidermal changes caused by squirrel poxvirus. J. Am. Vet. Med. Assoc. 1969, 155, 1120–1125. [Google Scholar] [PubMed]
- Novilla, M.N.; Flyger, V.; Jacobson, E.R.; Dutta, S.K.; Sacchi, E.M. Systemic phycomycosis and multiple fibromas in a gray squirrel (Sciurus carolinensis). J. Wildl. Dis. 1981, 17, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Herman, C.M.; Reilly, J.R. Skin tumors on squirrels. J. Wildl. Man. 1955, 19, 402–403. [Google Scholar] [CrossRef]
- Regnery, R.L. Preliminary studies on an unusual poxvirus of the western grey squirrel (Sciurus griseus griseus) of North America. Intervirology 1975, 5, 364–366. [Google Scholar] [PubMed]
- Martin, C.J. Observations on Myxomatosis cuniculi (Sanarelli) Made with a View to the Use of the Virus in the Control of Rabbit Plagues; C.S.I.R: Melbourne, Australia, 1936. [Google Scholar]
- Bull, L.B.; Dickinson, C.G. The specificity of the virus of rabbit myxomatosis. J. CSIR 1937, 10, 291–294. [Google Scholar]
- Marshall, I.D.; Regnery, D.C. Studies in the epidemiology of myxomatosis in California. III. The response of brush rabbits (Sylvilagus bachmani) to infection with exotic and enzootic strains of myxoma virus and the relative infectivity of the tumours for mosquitoes. Am. J. Hyg. 1963, 77, 213–219. [Google Scholar]
- Regnery, D.C. The epidemic potential of Brazilian myxoma virus (Lausanne strain) for three species of North American cottontails. Am. J. Epidemiol. 1971, 94, 514–519. [Google Scholar] [PubMed]
- Day, M.F.; Fenner, F.; Woodroofe, G.M.; McIntyre, G.A. Further studies on the mechanism of mosquito transmission of myxomatosis in the European rabbit. J. Hyg. 1956, 54, 258–283. [Google Scholar] [CrossRef] [PubMed]
- Dalmat, H.T.; Stanton, M.F. A comparative study of the Shope Fibroma in rabbits in relation to transmissibility by mosquitoes. J. Natl. Cancer Inst. 1959, 22, 593–615. [Google Scholar] [PubMed]
- Fenner, F.; Ross, J. Myxomatosis. In The European Rabbit. The History and Biology of a Successful Colonizer; Thompson, H.V., King, C.M., Eds.; Oxford University Press: Oxford, UK, 1994; pp. 205–240. [Google Scholar]
- Silvers, L.; Barnard, D.; Knowlton, F.; Inglis, B.; Labudovic, A.; Holland, M.K.; Janssens, P.A.; van Leeuwen, B.A.; Kerr, P.J. Host-specificity of myxoma virus: Pathogenesis of South American and North American strains of myxoma virus in two North American lagomorph species. Vet. Microbiol. 2010, 141, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Cameron, C.; Hota-Mitchell, S.; Chen, L.; Barrett, J.; Cao, J.-X.; Macaulay, C.; Willer, D.; Evans, D.; McFadden, G. The complete DNA sequence of myxoma virus. Virology 1999, 264, 298–318. [Google Scholar] [CrossRef] [PubMed]
- Morales, M.; Ramirez, M.A.; Cano, M.J.; Parraga, M.; Castilla, J.; Perez-Ordoyo, L.I.; Torres, J.M.; Barcena, J. Genome comparison of a nonpathogenic myxoma virus field strain with its ancestor, the virulent Lausanne strain. J. Virol. 2009, 83, 2397–2403. [Google Scholar] [CrossRef] [PubMed]
- Willer, D.O.; McFadden, G.; Evans, D.H. The complete genome sequence of Shope (rabbit) fibroma virus. Virology 1999, 264, 319–343. [Google Scholar] [CrossRef] [PubMed]
- Kerr, P.J.; Rogers, M.B.; Fitch, A.; Depasse, J.V.; Cattadori, I.M.; Hudson, P.J.; Tscharke, D.C.; Holmes, E.C.; Ghedin, E. Comparative analysis of the complete genome sequence of the California MSW strain of myxoma virus reveals potential host adaptations. J. Virol. 2013, 87, 12080–12089. [Google Scholar] [CrossRef] [PubMed]
- Upton, C.; Slack, S.; Hunter, A.L.; Ehlers, A.; Roper, R.L. Poxvirus orthologous clusters: Towards defining the minimal essential poxvirus genome. J. Virol. 2003, 77, 7590–7600. [Google Scholar] [CrossRef] [PubMed]
- Kerr, P.J. Myxomatosis in Australia and Europe: A model for emerging infectious diseases. Antiviral Res. 2012, 93, 387–415. [Google Scholar] [CrossRef] [PubMed]
- Allison, A.C. Immune responses to Shope fibroma virus in adult and newborn rabbits. J. Natl. Cancer Inst. 1966, 36, 869–875. [Google Scholar]
- Allison, A.C.; Friedman, R.M. Effects of immunosuppressants on Shope rabbit fibroma. J. Natl. Cancer Inst. 1966, 36, 859–868. [Google Scholar]
- Sell, S.; Scott, C.B. An immunohistologic study of Shope Fibroma Virus in rabbits: Tumor rejection by cellular reaction in adults and progressive systemic reticuloendothelial infection in neonates. J. Natl. Cancer Inst. 1981, 66, 363–373. [Google Scholar] [PubMed]
- Lalani, A.S.; Masters, J.; Graham, K.; Liu, L.; Lucas, A.; McFadden, G. Role of the myxoma virus soluble CC-chemokine inhibitor glycoprotein, M-T1, during myxoma virus pathogenesis. Virology 1999, 256, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Upton, C.; Macen, J.L.; Schreiber, M.; McFadden, G. Myxoma virus expresses a secreted protein with homology to the tumour necrosis factor receptor gene family that contributes to viral virulence. Virology 1991, 184, 370–382. [Google Scholar] [CrossRef] [PubMed]
- Macen, J.L.; Graham, K.A.; Fong Lee, S.; Schreiber, M.; Boshkov, L.K.; McFadden, G. Expression of the myxoma virus tumour necrosis factor receptor homologue and M11L genes is required to prevent virus-induced apoptosis in infected rabbit T lymphocytes. Virology 1996, 218, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, M.; Sedger, L.; McFadden, G. Distinct domains of M-T2, the myxoma virus tumor necrosis factor (TNF) receptor homolog, mediate extracellular TNF binding and intracellular apoptosis inhibition. J. Virol. 1997, 71, 2171–2191. [Google Scholar] [PubMed]
- Sedger, L.M.; Osvath, S.R.; Xu, X.-M.; Li, G.; Chan, F.K.-M.; Barrett, J.W.; McFadden, G. Poxvirus tumor necrosis factor receptor (TNFR)-like T2 proteins contain a conserved preligand assembly domain that inhibits cellular TNFR1-induced cell death. J. Virol. 2006, 80, 9300–9309. [Google Scholar] [CrossRef] [PubMed]
- González, J.M.; Esteban, M. A poxvirus Bcl-2-like gene family involved in regulation of host immune response: Sequence similarity and evolutionary history. Virol. J. 2010, 7, e59. [Google Scholar] [CrossRef]
- Hnatiuk, S.; Barry, M.; Zeng, W.; Liu, L.; Lucas, A.; Percy, D.; McFadden, G. Role of the C-terminal RDEL motif of the myxoma virus M-T4 protein in terms of apoptosis regulation and viral pathogenesis. Virology 1999, 263, 290–306. [Google Scholar] [CrossRef] [PubMed]
- Barry, M.; Hnatiuk, S.; Mossman, K.; Lee, S.F.; Boshkov, L.; McFadden, G. The myxoma virus M-T4 gene encodes a novel RDEL-containing protein that is retained within the endoplasmic reticulum and is important for the productive infection of lymphocytes. Virology 1997, 239, 360–377. [Google Scholar] [CrossRef] [PubMed]
- Mossman, K.; Fong Lee, S.; Barry, M.; Boshkov, L.; McFadden, G. Disruption of M-T5, a novel myxoma virus gene member of the poxvirus host range superfamily, results in dramatic attenuation of myxomatosis in infected European rabbits. J. Virol. 1996, 70, 4394–4410. [Google Scholar] [PubMed]
- Zhang, L.; Villa, N.Y.; McFadden, G. Interplay between poxviruses and the cellular ubiquitin/ ubiquitin-like pathways. FEBS Lett. 2009, 583, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Mossman, K.; Nation, P.; Macen, J.; Garbutt, M.; Lucas, A.; McFadden, G. Myxoma virus M-T7, a secreted homolog of the interferon-γ receptor, is a critical virulence factor for the development of myxomatosis in European rabbits. Virology 1996, 215, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Lalani, A.S.; Graham, K.; Mossman, K.; Rajarathnam, K.; Clark-Lewis, I.; Kelvin, D.; McFadden, G. The purified myxoma virus gamma interferon receptor homolog M-T7 interacts with the heparin-binding domains of chemokines. J. Virol. 1997, 71, 4356–4363. [Google Scholar] [PubMed]
- Macen, J.L.; Upton, C.; Nation, N.; McFadden, G. Serp 1, a secreted proteinase inhibitor encoded by myxoma virus is a secreted glycoprotein that interferes with inflammation. Virology 1993, 195, 348–363. [Google Scholar] [CrossRef] [PubMed]
- Opgenorth, A.; Graham, K.; Nation, N.; Strayer, D.; McFadden, G. Deletion analysis of two tandemly arranged virulence genes in myxoma virus, M11L and myxoma growth factor. J. Virol. 1992, 66, 4720–4731. [Google Scholar] [PubMed]
- Graham, K.A.; Opgenorth, A.; Upton, C.; McFadden, G. Myxoma virus M11L ORF encodes a protein for which cell surface localization is critical in manifestation of viral virulence. Virology 1992, 191, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Johnston, J.B.; Barrett, J.W.; Nazarian, S.H.; Goodwin, M.; Ricuttio, D.; Wang, G.; McFadden, G. A poxvirus-encoded pyrin domain protein interacts with ASC-1 to inhibit host inflammatory and apoptotic responses to infection. Immunity 2005, 23, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Myskiw, C.; Arsenio, J.; Hammett, C.; van Bruggen, R.; Deschambault, Y.; Beausoleil, N.; Babiuk, S.; Cao, J. Comparative analysis of poxvirus orthologues of the Vaccinia Virus E3 protein: Modulation of PKR activity, cytokine responses and virus pathogenicity. J. Virol. 2011, 85, 12280–12291. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Liu, J.; Chan, W.M.; Rothenburg, S.; McFadden, G. Myxoma virus protein M029 is a dual function immunomodulator that inhibits PKR and also conscripts RHA/DHX9 to promote expanded host-tropism and viral replication. PLOS Pathog. 2013, 9, e1003465. [Google Scholar] [CrossRef] [PubMed]
- Schweneker, M.; Lukassen, S.; Spath, M.; Wolferstatter, M.; Babel, E.; Brinkmann, K.; Wielert, U.; Chaplin, P.; Suter, M.; Hausmann, J. The vaccinia virus O1 protein is required for sustained activation of extracellular signal-regulated kinase 1/2 and promotes viral virulence. J. Virol. 2012, 86, 2323–2336. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wennier, S.; Zhang, L.; McFadden, G. M062 is a host range factor essential for myxoma virus pathogenesis and functions as an antagonist of host SAMD9 in human cells. J. Virol. 2011, 85, 3270–3282. [Google Scholar] [CrossRef] [PubMed]
- Barrett, J.W.; Shun Chang, C.; Wang, G.; Werden, S.J.; Shao, Z.; Barrett, C.; Gao, X.; Belsito, T.A.; Villenevue, D.; McFadden, G. Myxoma virus M063R is a host range gene essential for virus replication in rabbit cells. Virology 2007, 361, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wennier, S.; Moussatche, N.; Reinhard, M.; Condit, R.C.; McFadden, G. Myxoma virus M064 is a novel member of the poxvirus C7L superfamily of host-range factors that controls the kinetics of myxomatosis in European rabbits. J. Virol. 2012, 86, 5371–5375. [Google Scholar] [CrossRef] [PubMed]
- Cameron, C.M.; Barrett, J.W.; Mann, M.; Lucas, A.; McFadden, G. Myxoma virus M128L is expressed as a cell surface CD47-like virulence factor that contributes to the downregulation of macrophage activation in vivo. Virology 2005, 337, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Barrett, J.W.; Werden, S.J.; Wang, F.; McKillop, W.M.; Jimeneza, J.; Villeneuve, D.; McFadden, G.; Dekaban, G.A. Myxoma virus M130R is a novel virulence factor required for lethal myxomatosis in rabbits. Virus Res. 2009, 144, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.X.; Teoh, M.L.T.; Moon, M.; McFadden, G.; Evans, D.H. Leporipoxvirus Cu-Zn superoxide dismutase homologs inhibit cellular superoxide dismutase, but are not essential for virus replication or virulence. Virology 2002, 296, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Teoh, M.L.T.; Turner, P.V.; Evans, D.H. Tumorigenic poxviruses up-regulate intracellular superoxide to inhibit apoptosis and promote cell proliferation. J. Virol. 2005, 79, 5799–5811. [Google Scholar] [CrossRef] [PubMed]
- Teoh, M.L.T.; Walasek, P.J.; Evans, D.H. Leporipoxvirus. Cu, Zn-superoxide dismutase (SOD) homologs are catalytically inert decoy proteins that bind copper chaperone for SOD. J. Biol. Chem. 2003, 278, 33175–33184. [Google Scholar] [CrossRef] [PubMed]
- Barrett, J.W.; Sypula, J.; Wang, F.; Alston, L.R.; Shao, Z.; Gao, X.; Irvine, T.S.; McFadden, G. M135R is a novel cell surface virulence factor of myxoma virus. J. Virol. 2007, 81, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Moss, B. Poxviridae: The viruses and their replication. In Fields’ Virology, 5th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; Volume 2, pp. 2905–2945. [Google Scholar]
- Jackson, R.J.; Hall, D.F.; Kerr, P.J. Myxoma virus encodes an α2,3-sialyltransferase that enhances virulence. J. Virol. 1999, 73, 2376–2384. [Google Scholar] [PubMed]
- Cameron, C.M.; Barrett, J.W.; Liu, L.; Lucas, A.R.; McFadden, G. Myxoma virus M141R expresses a viral CD200 (vOX-2) that is responsible for down-regulation of macrophage and T-cell activation in vivo. J. Virol. 2005, 79, 6052–6067. [Google Scholar] [CrossRef] [PubMed]
- Hovey Nerenberg, B.T.; Taylor, J.; Bartee, E.; Gouveia, K.; Barry, M.; Früh, K. The poxviral RING protein p28 is a ubiquitin ligase that targets ubiquitin to viral replication factories. J. Virol. 2005, 79, 597–601. [Google Scholar] [CrossRef] [PubMed]
- Brick, D.J.; Burke, R.D.; Schiff, L.; Upton, C. Shope fibroma virus RING finger protein N1R binds DNA and inhibits apoptosis. Virology 1998, 249, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Blanié, S.; Mortier, J.; Delverdier, M.; Bertagnoli, S.; Camus-Bouclainville, C. M148R and M149R are two virulence factors for myxoma virus pathogenesis in the European rabbit. Vet. Res. 2009, 40, e11. [Google Scholar] [CrossRef]
- Camus-Bouclainville, C.; Fiette, L.; Bouchiha, S.; Pignolet, B.; Counor, D.; Filipe, C.; Gelfi, J.; Messud-Petit, F. A virulence factor of myxoma virus colocalizes with NF-kappaB in the nucleus and interferes with inflammation. J. Virol. 2004, 78, 2510–2516. [Google Scholar] [CrossRef] [PubMed]
- Blanié, S.; Gelfi, J.; Bertagnoli, S.; Camus-Bouclainville, C. MNF, an ankyrin repeat protein of myxoma virus, is part of a native cellular SCF complex during viral infection. Virol. J. 2010, 7, e56. [Google Scholar] [CrossRef]
- Messud-Petit, F.; Gelfi, J.; Delverdier, M.; Amardeilh, M.-F.; Py, R.; Sutter, G.; Bertagnoli, S. Serp 2, an inhibitor of the interleukin-1β-converting enzyme, is critical in the pathobiology of myxoma virus. J. Virol. 1998, 72, 7830–7839. [Google Scholar] [PubMed]
- Guérin, J.-L.; Gelfi, J.; Camus, C.; Delverdier, M.; Whisstock, J.C.; Amardeihl, M.-F.; Py, R.; Bertagnoli, S.; Messud-Petit, F. Characterization and functional analysis of Serp 3: A novel myxoma virus-encoded serpin involved in virulence. J. Gen. Virol. 2001, 82, 1407–1417. [Google Scholar] [PubMed]
- Collin, N.; Guérin, J.-L.; Drexler, I.; Blanié, S.; Gelfi, J.; Boullier, S.; Foucras, G.; Sutter, G.; Messud-Petit, F. The poxviral scrapin MV-LAP requires a myxoma viral infection context to efficiently downregulate MHC-I molecules. Virology 2005, 343, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Guérin, J.L.; Gelfi, J.; Boullier, S.; Delverdier, M.; Bellanger, F.A.; Bertagnoli, S.; Drexler, I.; Sutter, G.; Messud-Petit, F. Myxoma virus leukemia-associated protein is responsible for major histocompatibility complex class I and Fas-CD95 down-regulation and defines scrapins, a new group of surface cellular receptor abductor proteins. J. Virol. 2002, 76, 2912–2923. [Google Scholar] [CrossRef] [PubMed]
- Mansouri, M.; Bartee, E.; Gouveia, K.; Hovey Nerenberg, B.T.; Barrett, J.; Thomas, L.; Thomas, G.; McFadden, G.; Früh, K. The PHD/LAP-domain protein M153R of myxoma virus is a ubiquitin ligase that induces the rapid internalization and lysosomal destruction of CD4. J. Virol. 2003, 77, 1427–1440. [Google Scholar] [CrossRef] [PubMed]
- Gedey, R.; Jin, X.-L.; Hinthong, O.; Shisler, J.L. Poxviral regulation of the host-NF-κB response: The vaccinia virus M2L protein inhibits induction of NF-κB activation via an ERK2 pathway in virus-infected human embryonic kidney cells. J. Virol. 2006, 80, 8676–8685. [Google Scholar] [CrossRef] [PubMed]
- Ramelot, T.A.; Cort, J.R.; Yee, A.A.; Liu, F.; Goshe, M.B.; Edwards, A.M.; Smith, R.D.; Arrowsmith, C.H.; Dever, T.E.; Keenedy, M.A. Myxoma virus immunomodulatory protein M156R is a structural mimic of eukaryotic translation initiation factor eIF2α. J. Mol. Biol. 2002, 322, 943–954. [Google Scholar] [CrossRef] [PubMed]
- Upton, C.; Macen, J.L.; Wishart, D.S.; McFadden, G. Myxoma virus and malignant rabbit fibroma virus encode a serpin-like protein important for viral virulence. Virology 1990, 179, 618–631. [Google Scholar] [CrossRef] [PubMed]
- Fenner, F.; Woodroofe, G.M. The pathogenesis of infectious myxomatosis: The mechanism of infection and the immunological response in the European rabbit (Oryctolagus cuniculus). Br. J. Exp. Pathol. 1953, 34, 400–410. [Google Scholar] [PubMed]
- Best, S.M.; Kerr, P.J. Coevolution of host and virus: The pathogenesis of virulent and attenuated strains of myxoma virus in resistant and susceptible European rabbits. Virology 2000, 267, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Best, S.M.; Collins, S.V.; Kerr, P.J. Coevolution of host and virus: Cellular localization of myxoma virus infection of resistant and susceptible European rabbits. Virology 2000, 277, 76–91. [Google Scholar] [CrossRef] [PubMed]
- Hurst, E.W. Myxoma and the Shope fibroma. I. The histology of myxoma. Br. J. Exp. Pathol. 1937, 18, 1–15. [Google Scholar]
- Hobbs, J.R. Studies of the nature of infectious myxoma of rabbits. Am. J. Hyg. 1928, 8, 800–839. [Google Scholar]
- Duclos, P.; Tuaillon, P.; Joubert, L. Histopathologie de l’atteinte cutanéo-muqueuse et pulmonaire de la myxomatose. Bull. Acad. Vet. 1983, 56, 95–104. (in French). [Google Scholar]
- Marlier, D.; Mainil, J.; Sulon, J.; Beckers, J.F.; Linden, A.; Vindevogel, H. Study of the virulence of five strains of amyxomatous myxoma virus in crossbred New Zealand White/Californian conventional rabbits, with evidence of long-term testicular infection in recovered animals. J. Comp. Path. 2000, 122, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Spiesschaert, B.; McFadden, G.; Hermans, K.; Nauwynck, H.; van de Walle, G.R. The current status and future directions of myxoma virus, a master in immune evasion. Vet. Res. 2011, 42, 76. [Google Scholar] [CrossRef] [PubMed]
- Mims, C. Aspects of the pathogenesis of viral diseases. Bacteriol. Rev. 1964, 28, 30–71. [Google Scholar] [PubMed]
- Corbet, G.B. Taxonomy and origins. In The European Rabbit. The History and Biology of a Succesful Colonizer; Thompson, H.V., King, C.M., Eds.; Oxford University Press: Oxford, UK, 1994; pp. 1–7. [Google Scholar]
- Flux, J.E.C. World distribution. In The European Rabbit. The History and Biology of a Successful Colonizer; Thompson, H.V., King, C.M., Eds.; Oxford University Press: Oxford, UK, 1994; pp. 8–21. [Google Scholar]
- Carneiro, M.; Rubin, C.-J.; DiPalma, F.; Albert, F.W.; Alfoldi, J.; Barrio, A.M.; Pielberg, G.; Rafati, N.; Sayyab, S.; Turner-Maier, J.; et al. Rabbit genome analysis reveals a polygenic basis for phenotypic change during domestication. Science 2014, 345, 1074–1079. [Google Scholar] [CrossRef] [PubMed]
- Rolls, E.C. They All Ran Wild; Angus and Robertson: Melbourne, Australia, 1969. [Google Scholar]
- Cooke, B.D. Rabbits: Manageable environmental pests or participants in new Australian ecosystems? Wildl. Res. 2012, 39, 279–289. [Google Scholar] [CrossRef]
- Aragão, H.B. Myxoma of rabbits. Mem. Inst. Oswaldo Cruz 1927, 20, 237–247. [Google Scholar]
- Fenner, F.; Fantini, B. Biological Control of Vertebrate Pests. The History of Myxomatosis—An Experiment in Evolution; CAB International: New York, NY, USA, 1999. [Google Scholar]
- Calaby, J.H.; Gooding, C.D.; Tomlinson, A.R. Myxomatosis in Western Australia. CSIRO Wildl. Res. 1960, 5, 89–101. [Google Scholar] [CrossRef]
- Williams, K.; Parer, I.; Coman, B.; Burley, J.; Braysher, M. Managing Vertebrate Pests: Rabbits; Bureau of Rural Sciences: Canberra, Australia, 1995. [Google Scholar]
- Moses, A. O virus do mixoma dos coelhos. Mem. Instit. Oswaldo Cruz 1911, 3, 46–53. (in Portuguese). [Google Scholar] [CrossRef]
- Fenner, F. Changes in the mortality rate due to myxomatosis in the Australian wild rabbit. Nature 1953, 172, 228–230. [Google Scholar] [CrossRef] [PubMed]
- Myers, K.; Marshall, I.D.; Fenner, F. Studies in the epidemiology of infectious myxomatosis of rabbits. III. Observations on two succeeding epizootics in Australian wild rabbits on the Riverine plain of south-eastern Australia. J. Hyg. 1954, 52, 337–360. [Google Scholar] [CrossRef] [PubMed]
- Marshall, I.D.; Dyce, A.L.; Poole, W.E.; Fenner, F. Studies in the epidemiogy of infectious myxomatosis of rabbits. IV. Observations of disease behaviour in two localities near the northern limit of rabbit infestation in Australia, May 1952 to April 1953. J. Hyg. 1955, 53, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Fenner, F.; Day, M.F.; Woodroofe, G.M. Epidemiological consequences of the mechanical transmission of myxomatosis by mosquitoes. J. Hyg. 1956, 54, 284–303. [Google Scholar] [CrossRef] [PubMed]
- Fenner, F.; Woodroofe, G.M. Changes in the virulence and antigenic structure of strains of myxoma virus recovered from Australian wild rabbits between 1950 and 1964. Aust. J. Exp. Biol. Med. Sci. 1965, 43, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Sampford, M.R. The estimation of response time distribution. III. Truncation and survival. Biometrics 1954, 10, 531–561. [Google Scholar] [CrossRef]
- Fenner, F. Biological control as exemplified by smallpox eradication and myxomatosis. Proc. R. Soc. Lond. B 1983, 218, 259–285. [Google Scholar] [CrossRef] [PubMed]
- Saint, K.M.; French, N.; Kerr, P. Genetic variation in Australian isolates of myxoma virus: An evolutionary and epidemiological study. Arch. Virol. 2001, 146, 1105–1123. [Google Scholar] [CrossRef] [PubMed]
- Kerr, P.J.; Hone, J.; Perrin, L.; French, N.; Williams, C.K. Molecular and serological analysis of the epidemiology of myxoma virus in rabbits. Vet. Microbiol. 2010, 143, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Fenner, F.; Marshall, I.D.; Woodroofe, G.M. Studies in the epidemiology of infectious myxomatosis of rabbits. I. Recovery of Australian wild rabbits (Oryctolagus. cuniculus) from myxomatosis under field conditions. J. Hyg. 1953, 51, 225–244. [Google Scholar] [CrossRef] [PubMed]
- Marshall, I.D. The influence of ambient temperature on the course of myxomatosis in rabbits. J. Hyg. 1959, 57, 484–497. [Google Scholar] [CrossRef] [PubMed]
- Sobey, W.R.; Menzies, W.; Conolly, D.; Adams, K.M. Myxomatosis: The effect of raised ambient temperature on survival time. Aust. J. Sci. 1968, 30, 322–323. [Google Scholar]
- Marshall, I.D.; Fenner, F. Studies in the epidemiology of infectious myxomatosis of rabbits. V. Changes in the innate resistance of wild rabbits exposed to myxomatosis. J. Hyg. 1958, 56, 288–302. [Google Scholar] [CrossRef] [PubMed]
- Marshall, I.D.; Douglas, G.W. Studies in the epidemiology of infectious myxomatosis of rabbits. VIII. Further observations on changes in the innate resistance of Australian wild rabbits exposed to myxomatosis. J. Hyg. 1961, 59, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Ross, J. Myxomatosis: The natural evolution of the disease. In Animal Disease in Relation to Animal Conservation; Academic Press: London, UK, 1982; Volume 50, pp. 77–92. [Google Scholar]
- Kerr, P.J.; Perkins, H.D.; Inglis, B.; Stagg, R.; McLaughlin, E.; Collins, S.V.; van Leeuwen, B.H. Expression of rabbit IL-4 by recombinant myxoma viruses enhances virulence and overcomes genetic resistance to myxomatosis. Virology 2004, 324, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Kerr, P.J.; McFadden, G. Immune responses to myxoma virus. Viral Immunol. 2002, 15, 229–246. [Google Scholar] [CrossRef] [PubMed]
- Sobey, W.R.; Conolly, D. Myxomatosis: Non-genetic aspects of resistance to myxomatosis in rabbits Oryctolagus cuniculus. Aust. Wildl. Res. 1986, 13, 177–188. [Google Scholar] [CrossRef]
- Williams, C.K.; Moore, R.J. Inheritance of acquired immunity to myxomatosis. Aust. J. Zool. 1991, 39, 307–312. [Google Scholar] [CrossRef]
- Parer, I.; Sobey, W.R.; Conolly, D.; Morton, R. Sire transmission of acquired resistance to myxomatosis. Aust. J. Zool. 1995, 43, 459–465. [Google Scholar] [CrossRef]
- Edmonds, J.W.; Nolan, I.F.; Shephard, R.C.H.; Gocs, A. Myxomatosis: The virulence of field strains of myxoma virus in a population of wild rabbits (Oryctolagus cuniculus L.) with high resistance to myxomatosis. J. Hyg. 1975, 74, 417–418. [Google Scholar] [CrossRef] [PubMed]
- Parer, I.; Sobey, W.R.; Conolly, D.; Morton, R. Virulence of strains of myxoma virus and the resistance of wild rabbits (Oryctolagus cuniculus L.), from different locations in Australasia. Aust. J. Zool. 1994, 42, 347–362. [Google Scholar] [CrossRef]
- Anderson, R.M.; May, R.M. Coevolution of hosts and parasites. Parasitology 1982, 85, 411–426. [Google Scholar] [CrossRef] [PubMed]
- Dwyer, G.; Levin, S.; Buttel, L. A simulation model of the population dynamics and evolution of myxomatosis. Ecol. Monogr. 1990, 60, 423–447. [Google Scholar] [CrossRef]
- Fenner, F.; Poole, W.E.; Marshall, I.D.; Dyce, A.L. Studies in the epidemiology of infectious myxomatosis of rabbits. VI. The experimental introduction of the European strain of myxoma virus into Australian wild rabbit populations. J. Hyg. 1957, 55, 192–206. [Google Scholar] [CrossRef] [PubMed]
- Parer, I.; Conolly, D.; Sobey, W.R. Myxomatosis: The effects of annual introductions of an immunizing strain and a highly virulent strain of myxoma virus into rabbit populations at Urana, New South Wales, Australia. Aust. Wildl. Res. 1985, 12, 407–424. [Google Scholar] [CrossRef]
- Shephard, R.C.H.; Edmonds, J.W. Myxomatosis: Changes in the epidemiology of myxomatosis coincident with the establishment of the European rabbit flea Spilopsyllus cuniculi (Dale) in the mallee region of Victoria. J. Hyg. 1978, 81, 399–403. [Google Scholar] [CrossRef] [PubMed]
- Berman, D.; Kerr, P.J.; Stagg, R.; van Leeuwen, B.H.; Gonzalez, T. Should the 40-year-old practice of releasing virulent myxoma virus to control rabbits (Oryctolagus cuniculus) be continued? Wildl. Res. 2006, 33, 549–556. [Google Scholar] [CrossRef]
- Seymour, R.M. A study of the interaction of virulence, resistance and resource limitation in a model of myxomatosis mediated by the European rabbit flea Spilopsyllus cuniculi (Dale). Ecol. Model. 1992, 60, 281–308. [Google Scholar] [CrossRef]
- Mead-Briggs, A.R.; Vaughan, J.A. The differential transmissibility of myxoma virus strains of differing virulence grades by the rabbit flea Spilopsyllus. cuniculi (Dale). J. Hyg. 1975, 75, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Mykytowycz, R. The effect of season and mode of transmission on the severity of myxomatosis due to an attenuated strain of the virus. Aust. J. Exp. Biol. Med. Sci. 1956, 34, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Joubert, L.; Duclos, P.; Toaillen, P. La myxomatose des garennes dans le sud-est. La myxomatose amyxomateuse. Rev. Méd. Vét. 1982, 133, 739–753. (in French). [Google Scholar]
- Cattadori, I.M.; Albert, R.; Boag, B. Variation in host susceptibility and infectiousness generated by co-infection: The myxoma-Trichostrongylus retortaeformis case in wild rabbits. J. R. Soc. Interface 2007, 4, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Thompson, H.V. The rabbit in Britain. In The European Rabbit. The History and Biology of a Successful Colonizer; Thompson, H.V., King, C.M., Eds.; Oxford University Press: Oxford, UK, 1994; pp. 64–107. [Google Scholar]
- Lloyd, H.G. Post-myxomatosis rabbit populations in England and Wales. EPPO Public. Ser. A 1970, 197–215. [Google Scholar]
- Sobey, W.R.; Menzies, W. Myxomatosis: The introduction of the European rabbit flea Spilopsyllus cuniculi (Dale) into Australia. Aust. J. Sci. 1969, 31, 404–405. [Google Scholar]
- Cooke, B.D. Changes in the age-structure and size of populations of wild rabbits in South Australia, following the introduction of European rabbit fleas, Spilopsyllus cuniculi (Dale), as vectors of myxomatosis. Aust. Wildl. Res. 1983, 10, 105–120. [Google Scholar] [CrossRef]
- Cooke, B.D. Notes on the comparative reproductive biology and the laboratory breeding of the rabbit flea Xenopsylla cunicularis Smit (Siphonaptera: Pulicidae). Aust. J. Zool. 1990, 38, 527–534. [Google Scholar] [CrossRef]
- Joubert, L.; Leftheriotis, E.; Mouchet, J. La Myxomatose; L’expansion Scientifique Francaise: Paris, France, 1972. (in French) [Google Scholar]
- Bárcena, J.; Pagès-Manté, A.; March, R.; Morales, M.; Ramírez, M.A.; Sánchez-Vizcaíno, J.M.; Torres, J.M. Isolation of an attenuated myxoma virus field strain that can confer protection against myxomatosis on contacts of vaccinates. Arch. Virol. 2000, 146, 759–771. [Google Scholar]
- Fenner, F.; Chapple, P.J. Evolutionary changes in myxoma virus in Britain. An examination of 222 naturally occurring strains obtained from 80 counties during the period October-November 1962. J. Hyg. 1965, 63, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.; Sanders, M.F. Changes in the virulence of myxoma virus strains in Britain. Epidem. Inf. 1987, 98, 113–117. [Google Scholar] [CrossRef]
- Mead-Briggs, A.R. Some experiments concerning the interchange of rabbit fleas, Spilopsyllus cuniculi (Dale), between living rabbit hosts. J. Anim. Ecol. 1964, 33, 13–26. [Google Scholar] [CrossRef]
- Ross, J.; Sanders, M.F. Innate resistance to myxomatosis in wild rabbits in England. J. Hyg. 1977, 79, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.; Sanders, M.F. The development of genetic resistance to myxomatosis in wild rabbits in Britain. J. Hyg. 1984, 92, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Brun, A.; Godard, A.; Moreau, Y. La vaccination contre la myxomatose. Vaccins heterologues et homologues. Bull. Soc. Vet. Med. Comp. 1981, 83, 251–254. (in French). [Google Scholar]
- Marlier, D.; Cassart, D.; Boucrat-Baralon, C.; Coignoul, F.; Vindevogel, H. Experimental infection of specific pathogen-free New Zealand white rabbits with five strains of amyxomatous myxoma virus. J. Comp. Path. 1999, 121, 369–384. [Google Scholar] [CrossRef] [PubMed]
- Marlier, D.; Mainil, J.; Linden, A.; Vindevogel, H. Infectious agents associated with rabbit pneumonia: Isolation of amyxomatous myxoma virus strains. Vet. J. 2000, 159, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Marlier, D.; Herbots, J.; Detilleux, J.; Lemaire, M.; Thiry, E.; Vindevogel, H. Cross-sectional study of the association between pathological conditions and myxoma virus seroprevalence in intensive rabbit farms in Europe. Prev. Vet. Med. 2001, 48, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.T.; Dunsmore, J.D.; Parer, I. Evidence for the existence of a latent myxoma virus in rabbits Oryctolagus cuniculus (L.). Nature 1972, 238, 99–101. [Google Scholar] [CrossRef] [PubMed]
- Kritas, S.K.; Dovas, C.; Fortomaris, P.; Petridou, E.; Farsang, A.; Koptopoulos, G. A pathogenic myxoma virus in vaccinated and non-vaccinated commercial rabbits. Res. Vet. Sci. 2008, 85, 622–624. [Google Scholar] [CrossRef] [PubMed]
- Marlier, D.; Mainil, J.; Boucrat-Baralon, C.; Linden, A.; Vindevogel, H. The efficacy of two vaccination schemes against experimental infection with a virulent amyxomatous or a virulent nodular myxoma virus strain. J. Comp. Path. 2000, 122, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Kerr, P.J.; Merchant, J.C.; Silvers, L.; Hood, G.; Robinson, A.J. Monitoring the spread of myxoma virus in rabbit populations in the southern tablelands of New South Wales, Australia. II. Selection of a virus strain that was transmissible and could be monitored by polymerase chain reaction. Epidemiol. Infect. 2003, 130, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, C.; Ramírez, E.; Castro, F.; Ferreras, P.; Alves, P.C.; Redpath, S.; Villafuerte, R. Field experimental vaccination campaigns against myxomatosis and their effectiveness in the wild. Vaccine 2009, 27, 6998–7002. [Google Scholar] [CrossRef] [PubMed]
- Guitton, J.-S.; Devillard, S.; Guénézan, M.; Fouchet, D.; Pontier, D.; Marchandeau, S. Vaccination of free-living juvenile wild rabbits (Oryctolagus. cuniculus) against myxomatosis improved their survival. Vet. Prev. Med. 2008, 84, 1–10. [Google Scholar] [CrossRef]
- Saito, J.K.; McKercher, D.G.; Castrucci, G. Attenuation of the myxoma virus and use of the living attenuated virus as an immunizing agent for myxomatosis. J. Infect. Dis. 1964, 114, 417–428. [Google Scholar] [CrossRef] [PubMed]
- Saurat, P.; Gilbert, Y.; Ganiere, J.-P. Etude d’une souche de virus myxomateux modifée. Rev. Méd. Vét. 1978, 129, 415–451. (in French). [Google Scholar]
- Pšikal, I.; Šmid, B.; Rodák, L.; Valíček, L.; Bendová, J. Atypical myxomatosis—Virus isolation, experimental infection of rabbits and restriction endonuclease analysis of the isolate. J. Vet. Med. 2003, B50, 259–264. [Google Scholar] [CrossRef]
- Camus-Bouclainville, C.; Gretillat, M.; Py, R.; Gelfi, J.; Guérin, J.-L.; Bertagnoli, S. Genome sequence of SG33 strain and recombination between wild-type and vaccine myxoma viruses. Emerg. Inf. Dis. 2011, 17, 633–638. [Google Scholar] [CrossRef]
- Kerr, P.J.; Rogers, M.B.; Fitch, A.; DePasse, J.V.; Cattadori, I.M.; Twaddle, A.C.; Hudson, P.J.; Tscharke, D.C.; Read, A.F.; Holmes, E.C.; et al. Genome scale evolution of myxoma virus reveals host pathogen adaptation and rapid geographic spread. J. Virol. 2013, 87, 12900–12915. [Google Scholar] [CrossRef] [PubMed]
- Strayer, D.S.; Sell, S. Immunohistology of malignant rabbit fibroma virus—A comparative study with rabbit myxoma virus. J. Nalt. Cancer Inst. 1983, 71, 106–116. [Google Scholar]
- Block, W.; Upton, C.; Mcfadden, G. Tumorigenic poxviruses: Genomic organization of malignant rabbit virus a recombinant between Shope fibroma virus and myxoma virus. Virology 1985, 140, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Upton, C.; Macen, J.L.; Maranchuk, R.A.; Delange, A.M.; McFadden, G. Tumorigenic poxviruses: Fine analysis of the recombination junctions in malignant rabbit fibroma virus a recombinant between Shope fibroma virus and myxoma virus. Virology 1988, 166, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Berkowitz, E.M.; Pogo, B.G. Molecular characterization of two strains of Shope fibroma virus. Virology 1985, 142, 437–440. [Google Scholar] [CrossRef] [PubMed]
- Kerr, P.J.; Ghedin, E.; DePasse, J.V.; Fitch, A.; Cattadori, I.M.; Hudson, P.J.; Tscharke, D.C.; Read, A.F.; Holmes, E.C. Evolutionary history and attenuation of myxoma virus on two continents. PLOS Pathog. 2012, 8, e1002950. [Google Scholar] [CrossRef] [PubMed]
- Firth, C.; Kitchen, A.; Shapiro, B.; Suchard, M.; Holmes, E.C.; Rambaut, A. Using time-stranded data to estimate evolutionary rates of double-stranded DNA viruses. Mol. Biol. Evol. 2010, 27, 2038–2051. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.W. The Glenfield strain of myxoma virus. Its use in Victoria. J. Agric. Agric. Vict. 1962, 60, 511–516. [Google Scholar]
- Mykytowycz, R. An attenuated strain of the myxomatosis virus recovered from the field. Nature 1953, 172, 448–449. [Google Scholar] [CrossRef] [PubMed]
- Kerr, P.J.; Jackson, R.J.; Robinson, A.J.; Swan, J.; Silvers, L.; French, N.; Clarke, H.; Hall, D.F.; Holland, M.K. Infertility in female rabbits (Oryctolagus. cuniculus) alloimmunized with the rabbit zona pellucida protein ZPB either as a purified recombinant protein or expressed by recombinant myxoma virus. Biol. Reprod. 1999, 61, 601–613. [Google Scholar] [CrossRef]
- Alzhanova, D.; Edwards, D.M.; Hammarlund, E.; Scholz, I.G.; Horst, D.; Wagner, M.J.; Upton, C.; Wiertz, E.J.; Slifka, M.K.; Früh, K. Cowpox virus inhibits the transporter associated with antigen processing to evade T cell recognition. Cell Host Microb. 2009, 6, 4333–4445. [Google Scholar] [CrossRef]
- Belsham, G.J.; Polacek, C.; Breum, S.Ø.; Larsen, L.E.; Bøtner, A. Detection of myxoma viruses encoding a defective M135R gene from clinical cases of myxomatosis; possible implications for the role of the M135R protein as a virulence factor. Virol. J. 2010, 7, e7. [Google Scholar] [CrossRef]
- Dalton, K.P.; Nicieza, I.; Baragaño, A.; Alonso, J.M.M.; Parra, F. Molecular characterisation of virulence graded field isolates of myxoma virus. Virol. J. 2010, 7, e49. [Google Scholar] [CrossRef]
- Muller, A.; Silva, E.; Abrantes, J.; Esteves, P.J.; Ferreira, P.G.; Carvalheira, J.C.; Nowotny, N.; Thompson, G. Partial sequencing of recent Portuguese myxoma virus field isolates exhibits a high degree of genetic stability. Vet. Microbiol. 2010, 140, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Davison, A.J.; Moss, B. Structure of vaccinia virus early promoters. J. Mol. Biol. 1989, 2210, 749–769. [Google Scholar] [CrossRef]
- Davison, A.J.; Moss, B. Structure of vaccinia virus late promoters. J. Mol. Biol. 1989, 210, 771–784. [Google Scholar] [CrossRef] [PubMed]
- Esposito, J.J.; Sammons, S.A.; Frace, A.M.; Osborne, J.D.; Olsen-Rasmussen, M.; Zhang, M.; Govil, D.; Damon, I.K.; Kline, R.; Laker, M.; et al. Genome sequence diversity and clues to the evolution of Variola (smallpox) virus. Science 2006, 313, 807–812. [Google Scholar] [CrossRef] [PubMed]
- Hatcher, E.L.; Hendrickson, R.C.; Lefkowitz, E.J. Identification of nucleotide-level changes impacting gene content and genome evolution in orthopoxviruses. J. Virol. 2014, 88, 13651–13668. [Google Scholar] [CrossRef] [PubMed]
- Cooke, B.D.; Fenner, F. Rabbit haemorrhagic disease and the biological control of wild rabbits, Oryctolagus cuniculus, in Australia and New Zealand. Wildl. Res. 2002, 29, 689–706. [Google Scholar] [CrossRef]
- Di Giallonardo, F.; Holmes, E.C. Viral biocontrol: Grand experiments in disease emergence and evolution. Trend Microbiol. 2015, 23, 83–90. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kerr, P.J.; Liu, J.; Cattadori, I.; Ghedin, E.; Read, A.F.; Holmes, E.C. Myxoma Virus and the Leporipoxviruses: An Evolutionary Paradigm. Viruses 2015, 7, 1020-1061. https://doi.org/10.3390/v7031020
Kerr PJ, Liu J, Cattadori I, Ghedin E, Read AF, Holmes EC. Myxoma Virus and the Leporipoxviruses: An Evolutionary Paradigm. Viruses. 2015; 7(3):1020-1061. https://doi.org/10.3390/v7031020
Chicago/Turabian StyleKerr, Peter J., June Liu, Isabella Cattadori, Elodie Ghedin, Andrew F. Read, and Edward C. Holmes. 2015. "Myxoma Virus and the Leporipoxviruses: An Evolutionary Paradigm" Viruses 7, no. 3: 1020-1061. https://doi.org/10.3390/v7031020
APA StyleKerr, P. J., Liu, J., Cattadori, I., Ghedin, E., Read, A. F., & Holmes, E. C. (2015). Myxoma Virus and the Leporipoxviruses: An Evolutionary Paradigm. Viruses, 7(3), 1020-1061. https://doi.org/10.3390/v7031020