
Two Year Field Study to Evaluate the Efficacy of Mamestra brassicae Nucleopolyhedrovirus Combined with Proteins Derived from Xestia c-nigrum Granulovirus

Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects, Viruses and Additives
2.2. Field Trials

{kind=link}
| First trial, 2008 | Second trial, 2008 | First trial, 2009 | Second trial, 2009 | |
|---|---|---|---|---|
| Plant | cabbage | cabbage | cabbage | broccoli |
| Date of transplanting | 15 4 2008 | 23 5 2008 | 20 4 2009 | 12 8 2009 |
| Bt treatment | Bt kurstaki 15 5 2008 | None | None | Bt kurstaki 3 9 2009 |
| Date of pest release | 27 5 2008 | 29 6–1 7 2008 | 29–30 5 2009 | 11–12 9 2009 |
| (quantity/plant) | (ca. 160 eggs) | (ca. 320 eggs) | (300–400 eggs) | (300–400 eggs or larvae) |
| Date of virus application | 2 6 2008 | 7 7 2008 | 4 6 2009 | 19 9 2009 |
| (Larval stage) | (late 1st-instar) | (2nd to 3rd-instars) | (2nd-instar) | (2nd-instar) |
| Concentration of NPV | 106 OBs/mL | 105 OBs/mL | 105 OBs/mL | 105 OBs/mL |
| Concentration of GVPs | 0 or 10 μg/mL | 0 or 10 μg/mL | 5 or 10 μg/mL | 5 or 10 μg/mL |
| Date of larval collection | 6 6 2008 | 11 7 2008 | 8 6 2009 | 23 9 2009 |
| (Larval stage) | (late 2nd) | (4th) | (late 2nd to mid-3rd) | (3rd to 4th) |
2.3. Rearing of Larvae and Examination of the Cause of Death
2.4. Data Analysis
3. Results
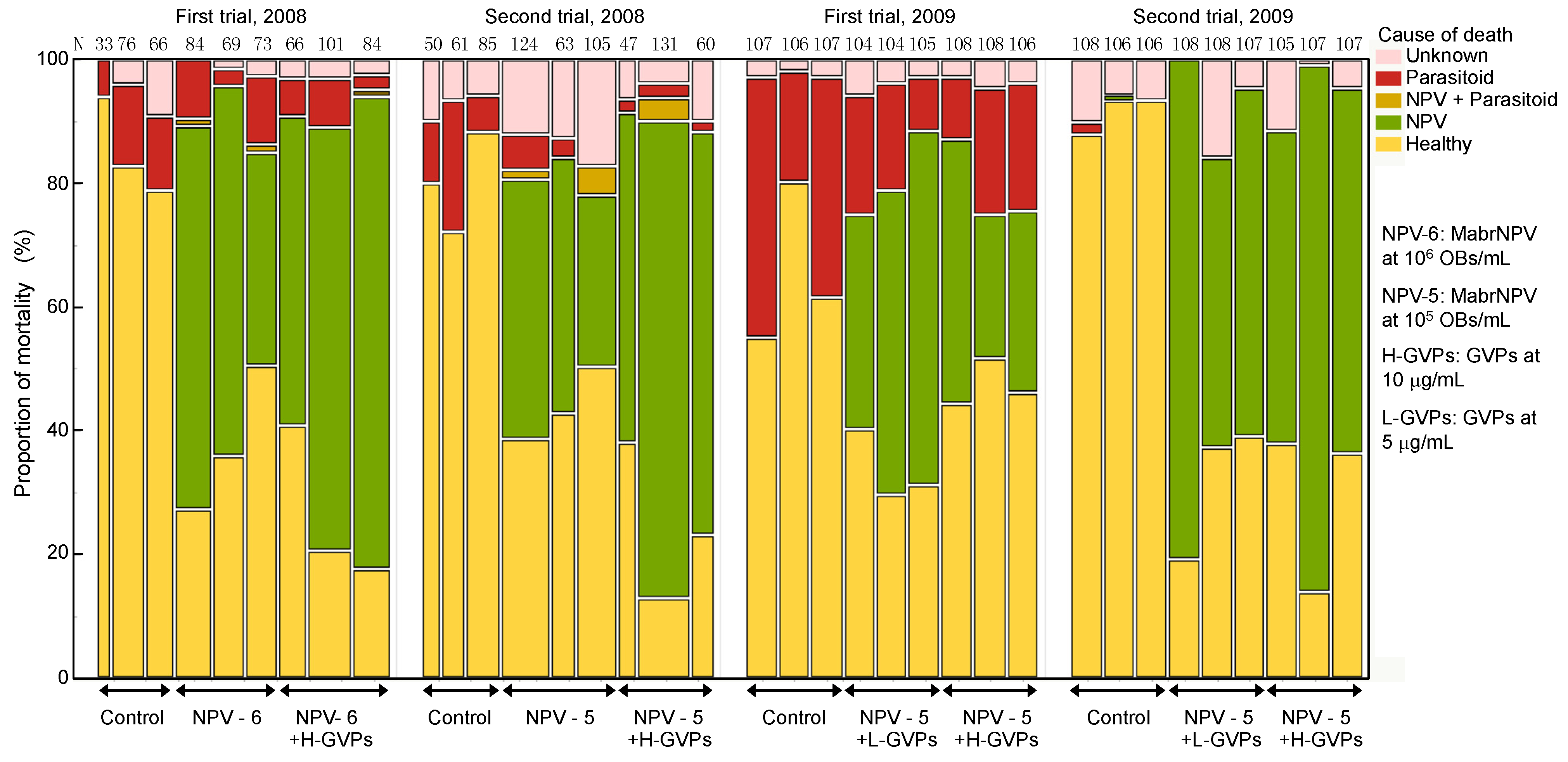
3.1. Field Trial in 2008
| Factor | Overall analysis 2 | Viral Infection 3 | Wasp Emergence 4 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| df | G | p | df | G | p | df | G | p | |
| First trial, 2008 (Cabbage) | |||||||||
| Treatment | 2 | 153.24 | <0.01 | 1 | 9.62 | <0.01 | 2 | 1.83 | 0.400 |
| Second trial, 2008 (Cabbage) | |||||||||
| Treatment | 2 | 146.95 | <0.01 | 1 | 50.94 | <0.01 | 2 | 6.57 | 0.038 |
| First trial, 2009 (Cabbage) | |||||||||
| Treatment | 2 | 75.63 | <0.01 | 1 | 13.79 | <0.01 | 2 | 4.67 | 0.097 |
| Plant 1 | 18 | 47.88 | <0.01 | 12 | 35.08 | <0.01 | 18 | 35.19 | <0.01 |
| Second trial, 2009 (Broccoli) | |||||||||
| Treatment | 2 | 340.21 | <0.01 | 1 | 1.76 | 0.185 | - 5 | ||
| Plant 1 | 18 | 102.39 | <0.01 | 12 | 74.18 | <0.01 | |||
| Overall Mortality | Mortality by Virus | Mortality by Wasps | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Treatment (crop) | Hazard Ratio 1 | 95% CL | Hazard Ratio 1 | 95% CL | Hazard Ratio 1 | 95% CL | |||
| Pairwise Treatment | Lower | Upper | Lower | Upper | Lower | Upper | |||
| First trial, 2008 (Cabbage)2 | |||||||||
| NPV alone/Cont. | 5.20 * | 3.54 | 7.90 | 1.51 | 0.81 | 2.81 | |||
| NPV + GVPs/Cont. | 7.35 * | 5.04 | 11.11 | 1.37 | 0.69 | 2.66 | |||
| NPV + GVPs/NPV alone | 1.41 * | 1.14 | 1.76 | 1.44 * | 1.14 | 1.83 | 0.91 | 0.46 | 1.76 |
| Second trial, 2008 (Cabbage)3 | |||||||||
| NPV alone/Cont. | 3.57 * | 2.53 | 5.18 | 0.51 * | 0.26 | 0.96 | |||
| NPV + GVPs/Cont. | 6.47 * | 4.59 | 9.36 | 0.43 * | 0.19 | 0.88 | |||
| NPV + GVPs/NPV alone | 1.81 * | 1.47 | 2.24 | 2.37 * | 1.87 | 3.02 | 0.84 | 0.37 | 1.84 |
| First trial, 2009 (Cabbage)4 | |||||||||
| Low GVPs/Cont. | 2.77 * | 2.19 | 3.54 | 0.83 | 0.58 | 1.18 | |||
| High GVPs/Cont. | 1.90 * | 1.49 | 2.45 | 0.69 * | 0.48 | 0.97 | |||
| High GVPs/Low GVPs | 0.69 * | 0.56 | 0.84 | 0.61 * | 0.46 | 0.79 | 0.82 | 0.55 | 1.23 |
| Second trial, 2009 (Broccoli)4 | |||||||||
| Low GVPs/Cont. | 17.37 * | 10.37 | 34.53 | ||||||
| High GVPs/Cont. | 19.79 * | 11.81 | 39.33 | ||||||
| High GVPs/ Low GVPs | 1.14 | 0.94 | 1.38 | 1.15 | 0.94 | 1.40 | |||
| Treatment (Crop) | No. of Observation | LT50 (Days) | 95% CL | Parametic Survival Analysis | |||
|---|---|---|---|---|---|---|---|
| Ingredients of Formulations | Lower | Upper | df | G | p | ||
| First trial, 2008 (Cabbage) | |||||||
| 106 OBs/mL of MabrNPV alone | 122 | 5.0 | 4.8 | 5.2 | 1 | 4.91 | 0.027 |
| 106 OBs/mL of MabrNPV + 10 μg/mL of GVPs | 167 | 4.7 | 4.6 | 4.9 | |||
| Second trial, 2008 (Cabbage) | |||||||
| 105 OBs/mL of MabrNPV alone | 113 | 4.2 | 3.9 | 4.5 | 1 | 5.87 | 0.015 |
| 105 OBs/mL of MabrNPV + 10 μg/mL of GVPs | 169 | 3.7 | 3.5 | 4.0 | |||
| First trial, 2009 (Cabbage) | |||||||
| 105 OBs/mL of MabrNPV + 5 μg/mL of GVPs | 147 | 4.2 | 4.0 | 4.5 | 1 | 0.55 | 0.459 |
| 105 OBs/mL of MabrNPV + 10 μg/mL of GVPs | 102 | 4.4 | 4.1 | 4.7 | |||
| Second trial, 2009 (Broccoli) | |||||||
| 105 OBs/mL of MabrNPV + 5 μg/mL of GVPs | 197 | 4.2 | 4.0 | 4.3 | 1 | 1.77 | 0.184 |
| 105 OBs/mL of MabrNPV + 10 μg/mL of GVPs | 207 | 4.0 | 3.9 | 4.2 | |||
| Treatment (Crop) | No. of Observation 1 | 50% Emergence (Days) 2 | 95% CL | Parametric Survival Analysis | |||
|---|---|---|---|---|---|---|---|
| Ingredients of Formulations | Lower | Upper | df | G | p | ||
| Second trial, 2008 (Cabbage) | |||||||
| Control | 23 | 3.1 a | 2.47 | 3.90 | |||
| 105 OBs/mL of MabrNPV alone | 16 | 2.1 a | 1.56 | 2.70 | 2 | 5.06 | 0.080 |
| 105 OBs/mL of MabrNPV + 10 μg/mL of GVPs | 10 | 2.4 a | 1.72 | 3.43 | |||
| First trial, 2009 (Cabbage) | |||||||
| Control | 102 | 6.2 a 3 | 5.9 | 6.4 | |||
| 105 OBs/mL of MabrNPV + 5 μg/mL of GVPs | 50 | 6.5 ab | 6.2 | 6.7 | 2 | 12.37 | 0.002 |
| 105 OBs/mL of MabrNPV + 10 μg/mL of GVPs | 56 | 6.9 b | 6.6 | 7.3 | |||
3.2. Field Trial in 2009
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kitazima, E. On the “Grasserie” of the Larva of the small cabbage-butterfly, Pieris rapae crucivora Boisd. Bull. Kagoshima Imp. Coll. Agric. For. 1938, 13, 111–116_2a. (In Japanese) [Google Scholar]
- Koyama, R. Studies on the epizootic diseases of forest insects in Japan (1) list of pathogen. Bull. Gov. For. Exp. Stn. 1959, 112, 23–31. [Google Scholar]
- Akutsu, K. The use of viruses for control of cabbage armyworm, Mamestra brassicae (LINNAEUS), and common cabbageworm, Pieris rapae crucivora Boisduval. In Proceedings of the Joint U.S.-Japan Seminar on Microbial Control of Insect Pests, Fukuoka, Japan, 21–23 April 1967; pp. 43–49.
- Okada, M. Studies on the utilization and mass production of Spodoptera litura Nuclear Polyhedrosis Virus for control of the tobacco cutworm, Spodoptera litura Fabricius. Bull. Chugoku Natl. Agric. Exp. Stn. Ser. E 1977, 12, 1–66. [Google Scholar]
- Kamiya, K.; Zhu, J.; Murata, M.; Laviña-Caoili, B.A.; Ikeda, M.; Kobayashi, M.; Kawamura, S. Cloning and comparative characterization of three distinct nucleopolyhedroviruses isolated from the common cutworm, Spodoptera litura (Lepidoptera: Noctuidae) in Japan. Biol. Control 2004, 31, 38–48. [Google Scholar] [CrossRef]
- Kamiya, K.; Suzuki, I.; Matsuo, N.; Kawamura, S. Development of a microbial pesticide for common cutworm, Spodoptera litura by the use of entomopathogenic viruses. Bull. Gifu Prefect. Agric. Technol. Cent. 2012, 12, 27–33. (In Japanese) [Google Scholar]
- Goto, C.; Tsutsui, H.; Honma, K.; Iizuka, T.; Nakashima, T. Stuides on nuclear polyhedrosis and granulosis viruses of the spotted cutworm, Xestia c-nigrum L. (Lepidoptera: Noctuidae). Jpn. J. Appl. Entomol. Zool. 1985, 29, 102–106. [Google Scholar] [CrossRef]
- Saito, O.; Goto, C. Control of the cabbage armyworm and the spotted cutworm by two baculovirus in sugar beet field. Annu. Rep. Soc. Plant Prot. N. Jpn. 1992, 43, 189–190. [Google Scholar]
- Sato, T.; Oho, N.; Kodomari, S. Utilization of granulosis viruses for controlling leafrollers in tea fields. Jpn. Agric. Res. Q. 1985, 19, 271–275. [Google Scholar]
- Asayama, T.; Osaki, N. A granulosis of the diamondback moth, Plutella xylostella. J. Invertebr. Pathol. 1970, 15, 284–286. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Hayakawa, T.; Ueno, Y.; Fujita, T.; Sano, Y.; Matsumoto, T. Sequence analysis of the Plutella xylostella granulovirus genome. Virology 2000, 275, 358–372. [Google Scholar] [CrossRef] [PubMed]
- Nakai, M. Biological control of tortricidae in tea fields in Japan using insect viruses and parasitoids. Virol. Sin. 2009, 24, 323–332. [Google Scholar] [CrossRef]
- Japan Plant Protection Association. Noyaku Yoran 2013 (Annual Statitistic on Pests and Pesticides); Japan Plant Protection Association: Tokyo, Japan, 2013; p. 749. (In Japanese) [Google Scholar]
- Bonning, B.C.; Boughton, A.J.; Jin, H.; Harrison, R.L. Genetic Enhancement of Baculovirus Insecticides. In Advances in Microbial Control of Insect Pests; Springer: New York, NY, USA, 2003; pp. 109–125. [Google Scholar]
- Allaway, G.P.; Payne, C.C. Host range and virulence of five baculoviruses from lepidopterous hosts. Ann. Appl. Biol. 1984, 105, 29–37. [Google Scholar] [CrossRef]
- Doyle, C.J.; Hirst, M.L.; Cory, J.S.; Entwistle, P.F. Risk assessment studies: Detailed host range testing of wild-type cabbage moth, Mamestra brassicae (Lepidoptera: Noctuidae), nuclear polyhedrosis virus. Appl. Environ. Microbiol. 1990, 56, 2704–2710. [Google Scholar] [PubMed]
- Choi, J.B.; Heo, W.I.; Shin, T.Y.; Bae, S.M.; Kim, W.J.; Kim, J.I.; Kwon, M.; Choi, J.Y.; Je, Y.H.; Jin, B.R.; et al. Complete genomic sequences and comparative analysis of Mamestra brassicae nucleopolyhedrovirus isolated in Korea. Virus Genes 2013, 47, 133–151. [Google Scholar] [CrossRef]
- Mukawa, S.; Goto, C. In vivo characterization of a group II nucleopolyhedrovirus isolated from Mamestra brassicae (Lepidoptera: Noctuidae) in Japan. J. Gen. Virol. 2006, 87, 1491–1500. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Tanada, Y. Biochemical properties of viral envelopes of insect baculoviruses and their role in infectivity. J. Invertebr. Pathol. 1978, 32, 202–211. [Google Scholar] [CrossRef]
- Shapiro, M.; Bell, R.A. Enhanced effectiveness of Lymantria dispar (Lepidoptera: Lymantriidae) nucleopolyhedrosis virus formulated with boric acid. Ann. Entomol. Soc. Am. 1982, 75, 346–349. [Google Scholar] [CrossRef]
- Cisneros, J.; Pérez, J.A.; Penagos, D.I.; Ruiz, V.J.; Goulson, D.; Caballero, P.; Cave, R.D.; Williams, T. Formulation of a nucleopolyhedrovirus with boric acid for control of Spodoptera frugiperda (Lepidoptera: Noctuidae) in Maize. Biol. Control 2002, 23, 87–95. [Google Scholar] [CrossRef]
- Shapiro, M.; Robertson, J.L. Enhancement of gypsy moth (Lepidoptera: Lymantriidae) baculovirus activity by optical brighteners. J. Econ. Entomol. 1992, 85, 1120–1124. [Google Scholar] [CrossRef]
- Martı́nez, A.M.; Goulson, D.; Chapman, J.W.; Caballero, P.; Cave, R.D.; Williams, T. Is it feasible to use optical brightener technology with a baculovirus bioinsecticide for resource-poor maize farmers in mesoamerica? Biol. Control 2000, 17, 174–181. [Google Scholar] [CrossRef]
- Lasa, R.; Ruiz-Portero, C.; Alcázar, M.D.; Belda, J.E.; Caballero, P.; Williams, T. Efficacy of optical brightener formulations of Spodoptera exigua multiple nucleopolyhedrovirus (SeMNPV) as a biological insecticide in greenhouses in southern Spain. Biol. Control 2007, 40, 89–96. [Google Scholar] [CrossRef]
- Tanada, Y. A synopsis of studies on the synergistic property of an insect baculovirus: A tribute to Edward A. Steinhaus. J. Invertebr. Pathol. 1985, 45, 125–138. [Google Scholar] [CrossRef]
- Derksen, A C.; Granados, R.R. Alteration of a lepidopteran peritrophic membrane by baculoviruses and enhancement of viral infectivity. Virology 1988, 167, 242–250. [Google Scholar]
- Goto, C. Enhancement of a nuclear polyhedrosis virus (NPV) infection by a granulosis virus (GV) isolated from the spotted cutworm, Xestia c-nigrum L. (Lepidoptera: Noctuidae). Appl. Entomol. Zool. 1990, 25, 135–137. [Google Scholar]
- Shapiro, M. Effect of two granulosis viruses on the activity of the gypsy moth (Lepidoptera: Lymantriidae) nuclear polyhedrosis virus. J. Econ. Entomol. 2000, 93, 1633–1637. [Google Scholar] [CrossRef] [PubMed]
- Arif, B.M. Recent advances in the molecular biology of entomopoxviruses. J. Gen. Virol. 1995, 76, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Mitsuhashi, W. Recent advances in studies for the application of a protein produced by entomopoxviruses (poxviridae) for insect-pest control. Jpn. Agric. Res. Q. 2009, 43, 289–294. [Google Scholar] [CrossRef]
- Tanada, Y.; Hukuhara, T. Enhanced infection of a nuclear-polyhedrosis virus in larvae of the armyworm, Pseudaletia unipuncta, by a factor in the capsule of a granulosis virus. J. Invertebr. Pathol. 1971, 17, 116–126. [Google Scholar] [CrossRef]
- Yamamoto, T.; Kita, H.; Tanada, Y. Capsule components of two strains of a granulosis virus of the armyworm (Pseudaletia unipuncta): Purification by affinity chromatography from proteinase-free capsules and analysis by peptide mapping. J. Gen. Virol. 1979, 45, 371–381. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Corsaro, B.G.; Granados, R.R. Location and nucleotide sequence of the gene encoding the viral enhancing factor of the Trichoplusia ni granulosis virus. J. Gen. Virol. 1991, 72, 2645–2651. [Google Scholar] [CrossRef] [PubMed]
- Gijzen, M.; Roelvink, P.; Granados, R. Characterization of viral enhancing activity from Trichoplusia ni granulosis virus. J. Invertebr. Pathol. 1995, 65, 289–294. [Google Scholar] [CrossRef]
- Roelvink, P.W.; Corsaro, B.G.; Granados, R.R. Characterization of the Helicoverpa armigera and Pseudaletia unipuncta granulovirus enhancin genes. J. Gen. Virol. 1995, 76, 2693–2705. [Google Scholar] [CrossRef] [PubMed]
- Goto, C.; Hayakawa, T.; Maeda, S. Genome organization of Xestia c-nigrum granulovirus. Virus Genes 1998, 16, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, T.; Ko, R.; Okano, K.; Seong, S.I.; Goto, C.; Maeda, S. Sequence analysis of the Xestia c-nigrum granulovirus genome. Virology 1999, 262, 277–297. [Google Scholar] [CrossRef] [PubMed]
- Mukawa, S.; Goto, C. Enhancement of nucleopolyhedrovirus infectivity against Mamestra brassicae (Lepidoptera: Noctuidae) by proteins derived from granulovirus and a fluorescent brightener. J. Econ. Entomol. 2007, 100, 1075–1083. [Google Scholar] [CrossRef] [PubMed]
- Mukawa, S.; Goto, C.; Suzuki, Y. Effect of proteins derived from granulovirus on the infectivity of Mamestra brassicae nucleopolyhedrovirus in Helicoverpa armigera (Lepidoptera: Noctuidae). Appl. Entomol. Zool. 2008, 43, 323–329. [Google Scholar] [CrossRef]
- Mukawa, S.; Goto, C. Mamestra brassicae nucleopolyhedrovirus infection and enhancing effect of proteins derived from Xestia c-nigrum granulovirus in Larvae of Mamestra brassicae and Helicoverpa armigera (Lepidoptera: Noctuidae) on Cabbage. J. Econ. Entomol. 2010, 103, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Mukawa, S.; Goto, C. Enhancing effect of proteins derived from Xestia c-nigrum granulovirus on Mamestra brassicae nucleopolyhedrovirus infection in larvae of Autographa nigrisigna (Lepidoptera: Noctuidae) on cabbage. Appl. Entomol. Zool. 2011, 46, 55–63. [Google Scholar] [CrossRef]
- Goto, C.; Minobe, Y.; Iizuka, T. Restriction endonuclease analysis and mapping of the genomes of granulosis viruses isolated from Xestia c-nigrum and five other noctuid species. J. Gen. Virol. 1992, 73, 1491–1497. [Google Scholar] [CrossRef] [PubMed]
- Hosmer, D.W.; Lemeshow, S.; May, S. Applied Survival Analysis: Regression Modeling of Time to Event Data, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2008; p. 416. [Google Scholar]
- Kleinbaum, D.G.; Klein, M. Survival Analysis a Self-Learning Text, 3rd ed.; Springer: Berlin, Germany, 2012; p. 700. [Google Scholar]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practices of Statistics in Biological Research, 3rd ed.; W.H. Freeman: New York, NY, USA, 1995; p. 880. [Google Scholar]
- Moscardi, F. Assessment of the application of baculoviruses for control of Lepidoptera. Annu. Rev. Entomol. 1999, 44, 257–289. [Google Scholar] [CrossRef] [PubMed]
- Moscardi, F.; De Souza, M.L.; De Castro, M.E.B. Baculovirus pesticides: Present state and future perspectives. In Microbes and Microbial Technology; Ahmad, I., Ahmad, F., Pichtel, J., Eds.; Springer: New York, NY, USA, 2011; pp. 415–445. [Google Scholar]
- Lacey, L.; Frutos, R.; Kaya, H.; Vail, P. Insect pathogens as biological control agents: Do they have a future? Biol. Control 2001, 21, 230–248. [Google Scholar] [CrossRef]
- Okuno, S.; Takatsuka, J.; Nakai, M.; Ototake, S.; Masui, A.; Kunimi, Y. Viral-enhancing activity of various stilbene-derived brighteners for a Spodoptera litura (Lepidoptera: Noctuidae) nucleopolyhedrovirus. Biol. Control 2003, 26, 146–152. [Google Scholar] [CrossRef]
- Goulson, D.; Marti, A.; Hughes, W.O.H.; Williams, T. Effects of optical brighteners used in biopesticide formulations on the behavior of pollinators. Biol. Control 2000, 19, 232–236. [Google Scholar] [CrossRef]
- Arakawa, T. Promotion of nucleopolyhedrovirus infection in larvae of the silkworm, Bombyx mori (Lepidoptera: Bombycidae) by flufenoxuron. Appl. Entomol. Zool. 2002, 37, 7–11. [Google Scholar] [CrossRef]
- Arakawa, T. Chitin synthesis inhibiting antifungal agents promote nucleopolyhedrovirus infection in silkworm, Bombyx mori (Lepidoptera: Bombycidae) larvae. J. Invertebr. Pathol. 2003, 83, 261–263. [Google Scholar] [CrossRef] [PubMed]
- Bixby-Brosi, A.J.; Potter, D.A. Can a chitin-synthesis-inhibiting turfgrass fungicide enhance black cutworm susceptibility to a baculovirus? Pest Manag. Sci. 2012, 68, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Boughton, A.J.; Lewis, L.C.; Bonning, B.C. Potential of Agrotis ipsilon nucleopolyhedrovirus for suppression of the black cutworm (Lepidoptera: Noctuidae) and effect of an optical brightener on virus efficacy. J. Econ. Entomol. 2001, 94, 1045–1052. [Google Scholar] [CrossRef] [PubMed]
- Gallo, L.G.; Corsaro, B.G.; Hughes, P.R.; Granados, R.R. In vivo enhancement of baculovirus infection by the viral enhancing factor of a granulosis virus of the cabbage looper, Trichoplusia ni (Lepidoptera: Noctuidae). J. Invertebr. Pathol. 1991, 58, 203–210. [Google Scholar] [CrossRef]
- Jiang, J.; Zeng, A.; Ji, X.; Wan, N.; Chen, X. Combined effect of nucleopolyhedrovirus and Microplitis pallidipes for the control of the beet armyworm, Spodoptera exigua. Pest Manag. Sci. 2011, 67, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Fang, J.; Zhong, W.; Liu, B. Interactions between Meteorus pulchricornis and Spodoptera exigua multiple nucleopolyhedrovirus. J. Insect Sci. 2013, 13, e12. [Google Scholar] [CrossRef]
- Chang, J.H.; Choi, J.Y.; Jin, B.R.; Roh, J.Y.; Olszewski, J.A.; Seo, S.J.; O’Reilly, D.R.; Je, Y.H. An improved baculovirus insecticide producing occlusion bodies that contain Bacillus thuringiensis insect toxin. J. Invertebr. Pathol. 2003, 84, 30–37. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goto, C.; Mukawa, S.; Mitsunaga, T. Two Year Field Study to Evaluate the Efficacy of Mamestra brassicae Nucleopolyhedrovirus Combined with Proteins Derived from Xestia c-nigrum Granulovirus. Viruses 2015, 7, 1062-1078. https://doi.org/10.3390/v7031062
Goto C, Mukawa S, Mitsunaga T. Two Year Field Study to Evaluate the Efficacy of Mamestra brassicae Nucleopolyhedrovirus Combined with Proteins Derived from Xestia c-nigrum Granulovirus. Viruses. 2015; 7(3):1062-1078. https://doi.org/10.3390/v7031062
Chicago/Turabian StyleGoto, Chie, Shigeyuki Mukawa, and Takayuki Mitsunaga. 2015. "Two Year Field Study to Evaluate the Efficacy of Mamestra brassicae Nucleopolyhedrovirus Combined with Proteins Derived from Xestia c-nigrum Granulovirus" Viruses 7, no. 3: 1062-1078. https://doi.org/10.3390/v7031062
APA StyleGoto, C., Mukawa, S., & Mitsunaga, T. (2015). Two Year Field Study to Evaluate the Efficacy of Mamestra brassicae Nucleopolyhedrovirus Combined with Proteins Derived from Xestia c-nigrum Granulovirus. Viruses, 7(3), 1062-1078. https://doi.org/10.3390/v7031062